野生西瓜抗枯萎病鑒定研究
2010-02-10 06:56:46覃斯華黃金艷洪日新付崗韓金星李文信
長江蔬菜 2010年8期
覃斯華 ,黃金艷 ,洪日新 ,付崗 ,韓金星 ,李文信
(1.廣西農科院園藝研究所,廣西南寧,530007;2.廣西農科院植物保護研究所;3.廣西大學農學院)
西瓜枯萎病,俗稱死秧、死藤,是由鐮刀菌寄生引起的土傳病害,苗期至收獲期均可發生,是瓜類蔬菜的重要病害之一,全國各地均有發生,特別是重茬地區,發病情況尤為普遍和嚴重。目前栽培品種中高抗枯萎病的西瓜材料缺乏,枯萎病主要靠嫁接換根預防,野生西瓜就是高抗枯萎病的嫁接砧木之一。本試驗主要對從國內外收集到的野生西瓜材料進行抗枯萎病鑒定,并對西瓜幼苗受到枯萎病病菌侵染后,SOD、POD、CAT酶活性的變化情況進行分析測定。以期為深入研究西瓜枯萎病的發病規律、有效防治方法及抗性育種提供理論依據。
1 材料與方法
1.1 試驗材料
供試西瓜材料均由廣西農業科學院園藝研究所提供,野生材料分別為1號野生、2號野生、3號野生、4號野生、5號野生,以黑公子(栽培二倍體)、廣西三號(栽培三倍體)、廣西409(栽培四倍體)為對照。
1.2 菌種培養
西瓜枯萎病菌由鄭州果樹研究所、廣西農科院植物保護研究所提供。枯萎病菌種在廣西農科院植物保護研究所微生物研究室內進行培養。首先將活化的枯萎病菌種,接種到PDA培養基上,在28℃的恒溫箱中培養10 d,然后加無菌水,經4層紗布過濾后配成1×105個/mL的孢子懸浮液,備用。
1.3 試驗方法
西瓜在9月20日進行育苗,9月30日移栽至泡沫箱,試驗設3次重復,每重復10株,8個材料,共240株,并對每組材料設一對照,試驗小區隨機排列。
