光皮樺SSR分子標記體系的建立
2010-05-29 13:16:04尤衛艷黃華宏程龍軍童再康朱玉球
浙江農林大學學報 2010年3期
尤衛艷,黃華宏,程龍軍,童再康,朱玉球
(浙江林學院 林業與生物技術學院,浙江 臨安 311300)
光皮樺Betula luminifera屬樺木科Betulaceae樺木屬Betula[1],為中國特有優良用材樹種,天然分布于秦嶺淮河流域以南的河南、四川、貴州、云南、安徽、湖北、湖南、廣東、廣西、江西、浙江、福建等10多個省(自治區)。其木材呈淡黃色或紅褐色,材質細膩堅韌,切面光滑,不撓不裂,干燥性能良好,廣泛應用于航空、建筑、家具、造紙等各行業,也可用于提取木醇和芳香油等物質。近年來光皮樺人工造林發展迅速,有關光皮樺的研究也不斷增多。在分子方面,謝一青等[2-3]通過對比和優化實驗,建立光皮樺DNA提取及其隨機擴增多態性DNA(random amplified polymorphic DNA,RAPD)技術分析體系。同時陳偉[4]應用RAPD和簡單序列重復區間擴增多態性(inter-simple sequence repeat,ISSR)技術對福建省光皮樺天然群體的的遺傳多樣性進行分析,發現11個群體明顯分為兩大類。童再康等[5]建立了光皮樺的擴增片段長度多態性(amplified fragment length polymorphism,AFLP)反應體系。簡單重復序列[6](simple sequence repeat,SSR)或者微衛星序列[7](microsatellite,MS),是一類由幾個到十幾個核苷酸串聯重復組成的DNA序列。其長度一般在100 bp以下,廣泛分布于生物體基因組的不同位置,由于重復次數不同及重復程度不完全造成的每個位點的多態性。微衛星序列兩端多是相對保守的單拷貝序列,因此可以根據兩端序列設計特異引物對基因組DNA進行PCR擴增,擴增片段的長度多態性可用作分子標記。SSR標記具有共顯性、高度重復性、高度豐富的多態性等優點,成為構建遺傳連鎖圖譜[8],研究群體遺傳學[9],進行分子標記輔助育種[10],繪制品種指紋圖譜[11],檢測品種純度[12]等的理想工具。SSR標記的眾多優點使它成為最理想的分子標記之一。本研究利用近緣種的引物在光皮樺中進行擴增,建立光皮樺的SSR反應體系,為應用SSR分子標記對光皮樺群體進行研究奠定基礎。
1 材料與方法
1.1 材料與試劑
