Mir-106b對阿爾茨海默病晝夜節律調控的初步研究
2010-02-01 09:27:48王海林劉嘉琳宗園媛張連峰
中國比較醫學雜志 2010年4期
王海林,劉嘉琳,宗園媛,張連峰,秦 川
(中國醫學科學院醫學實驗動物研究所,北京協和醫學院比較醫學中心,北京 100021)
阿爾茨海默病(Alzheimer's disease,AD)是一種神經退行性疾病,以進行性癡呆為主要臨床表現,同時伴有精神行為異常和明顯的社會生活功能減退。超過80%的65歲以上的老年人存在晝夜節律的紊亂[1],如睡眠障礙,在AD病人中這種晝夜節律紊亂的比例更高[2],嚴重影響著 AD病人的生活質量。哺乳動物的生命節律由位于視交叉上核的生物鐘基因調控,NPAS2作為生物鐘基因的旁系同源類似物,在哺乳動物的前腦發揮著類似的功能,NPAS2缺陷的小鼠睡眠模式和行為的適應能力發生改變[3]。M icroRNAs(miRNAs)是近年來發現的長約18-25 nt小調節RNAs,通過與靶基因mRNA的3’UTR結合,在轉錄后水平調節基因的表達。現有的研究表明,microRNAs參與多種生理和病理過程,與AD的發生也密切相關[4,5],但是 m icroRNAs對靶基因的調節作用是否會影響AD病人的生活節律,目前這方面的研究尚未見報道。本實驗利用芯片和real time PCR的方法在AD模型小鼠中篩選出表達有差異的 microRNAs,并通過體外實驗探討microRNA是否在 AD病人的晝夜節律紊亂中發揮一定的調節作用。
1 材料和方法
1.1 m icroRNA芯片
μParafloTM microRNA微陣列基因表達實驗和分析由LC Sciences公司提供。miRNA探針序列信息來自于Sanger m iRbase Release 9.2版本數據庫,每種探針至少重復3次。數據處理和分析首先是扣除背景,計算重復點平均值和標準偏差,然后通過LOWESS(locally-weighted regression)過濾進行標準化。對于雙色標記實驗,將計算兩種檢測信號的比值(log2)和t-test的P值,以P <0.01定義顯著差異性表達。數據處理結果根據實驗設計采用hierarchical clustering、K-means/medians clustering或者ANOVA(analysis of variance)等方法。
1.2 轉基因小鼠的鑒定
將轉APPswe基因和 PSΔE9基因的雙轉 C57/ BL6小鼠和野生型 C57/BL6小鼠雜交,提取鼠尾DNA,PCR鑒定 APPswe/PSΔE9雙轉基因小鼠的基因表型,篩選陽性小鼠進行進一步實驗。
1.3 逆轉錄反應和real tim e PCR
利用m irVanaTMmiRNA Isolation Kit(Ambion)提取6月齡APPswe/PSΔE9雙轉基因小鼠和同月齡對照小鼠總RNA,在紫外分光光度計上測濃度。利用 m iScript Reverse Transcription Kit(Qiagen)將RNA逆轉錄為 cDNA。Real-time PCR是利用miScript SYBR Green PCR Kit(Qiagen)進行的,以U6為內參。用delta-delta C(t)的方法和t-test對數據進行分析和統計學處理,以P<0.05定義顯著差異性表達。