試驗采用浸根法,將株高和長勢一致的西瓜幼苗輕輕拔出,用清水洗凈根部后在孢子懸浮液中浸5 min后取出,重新種入滅菌砂土盤中。從接種之日起,每天觀察幼苗的發病情況,共調查25 d,統計總發病株數,計算發病率。 分別在處理后的第 0、2、4、6、8 天,取幼苗根系和生長點下第2片展開葉,測定各項指標,每處理分別取5株,對其枯萎病抗性生理進行研究。
1.4 抗性分級標準
抗性分級標準按照Elmstron和Hopkins的標準進行分級[3]。
1.5 SOD、POD、CAT活性的測定
①POD活性測定方法 POD活性采用愈創木酚法測定,按不同部位分別稱取根0.2 g、葉0.3 g,切碎放入研缽中,加1.5 mL pH值6.0的磷酸緩沖液,冰浴中研磨,4℃下以10 000 r/min離心15 min,取上清液待用。在光徑為1 cm的玻璃比色杯內,先加入pH值6.0磷酸緩沖液2 mL,0.05 mol/L愈創木酚1 mL,再加入10 μL酶液,最后加入l mL的2%H2O2迅速顛倒混勻,立即把比色杯插入比色架上。在470 nm波長下每隔30 s測定A470吸光值。每個樣品重復3次。酶活力計算:以時間為橫坐標,A470值(重復測定的平均值)為縱坐標,對所得數據進行作圖,求出圖中所作直線(回歸方程)的斜率,即△A470/min,該斜率代表酶反應的初速度,求出樣品中過氧化物酶活性,以△A470·min-1·g-1表示。

表1 抗性分級標準說明
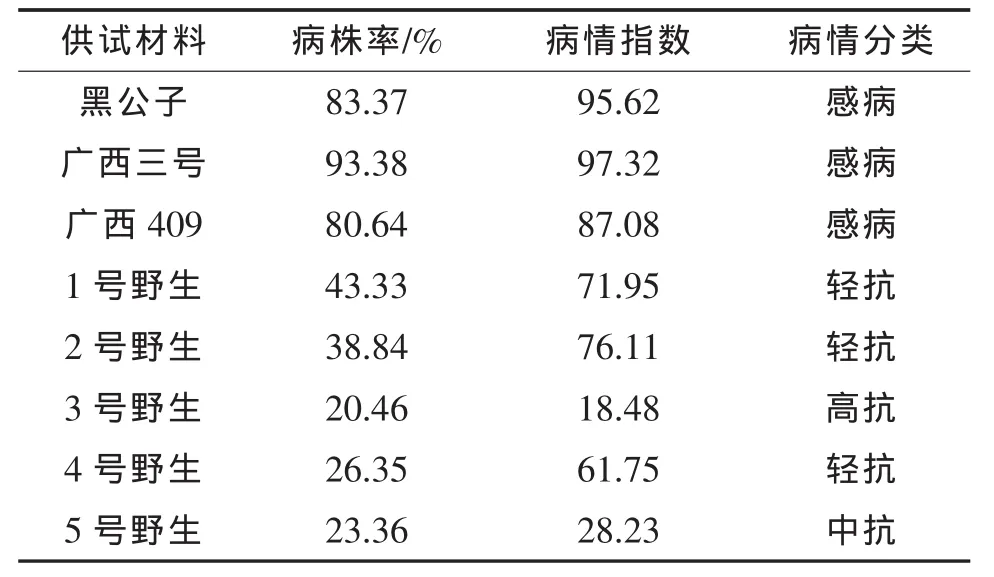
表2 植株病情分類結果
②SOD酶活性測定方法 SOD酶活性測定參照鄒琦主編的《植物生理學試驗指導》的方法,即按試驗要求分別稱取根0.2 g、葉0.3 g,加2 mL預冷提取介質冰浴中研磨成勻漿,加入介質沖洗研缽,并使終體積為10 mL。取5 mL于4℃10 000 r/min離心15 min,取上清液為酶提取液。一個酶活單位定義為將NBT還原抑制到對照一半(50%)時所需的酶量。
