人端粒酶逆轉錄酶啟動子調控的真核熒光表達載體的構建與表達
2010-03-19 00:13:46陰志剛王純耀張旭東
鄭州大學學報(醫學版) 2010年3期
董 翠,陰志剛,王純耀#,張旭東
1)鄭州大學生物工程系鄭州 450001 2)鄭州大學實驗動物中心鄭州 450052 3)鄭州大學第一附屬醫院腫瘤科鄭州 450052
#通訊作者,男,1962年11月生,教授,研究方向:分子生物學,E-mail:chunyao@zzu.edu.cn
端粒是真核細胞染色體末端的重復DNA序列,其生物學功能是防止染色體 DNA降解、末端融合、非正常重組和染色體的缺失[1]。端粒由端粒酶合成。人端粒酶主要由2部分組成,即端粒酶RNA模板和端粒酶逆轉錄酶(human telomerase reverse transcriptase,hTERT)[2]。端粒酶是腫瘤細胞特異的蛋白酶,90%以上的腫瘤細胞有端粒酶活性,而絕大多數正常細胞則檢測不到或活性極低[3]。hTERT的轉錄水平和腫瘤的端粒酶活性密切相關,因此hTERT的啟動子有可能成為一個靶向腫瘤的特異性啟動子[4]。根據這一特點,作者克隆了hTERT的啟動子,將其構建到真核熒光表達載體pEGFP-C3上,取代其原有的巨細胞病毒(CMV)啟動子,并檢測其功能,為后續的腫瘤靶向性治療奠定了基礎。
1 材料與方法
1.1 材料 基因組DNA小量抽提試劑盒、質粒DNA小量抽提試劑盒、膠回收試劑盒和PCR清潔試劑盒為愛思進生物技術(杭州)有限公司產品,T4連接酶和限制性內切酶為TaKaRa公司產品,LA Taq DNA聚合酶和pGEM-TEasy載體為Promega公司產品,DMEM細胞培養粉為Gibco公司產品,胎牛血清購于杭州四季青公司,脂質體LipofectamineTM2000購自美國Invitrogen公司。真核熒光表達載體pEGFP-C3為鄭州大學實驗動物中心與韓志強教授合作實驗室保種,其原有多克隆序列已通過 BglⅡ和HpaⅠ雙酶切去除。大腸桿菌E.coli Top10和DH5α及人胚腎 293A細胞由鄭州大學韓志強教授惠贈。
1.2 hTERTp引物的設計及合成 根據GenBank所提供的hTERT基因的序列設計引物,上游引物5'-ATTCGCGGGCACAGACGCCCAGGACCGCGC-3',下游引物5'-CGCGGGGGTGGCCGGGGCCAGGGCT TCCCA-3',擴增產物大小為276 bp。引物不含酶切位點,在表達載體pEGFP-C3上引入AseⅠ、SpeⅠ、NotⅠ、NdeⅠ、NruⅠ和AgeⅠ酶切位點。引物和多克隆位點由上海生工生物工程有限公司合成。