1.4 m ir-106b表達載體的構建
按照 BLOCK-iTTMPol II m iR RNAi Expression Vector Kit(invitrogen)的說明書進行,簡述如下:首先合成 mir-106b的 Top strand oligo(TGCTGTAAA GTGCTGACAGTGCAGATGTTTTGGCCACTGACTGAC ATCTGCACTCAGCACTTTA)和 bottom strand oligo (CCTGTAAAGTGCTGAGTGCAGATGTCAGTCAGTGG CCAAAACATCTGCACTGTCAGCACTTTAC),然后將二者退火結合生成ds oligo,在T4 DNA連接酶的作用下,將ds olig與pcDNATM6.2-GW/EmGFP-miR載體進行連接,把連接好的質粒轉化到感受態大腸桿菌中進行擴增,將菌液涂布在 LB培養板上(含50μg/m L壯觀霉素),37℃培養過夜,挑取單克隆,進行搖菌測序。陰性對照質粒由試劑盒提供。
1.5 m ir-106b穩轉細胞的構建與鑒定
SH-SY5Y細胞用DMEM培養基進行培養,其中添加10%熱滅活的胎牛血清和100 U/m L青鏈霉素,37℃、5%CO2孵箱中培養。轉染前1 d將細胞接種到24孔板,換成無抗生素的培養基,轉染按照Lipofectamine 2 000(Invitrogen)的說明書進行。轉染后6 h將24孔板中的培養基換成全培養基。轉染后第2天將24孔板的細胞用胰酶消化下來,按1:10傳到60 cm2培養皿中培養,在培養基中添加2 μg/m L的Blasticidin進行篩選。此后,每隔3 d換1次含2μg/m L的 Blasticidin的培養基,連續培養2周,然后在熒光顯微鏡下挑選帶GFP的陽性克隆,進行擴大培養。構建成功后,提取穩轉細胞的RNA,逆轉成 cDNA后,用 real-time PCR檢測 m ir-106b的表達,鑒定mir-106b穩轉細胞系的構建是否成功。
1.6 m ir-106b靶基因的預測
用Targetsan、Pictar、miRanda等 microRNA靶基因預測軟件,對mir-106b的靶基因進行預測。
1.7 W estern b lot
從穩轉細胞中提取總蛋白,蛋白上樣量為70 μg,利用10%的SDS-PAGE膠進行分離,然后將其轉移至NC膜上,室溫下用TBST配制的5%牛奶封閉1 h,與兔多抗NPAS2抗體(abcam)進行雜交,室溫下孵育2 h,用TBST洗膜3次,與HRP標記的羊抗兔 IgG和 β-actin進行雜交,室溫下孵育 1h,用TBST洗膜3次,然后進行化學發光。
1.8 雙熒光素酶報告檢測
根據 Targetsan、Pictar、m iRanda等靶基因預測軟件,預測mir-106b與NPAS2 3’UTR的結合位點。PCR擴增NPAS2的3’UTR(包含與m ir-106b的預測結合位點并帶Xho I和Not I兩個酶切位點),將PCR擴增產物克隆到 pMDTM18-T載體,命名為 TNPAS2-3’UTR。突變預測與 mir-106b結合位點的堿基,構建突變的 pMDTM18-T載體.根據 Targetsan的預測,NPAS2與 m ir-106b的結合位點有兩個,因此我們分別構建了兩個單突變 pMDTM18-T載體和一個雙突變pMDTM18-T載體分別命名為T-NPAS2-3’UTR-M1,T-NPAS2-3’UTR-M2,T-NPAS2-3’UTR-2M。