SOD 活性=(A0-As)×VT/[A0×0.5×FW×V1];其中:A0-照光對照管的光吸收值;As-樣品管的光吸收值;VT-樣液總體積 (mL);V1-測定時樣品用量(mL);FW-樣品鮮質量(g)。
③CAT酶活性測定方法 過氧化氫酶(CAT)活性按郝再彬方法測定,各處理分別取鮮葉和根系0.8 g,加入預冷的8 mL 50 mmol/L pH值7.8的磷酸緩沖液(內含1%聚乙烯聚吡咯烷酮PVPP)和少量石英砂,在冰浴上研磨成勻漿,勻漿在4℃12 000 r/min的冰凍離心機上離心15 min,上清液為粗酶提取液,然后取反應液3 mL(2 mL 100 mmol/L pH值7.8磷酸緩沖液,1 mL 0.08%雙氧水)加入粗酶液0.1 mL,迅速搖勻后倒入石英比色皿中,以100 mmol/L pH值7.8磷酸緩沖液為空白,于波長240 nm處測定吸光值,每隔15 s讀一次數,共3 min。以每1 min內A240減少0.1的酶量為一個酶活單位,用U·g-1·min-1FW表示CAT活性。
CAT 活性=△A240×酶提取液總量/(0.1×反應吸取的酶液量×樣品鮮質量×反應時間)。
2 結果與分析
2.1 植株病情分類
從表2可看出,野生材料的枯萎病抗性表現均強于栽培材料,其中3號野生高抗枯萎病,5號野生中抗枯萎病,1號野生、2號野生、4號野生輕抗枯萎病,對照(黑公子、廣西三號、廣西409)為感病材料。
2.2 抗病材料與感病材料苗體內POD酶活性的動態變化
由圖1可以看出,西瓜材料間POD活性的差異在正常生長狀態下,野生材料葉片中POD活性高于栽培材料,但野生材料間葉片中POD活性差異不明顯。根系中POD活性材料間差異明顯。
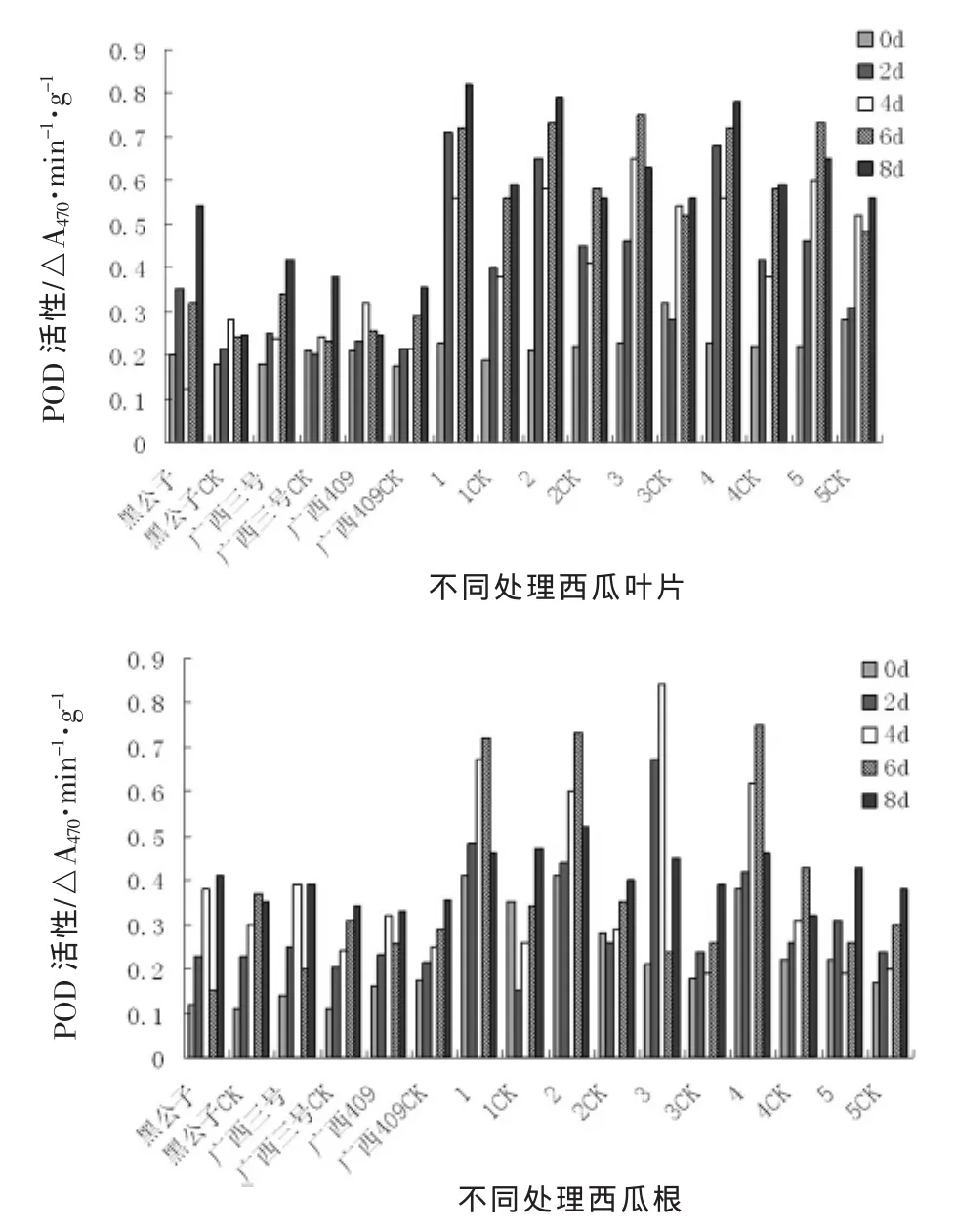
圖1 接種枯萎病后西瓜苗體內POD活性變化動態