光皮樺材料來自浙江林學院智能溫室,取正常生長的嫩葉,置-70℃冰柜保存備用。Taq DNA聚合酶(83.35 mkat·L-1),脫氧核糖核苷酸 dNTPs(各 2.5 mol·L-1),氯化鎂 MgCl2(25.0 mol·L-1)等試劑均購于由寶生物工程(大連)有限公司,引物由GenScript公司合成。
1.2 方法
1.2.1 光皮樺DNA的提取和檢測 本研究采用十六烷基三甲基溴化銨(CTAB)-硅珠吸附法提取光皮樺葉片DNA,用NanoDrop微量分光光度計(ND-1000)測定DNA的純度和濃度。同時每個樣品取5 μL,加1 μL上樣緩沖液混合后,于10.00 g·L-1瓊脂糖凝膠(含 0.05 g·L-1溴化乙錠)進行電泳檢測,在凝膠成像系統(Gel DocTM,Bio-Rad)下觀察并拍照。將樣品稀釋至20.0 mg·L-1,置于-20℃下保存。
1.2.2 PCR擴增及其產物檢測 聚合酶鏈式反應(PCR)擴增在ABI 9700 PCR儀上進行。擴增程序為:94℃,2 min;94℃,30 s;51~60℃(各引物退火溫度不同),30 s;72℃,30 s(30個循環);72℃,5 min。反應總體積為20 μL。體系建立過程中,分別對Mg2+濃度、dNTPs濃度、Taq DNA聚合酶濃度、引物濃度和模板DNA用量等設置不同的濃度梯度,以確定最佳反應體系。
PCR產物經15.00 g·L-1瓊脂糖凝膠電泳檢測,在凝膠成像系統(Gel DocTM,Bio-Rad)上觀察、拍照和分析。
1.2.3 PCR反應因素水平的確定為了確定PCR反應中5個因素(模板DNA用量、dNTPs濃度、Mg2+濃度、引物濃度、Taq DNA聚合酶濃度)的水平,選用引物AB084473采用單因素法進行試驗,實驗重復2次)。PCR反應的因素水平見表1。
1.2.4 引物合成 SSR引物的序列來源于近緣種日本白樺Betula platyphylla var.japonica[13]和歐洲白樺 B.pendula[14]的 36 對 SSR 引物。
1.2.5 電泳檢測 PCR擴增產物在80 g·L-1聚丙烯酰胺凝膠上電泳檢測,染色方法參照胥猛[15]等的方法,功率恒為75 W,電泳1.0~1.5 h。
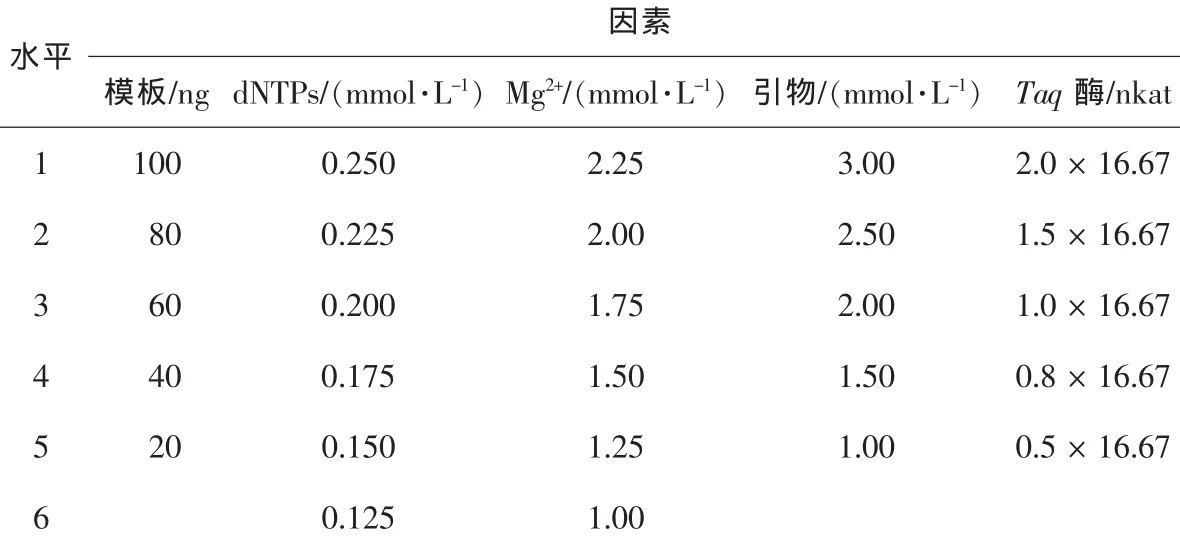
表1 聚合酶鏈式反應的因素水平Table1 Factors and levels of PCR