1.3 hTERTp的PCR擴增 使用基因組DNA小量抽提試劑盒,按照操作說明從 293A細胞中提取人基因組DNA,以人基因組DNA為模板,PCR擴增hTERTp。反應體系:2×GC Buffer 25μL,dNTPmix 6μL,人基因組DNA 3μL,上、下游引物各1μL,LA Taq酶0.5μL,ddH2O 13.5μL。反應條件:95℃預變性5min;95℃30 s,62℃30 s,72℃1 min,30個循環;72℃延伸6 min。10 g/L的瓊脂糖凝膠鑒定克隆產物為所需目的基因后,使用DNA膠回收試劑盒回收hTERTp。
1.4 目的基因與 T載體的連接、轉化及鑒定 膠回收的hTERTp用10 g/L的瓊脂糖凝膠定量,然后與pGEM-T Easy載體連接,連接體系轉化感受態E.coli Top10,在氨芐青霉素陽性LB平板上經藍白斑篩選。重組質粒經PCR和NotⅠ單酶切鑒定,并送上海生工生物工程有限公司測序。
1.5 重組載體pEGFP-C3的構建及鑒定 用AseⅠ和AgeⅠ雙酶切質粒pEGFP-C3,去除載體原來的CMV啟動子,10 g/L的瓊脂糖凝膠鑒定后回收目的片段。將合成的多克隆位點兩鏈退火,連接到 pEGFP-C3去除CMV啟動子的膠回收片段上,連接體系轉化感受態E.coliDH5α,并接種于卡那霉素陽性平板上。重組pEGFP-C3質粒分別經NotⅠ、NdeⅠ酶切鑒定。
1.6 pEGFP-C3-hTERTp載體的構建及鑒定NotⅠ酶切重組真核表達載體pEGFP-C3和pGEM-hTERTp,膠回收hTERTp和pEGFP-C3雙黏片段。將hTERTp與去磷酸化載體pEGFP-C3用T4連接酶連接,轉化感受態E.coli DH5α,并接種于卡那霉素陽性LB平板上。重組載體經PCR鑒定,并集菌送測序。
1.7 重組表達載體pEGFP-C3-hTERTp轉染293A細胞 轉染前 1 d,取對數期生長的 293A細胞接種到 6孔板,使轉染當天細胞匯合度達到90%~95%。用脂質體LipofectamineTM2000介導質粒轉染,設pEGFP-C3轉染組為陽性對照,按試劑說明操作。轉染 48~72 h后在熒光顯微鏡下觀察熒光表達情況并照相。
2 結果
2.1 hTERTp的PCR擴增結果 見圖1。擴增的276 bp條帶與預期相符。

圖1 hTERTp的PCR擴增結果M:DNA Marker;1:hTERTp。
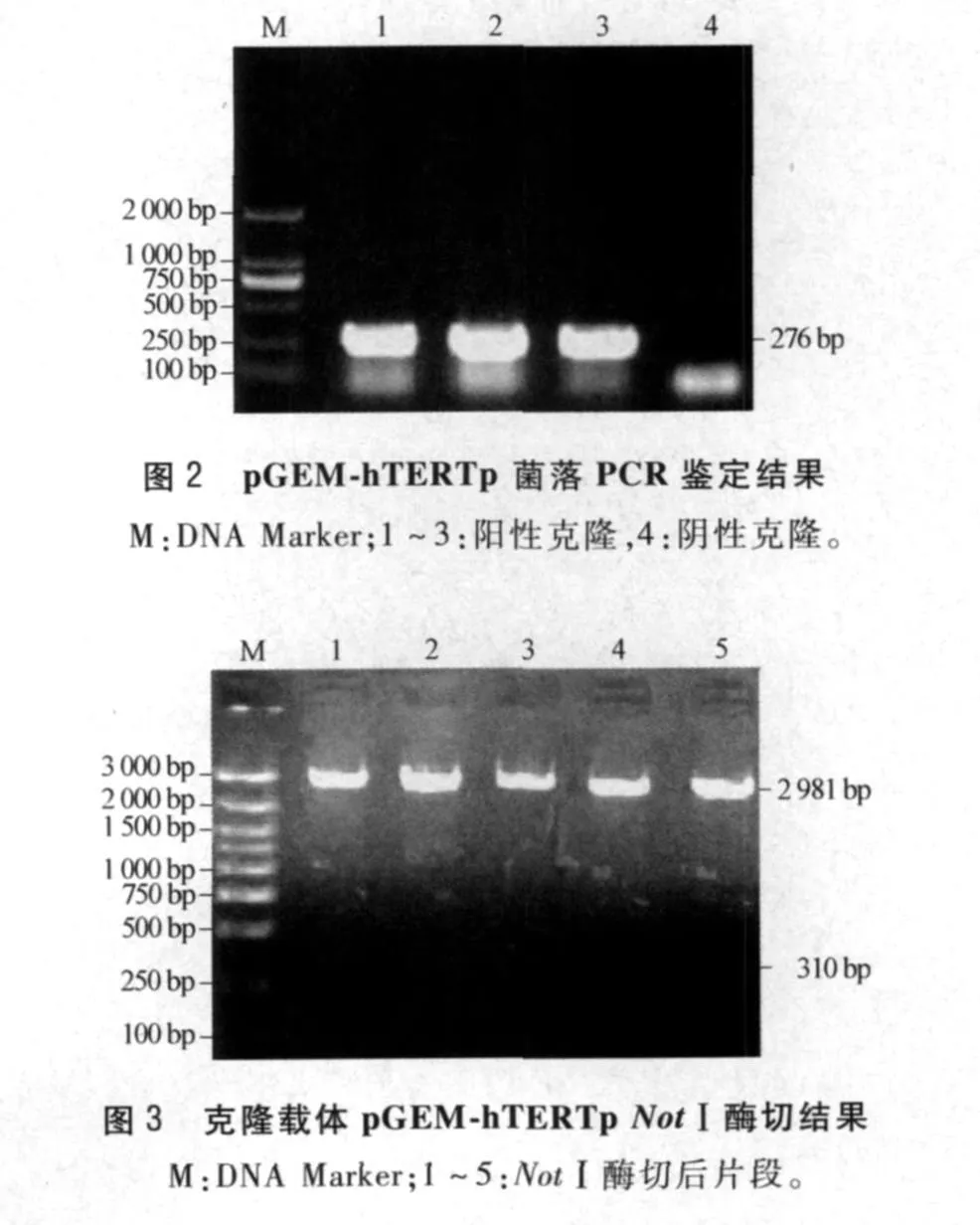
2.2 克隆載體pGEM-hTERTp的鑒定 pGEM-hTERTp菌落PCR鑒定結果見圖2。NotⅠ酶切克隆載體pGEM-hTERTp,得到310 bp的目的基因和2 981 bp的線狀片段(圖3)。