枯萎病病菌侵染后,西瓜幼苗葉片和根系組織中POD活性明顯升高,但表現出不同的動態。抗病材料葉片組織中POD活性從接種后第2天就顯著高于對照,而感病材料則表現出不明顯變化,到接種后第4天之后,才明顯高于對照,但其峰值明顯低于抗性材料。根系組織中POD活性在接種后第2天就有明顯變化。而黑公子,廣西三號,廣西409三個材料處理與對照之間差異不顯著,POD活性變化不明顯,野生材料POD含量呈現先升后降的趨勢,但以高抗材料3號野生西瓜表現最為明顯。
抗病材料與感病材料苗體內POD酶活性的變化存在明顯差異,但5號野生表現不同于其他野生材料,這可能與其受到枯萎病病菌侵染后,本身自我調節能力強弱有關。
2.3 抗病材料與感病材料苗體內SOD酶活性的動態變化
由圖2可以看出,所有處理的西瓜材料苗體內SOD酶活性變化比較復雜,葉片內SOD酶活性的變化整體上呈現先升后降的趨勢,在苗體接種2 d后,SOD酶活性達到最高值,隨著時間的推移,SOD酶活性呈下降趨勢,但抗性材料的SOD酶活性下降緩慢,并且保持一定水平,而感病材料的SOD酶活性下降迅速,這表明感病材料在受到枯萎病病菌侵染后,活性細胞的氧化機制已被破壞,而抗性材料的自我修復調節能力高于感病材料,故表現比較好。
根系SOD酶活性的變化與葉片SOD酶活性變化相似,抗性材料的變化趨勢相似,且在受到枯萎病病菌侵染后,SOD酶活性迅速升高,隨著時間的推移,SOD酶活性下降明顯,所有材料的處理與對照差異均不顯著,這也說明在受到枯萎病病菌侵染后根系的氧化機制已被破壞,自我修復能力弱。
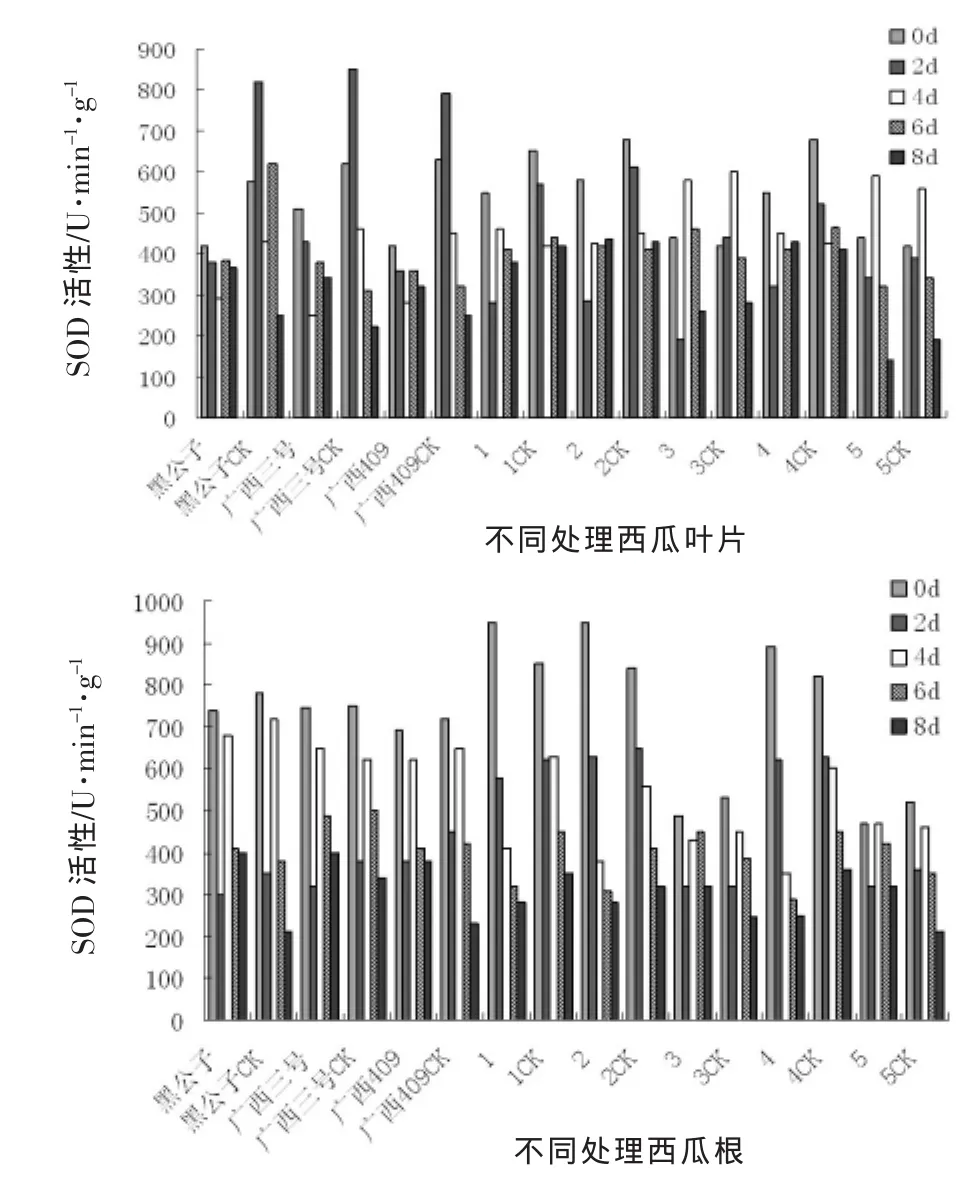
圖2 接種枯萎病后西瓜苗體內SOD活性變化動態
2.4 抗病和感病西瓜材料苗體內CAT酶活性的動態變化
由圖3可以看出,不論在西瓜材料的葉片還是在根系組織中,所有處理西瓜材料的苗體內CAT酶活性差異不顯著,感病材料與對照材料相比較,CAT酶活性變化略高,但均呈現出由高到低下降的變化趨勢。