psiCHECKTM-2載體(Promega)含有兩種熒光素酶:螢火蟲熒光素酶和海腎熒光素酶。將所構建的T-NPAS2-3’UTR、T-NPAS2-3’UTR-M1、T-NPAS2-3’ UTR-M2、T-NPAS2-3’ UTR-2M 載 體 與psiCHECKTM-2載體進行Xho I和Not I雙酶切實驗,將野生型或者含有突變位點的NPAS2的3’UTR克隆至psiCHECKTM-2載體的海腎熒光素酶開放讀碼框架的下游,構建含 NPAS2-3’UTR及突變的NPAS2-3’UTR的熒光素酶報告載體,分別命名為PNPAS2-3’UTR、P-NPAS2-3’UTR-M1、P-NPAS2-3’UTR-M2、P-NPAS2-3’UTR-2M。將所構建的熒光素酶報告載體與 mir-106b表達載體或者陰性對照載體在HEK-293T細胞進行共轉染,轉染后48h,用雙熒光素酶報告檢測試劑盒(Promega)及熒光素酶活性檢測儀分別檢測螢火蟲熒光素酶和海腎熒光素酶的活性,并將海腎熒光素酶的活性用螢火蟲熒光素酶的活性進行標準化,比較熒光強度的變化。
2 結果
2.1 芯片結果和real tim e PCR驗證的結果
芯片結果顯示:與野生型C57BL/6J小鼠相比,6月齡APPswe/PSΔE9小鼠腦組織中mir-106b表達水平明顯降低。由于芯片結果存在一定的假陽性,我們用real—time PCR的方法對芯片結果進行進一步驗證,選取6月齡APPswe/PSΔE9小鼠6只(雌雄各半)和同月齡的野生型C57BL/6J小鼠6只(雌雄各半),每只重復3次,經t檢驗結果有統計學差別(P=0.03),采用delta-delta C(t)的方法計算其比值為1.46,與野生型 C57BL/6J小鼠相比,6月齡APPswe/PSΔE9小鼠腦組織中m ir-106b的表達水平升高。6月齡小鼠腦組織中 mir-106b表達情況的real time PCR結果見圖1。
2.2 穩轉細胞的構建及鑒定結果。
我們構建的mir-106 b表達載體和試劑盒里提供的陰性對照質粒均含有綠色熒光蛋白(GFP)表達結構域,當其轉染至細胞后,能夠表達GFP,,因此我們可以通過熒光顯微鏡來觀察所構建的穩轉細胞的情況。穩轉細胞系構建成功后,可見80%以上的細胞均表達GFP(圖2,見彩插1)。
Real time PCR檢測穩轉細胞m ir-106b的表達,共檢測了6個m ir-106b穩轉細胞克隆(命名為m ir-106b-1、2、3、4、5、6)。用delta-delta C(t)的方法進行計算,我們發現與陰性對照質粒的穩轉細胞相比,m ir-106b穩轉細胞的m ir-106b的表達升高2~5倍,說明我們的穩轉細胞的構建是成功的,real-time PCR的結果見圖3。
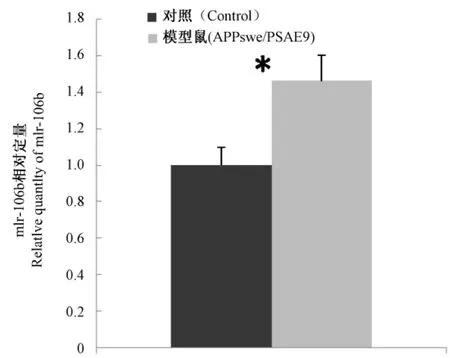
圖1 6月齡小鼠mir-106b表達情況的real time PCR結果Fig.1 The expression level of mir-106b in six-month old mice detected by real-time PCR注:“*”:P<0.05。 note:“*”:P<0.05.