1.2.6 引物通用性研究 隨機選取引物AF310851,AB084479,AB084480(GenBank accession number),PCR擴增產物在80.00 g·L-1聚丙烯酰胺凝膠上電泳,參照李明芳[16]的方法,割取多態性片段,進行擴增。擴增后在瓊脂糖上電泳檢測,用試劑盒回收擴增產物,4℃連接過夜,轉入大腸桿菌Escherichia coli培養12 h以上,如藍白斑區別不明顯,4℃放置1 h,挑單克隆,培養8 h進行菌液PCR檢測,測序。
2 結果和分析
2.1 不同DNA用量對SSR反應結果的影響
當其他條件一定 (0.200 mmol·L-1dNTPs,1.50 mmol·L-1Mg2+,2.50 mmol·L-1引物,1 × 16.67 nkat Taq酶),由圖1可知,模板DNA用量為20或40 ng時,擴增出的條帶較弱;而用量為60或80 ng時,有較好的結果;當用量為100 ng時,擴增條帶反而變弱,可能是模板DNA濃度較大,影響反應體系濃度,進而影響PCR反應等因素造成的。為了節約試劑和減少非特異性條帶的產生,所以本實驗選用的模板DNA用量為60 ng。
2.2 不同dNTPs濃度對結果的影響
當其他條件一定(模板 DNA 用量為 60 ng,1.50 mmol·L-1Mg2+,2.50 mmol·L-1引物,1 × 16.67 nkat Taq酶),6個dNTPs濃度均可擴增出條帶。圖2顯示,當dNTPs濃度為0.125 mmol·L-1時,擴增出的條帶較弱;而dNTPs濃度為0.200,0.175,0.150 mmol·L-1時,有較好的結果;當濃度為0.250和0.225 mmol·L-1時,擴增條帶反而變弱,可能是擴增的片段較小,當dNTPs濃度增大時擴增產物不隨之增加造成的。體系選用0.175 mmol·L-1為dNTPs最終濃度。
2.3 不同Mg2+濃度對結果的影響
當其他條件一定(模板 DNA 用量為 60 ng,0.175 mmol·L-1dNTPs,2.50 mmol·L-1引物,1 × 16.67 nkat Taq酶),Mg2+濃度對PCR反應有很大影響,Mg2+濃度過高容易引起非特異性擴增,導致陽性克隆較多,影響數據分析結果。當Mg2+濃度為1.00和1.25 mmol·L-1時,擴增條帶很弱;而Mg2+濃度為1.50 mmol·L-1時,有較好的結果;當濃度為1.75 mmol·L-1時,擴增主條帶反而變弱;當Mg2+濃度為2.00和2.25 mmol·L-1時,主條帶亮度又增強,可能此時已經在進行非特異片段的擴增(圖3)。所以選用 Mg2+最終濃度為 1.50 mmol·L-1。