在受到枯萎病病菌侵染后,西瓜材料苗體葉片內CAT酶活性立即升到最大,野生抗性材料高于感病材料。在接種后第2天,抗病材料葉片內CAT酶活性變化基本保持不變,并且隨著侵染時間的推移,下降趨勢緩慢,而感病材料則表現為迅速下降,但表現不穩定,在接種后第8天,西瓜感病材料苗體內CAT酶活性達到最小值,以廣西三號表現最突出。
在受到枯萎病病菌侵染后,西瓜材料苗體根系組織內CAT酶活性和葉片的相似,但是各個處理的CAT酶活性均小于葉片,且野生材料表現好于栽培材料,這也與前面進行的抗性鑒定試驗結果相吻合。
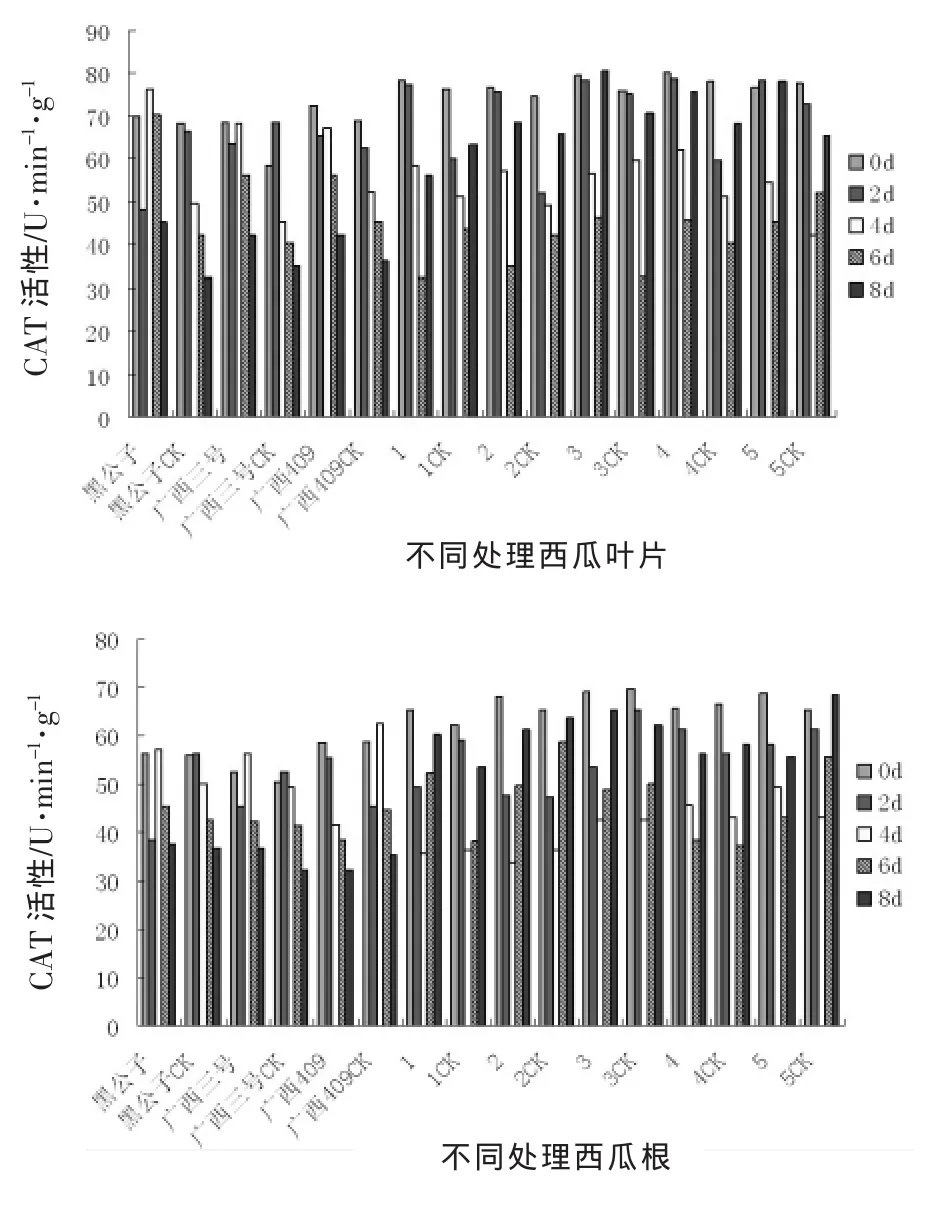
圖3接種枯萎病后西瓜苗體內CAT活性變化動態
3 小結與討論
西瓜在逆境脅迫的條件下,體內會產生大量的超氧陰離子自由基、活性氧等,造成細胞膜的傷害,超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)等屬于細胞膜保護酶,在清除活性氧中有重要功能。
POD參與木質素和植保素的合成,在植物抵抗病原物的侵入中起作用。有關過氧化物酶與植物抗病性的關系研究比較多,大多數研究結果表明,在植物與病原物互作中,不同類型的材料POD活性變化均表現出一定的差異,通常抗病材料的POD活性水平比感病材料POD活性水平高。本研究發現,在西瓜枯萎病的侵染病程中,野生材料葉片組織中POD活性從接種后第2天就顯著高于對照,而感病材料則表現不顯著,其峰值明顯低于野生抗病材料。根系組織中POD活性在接種后第2天就有明顯變化。這一試驗結果說明POD在西瓜對枯萎病的抵抗中具有重要作用。在受枯萎病菌侵染后,抗病材料立即高效表達與POD相關的基因,合成POD及相關的酶系,因此POD的活性高峰到來得早,且峰值高、持續時間長,在宏觀上表現為抗病。