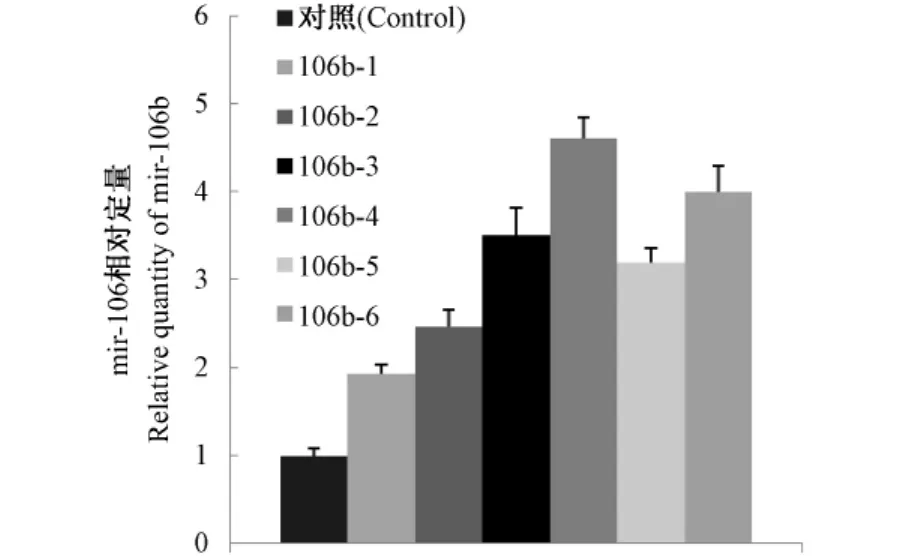
圖3 mir-106b穩轉細胞系real time PCR鑒定結果Fig.3 The real-time PCR Results of mir-106b stable transfection cell lines
2.3 W estern b lot結果
用提取陰性對照質粒穩轉細胞和 mir-106b穩轉細胞總蛋白,進行 western blot檢測。我們發現,與陰性對照質粒的穩轉細胞相比,在m ir-106b穩轉細胞中,NPAS2表達水平明顯降低(圖4)。
2.4 雙熒光素酶報告檢測實驗結果
熒光素酶活性檢測儀分別檢測螢火蟲熒光素酶和海腎熒光素酶的活性,并且用螢火蟲熒光素酶的活性對海腎熒光素酶的活性進行標準化,可見與陰性對照相比,P-NPAS2-3’UTR與 mir-106b共轉染,海腎熒光素酶的活性下降。熒光素酶報告載體的NPAS2-3’UTR突變后,與mir-106b共轉染其海腎熒光素酶的活性升高;并且熒光素酶報告載體的NPAS2-3’UTR雙突變與mir-106b共轉染,海腎熒光素酶/螢火蟲熒光素酶的活性較單突變更高(圖5)。
3 討論
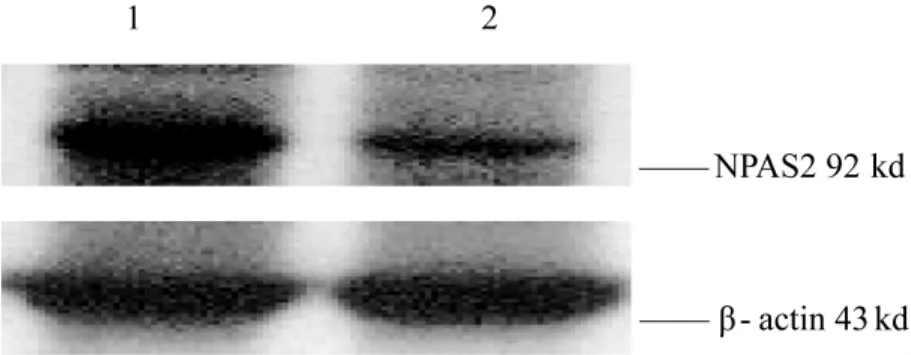
圖4 NPAS2的Western blot檢測結果Note:1:Stable transfection cell line generated with negative control plasm id;2:Stable transfection cell line generated with mir-106bFig.4 The results of Western blot analysis of NPAS2

圖5 雙熒光素酶報告檢測結果Fig.5 The results of dual-luciferase reporter assay
我們用real-time PCR對microRNA芯片的結果進行驗證,發現在AD雙轉小鼠模型腦組織中m ir-106b的表達升高,實驗組與對照組相比差異具有統計學意義。Hébert等[6]的研究發現 mir-106b的表達在AD病人的腦組織中有改變,可能通過調控淀粉樣前體蛋白(Amyloid precursor protein,APP)的表達參與AD的發病。這些研究表明mir-106b可能在AD的發生發展過程中發揮一定的調控作用。
NPAS2是一種主要在哺乳動物前腦表達的轉錄因子,與 BMAL1結合成異二聚體,激活內源性Per1、Per2和Cry1基因,從而在哺乳動物腦中發揮分子鐘的功能[7]。我們的研究發現 mir-106b可以調控NPAS2的表達:(1)在SH-SY5Y的mir-106b的穩轉細胞系中,m ir-106b的表達升高的同時,NPAS2的表達下降。(2)雙熒光素酶報告檢測實驗證實,mir-106b與 P-NPAS2-3’UTR共轉染與對照質粒與P-NPAS2-3’UTR共轉染相比,海腎熒光素酶/螢火蟲熒光素酶的活性明顯降低。microRNA靶基因預測軟件 Targetsans和 Pictar均預測 m ir-106b與NPAS2的3’UTR有2個結合位點,對這2個預測的結合位點進行突變,發現熒光素酶報告載體的NPAS2-3’UTR突變后與mir-106b共轉染,其海腎熒光素酶/螢火蟲熒光素酶的活性單突變較未突變的升高,雙突變的最高。證實mir-106b通過與NPAS2-3’UTR的這兩個位點結合來發揮其翻譯抑制作用,對這兩個位點進行突變能夠使 mir-106b喪失與NPAS2的 3’UTR的結合功能而無法發揮其對NPAS2的翻譯抑制作用。此外,我們的研究發現PNPAS2-3’UTR-2M 與 m ir-106b的共轉染較 PNPAS2-3’UTR與陰性對照質粒的共轉染海腎熒光素酶/螢火蟲熒光素酶的活性升高,理論上講兩者均不存在轉染 mir-106b的抑制作用。之所以出現這種現象,我們分析可能是因為 m icroRNA在許多組織和細胞廣泛表達,轉染所用的HEK-293T細胞存在的內源性的m icroRNA干擾的影響。