2.3 重組載體pEGFP-C3的鑒定 AseⅠ和AgeⅠ雙酶切重組質粒pEGFP-C3得到593 bp的CMV啟動子片段和3 955 bp的載體片段(圖4)。重組載體引入多克隆序列,經NotⅠ和NdeⅠ分別單酶切均得到3 986 bp的線性載體片段,與預期結果相符(圖5)。

2.4 重組表達載體pEGFP-C3-hTERTp的鑒定
重組表達載體pEGFP-C3-hTERTp菌落PCR鑒定結果見圖6。

圖6 重組表達載體
2.5 重組表達載體pEGFP-C3-hTERTp在真核細胞中的表達 見圖7。
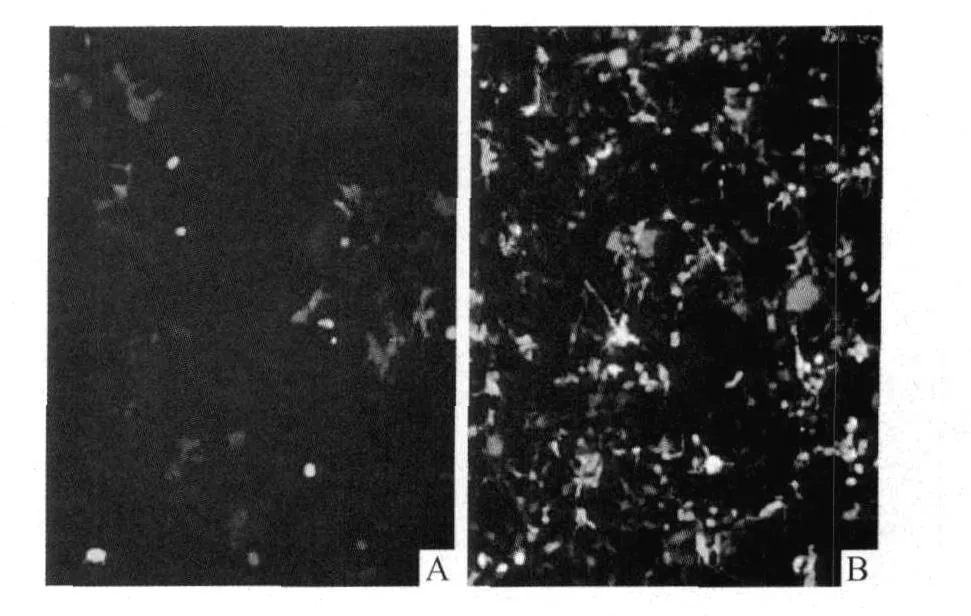
圖7 重組表達載體轉染293A細胞(×100)A:pEGFP-C3-hTERTp轉染組;B:陽性對照。
3 討論
端粒酶在大多數惡性腫瘤細胞中處于激活狀態,而在正常細胞中卻失活,并且其活性高低與腫瘤惡化程度有關,是目前所報道的最廣譜的腫瘤生物分子標記[5]。目前已發現的端粒酶組分中,只有hTERT在端粒酶陽性的腫瘤細胞中特異性表達,且表達水平與腫瘤細胞端粒酶的活性一致[6],提示hTERT啟動子可能是一個好的腫瘤特異性啟動子,以hTERT啟動子調控腫瘤治療基因,可以達到靶向腫瘤基因治療的目的。
作者采用PCR方法擴增了包含核心區的276 bp的hTERTp片段,并用其取代真核熒光表達載體pEGFP-C3原有CMV啟動子,構建重組載體pEGFPC3-hTERTp。重組質粒轉染端粒酶陽性的人胚腎293A細胞后,發現hTERTp能夠啟動綠色熒光蛋白GFP基因的表達。該研究為應用hTERTp作為轉錄調控元件,調控不同作用機制的治療基因在多種腫瘤細胞中的特異性表達,實現腫瘤靶向性基因治療奠定了基礎。
[1]許晶虹.端粒酶與癌基因[J].實用腫瘤雜志,2002,17(4):288
[2]胡建,覃文新,萬大方.端粒及端粒酶研究的最新進展[J].生命科學,2001,13(3):113
[3]李賀,張曉偉,張宗玉.端粒酶與腫瘤關系的研究進展[J].老年醫學與保健,2001,7(3):181
[4]黃雁西,趙學軍.端粒酶啟動子結構及其調控[J].癌癥,2001,20(6):667
[5]Nettelbeck DM,Jerome V,Muller R.Gene therapy:designer p romoters for tumour targeting[J].Trends Genet, 2000,16(4):174
[6]文忠.端粒酶催化亞單位在腫瘤特異基因治療作用中的研究[J].腫瘤防治研究,2005,32(2):122