超氧化物歧化酶(SOD)是需氧生物中普遍存在的一種含金屬酶。它與過氧化氫酶、過氧化物酶等酶協同作用防御活性氧或其他過氧化物自由基對細胞膜系統的傷害;超氧化物歧化酶可以催化氧自由基的歧化反應,生成過氧化氫,過氧化氫又可以被過氧化氫酶轉化成無害的分子氧和水。從結果中可以看出,在抗、感病材料中酶活性的變化及酶活性數量的增減,SOD酶和CAT酶并不相同,這說明植物體內活性氧的產生和消除是受SOD酶和CAT酶在內的多種酶的控制,而且不同酶間相互協調對植物的抗性反應起著十分重要的作用。
綜合各種酶的變化規律,可以看出,在接種2 d后,POD酶、SOD酶和CAT酶活性變化,中抗材料在受到枯萎病病菌侵染后,比感病材料表現穩定。輕抗材料介于二者之間,相對于中抗材料,它的酶活性變化幅度要大,相對于感病材料,它恢復正常的時間要短。這也可能是由于抗病材料能盡快地接受病菌侵染信號并迅速傳遞、及時啟動防衛反應的結果。
野生抗性材料在受到病原菌的侵染后,SOD酶和CAT酶活性的大小,均能在短時間內(接菌后2 d左右)基本恢復到原來的正常狀態,而感病材料則發生恢復慢,甚至難以恢復到原來的正常狀態,這說明材料是否抗病,POD、SOD和CAT酶活性的大小固然可以作為參考,但更為重要的是應看寄主在受到病原菌侵染后,POD、SOD和CAT酶活性能否通過自我調節而恢復到正常狀態能力的大小,有關其自我調節、維持動態平衡的抗性機制有待作進一步的探討。
[1]王金勝.農業生物化學技術[M].太原:山西科學技術出版社,1997.
[2]鄒琦.植物生理學實驗指導[M].北京:中國農業出版社,2000.
[3]Elmstron G W,Hopkins D L.Resistance of water melon cultivars toFusarium wilt[J].Plant disease,1981(65):825-827.