越來越多的研究發現microRNAs參與AD的發生發展[8],干擾 microRNAs的表達可能為 AD的治療提供一種新的思路和方法[9]。由于AD病人經常表現出較正常的老年人更為嚴重的晝夜節律紊亂,因此我們的研究表明 mir-106b可能通過調控生物鐘基因NPAS2的表達參與調控AD病人的生活節律,干擾mir-106b的表達可能有助于改善AD病人的這種紊亂,提高其生活質量。
[1] Foley DJ,Monjan AA,Brown SL,et al.Sleep complaints amongelderly persons:an epidemiologic study of three communities[J].Sleep,1995,18(6):425-432.
[2]Van Someren EJ.Circadian and sleep disturbances in the elderly[J].Exp Gerontol,2000,35(9-10):1229-1237.
[3]Dudley CA,Erbel-Sieler C,Estill SJ,et al.Altered patterns of sleep and behavioral adaptability in NPAS2-deficient mice[J].Science,2003,301(5631):379-383.
[4] Wang WX,Rajeev BW,Stromberg AJ,et al.The expression of microRNA miR-107 decreases early in Alzheimer's disease and may accelerate disease progression through regulation of beta-site amyloid precursor protein-cleaving enzyme 1[J].J Neurosci,2008,28(5):1213-1223.
[5]Hébert SS,HorréK,Nicola?L,et al.Loss ofmicroRNA cluster miR-29a/b-1 in sporadic Alzheimer's disease correlates with increased BACE1/beta-secretase expression[J].Proc Natl Acad Sci U S A,2008,105(17):6415-6420.
[6]Hébert SS,HorréK,Nicola?L,et al.MicroRNA regulation of Alzheimer's Amyloid precursor protein expression[J].Neurobiol Dis,2009,33(3):422-428.
[7] Reick M,Garcia JA,Dudley C,et al.NPAS2:an analog of clock operative in themammalian forebrain[J].Science,2001,293(5529):506-509.
[8] Boissonneault V,Plante I,Rivest S,et al.MicroRNA-298 and microRNA-328 regulate expression of mouse beta-amyloid precursor protein-converting enzyme 1[J].J Biol Chem,2009,284(4):1971-1981.
[9]Krützfeldt J,Rajewsky N,Braich R,et al.Silencing of microRNAs in vivo with‘antagomirs’[J].Nature,2005,438 (7068):685-689.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