2.4 不同引物濃度對結果的影響
當其他條件一定(模板 DNA 用量為 60 ng,0.175 mmol·L-1dNTPs,1.50 mmol·L-1Mg2+,1 × 16.67 nkat Taq酶),引物濃度為1.00,1.50,2.00 mmol·L-1時,擴增的條帶極其微弱,幾乎檢測不到;當引物濃度增加到 2.50和 3.00 mmol·L-1時,條帶亮度增高(圖 4)。所以選用 2.50 mmol·L-1為最終濃度。
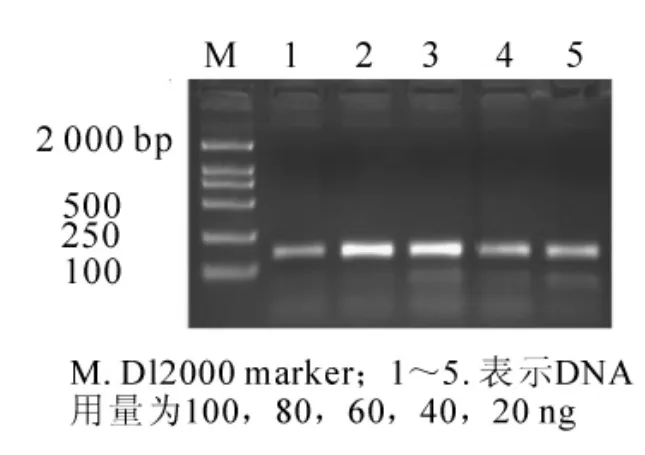
圖1 DNA用量對SSR反應結果的影響Figure1 Effects of DNA dosages on result of SSR reaction
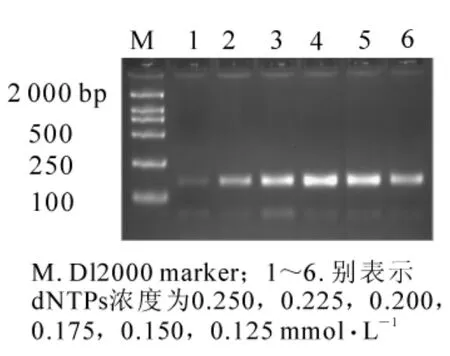
圖2 不同dNTPs濃度對SSR反應結果的影響Figure2 Effects of different dNTPs concentrations on result of SSR reaction
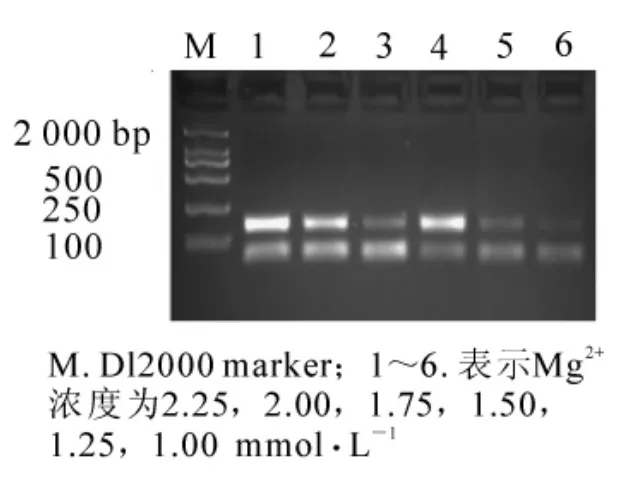
圖3 不同Mg2+濃度對SSR反應結果的影響Figure3 Effects of different Mg2+concentrations on result of SSR reaction
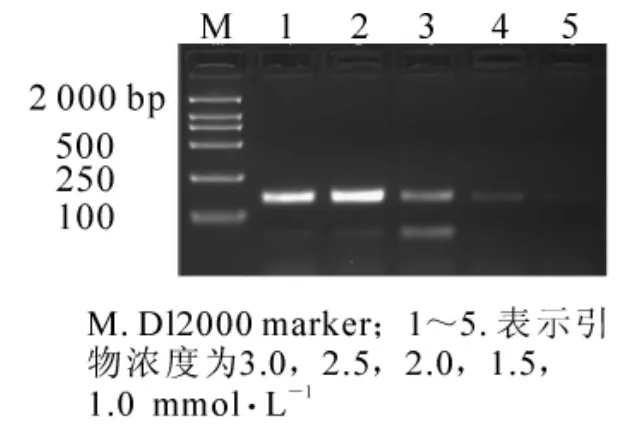
圖4 不同引物濃度對SSR反應結果的影響Figure4 Effects of different primers concentrations on result of SSR reaction
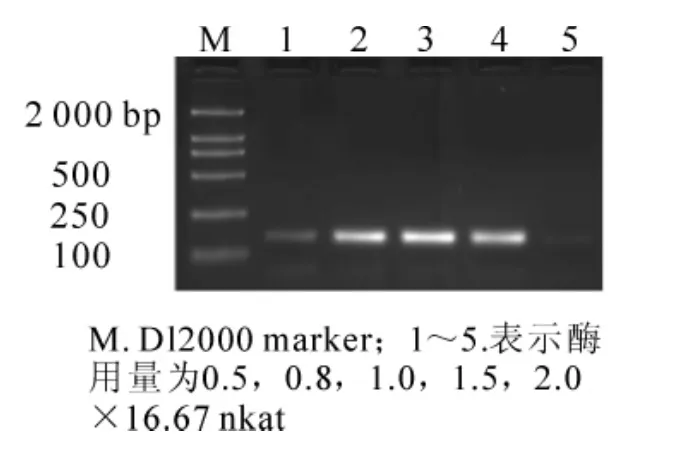
圖5 不同酶用量對SSR反應結果的影響Figure5 Effects of different enzyme dosages on result of SSR reaction
2.5 不同酶用量對結果的影響
當其他條件一定(模板 DNA 用量為 60 ng,0.175 mmol·L-1dNTPs,1.50 mmol·L-1Mg2+,2.50 mmol·L-1引物),Taq DNA聚合酶用量是PCR反應中最為重要的影響因子之一。Taq DNA聚合酶濃度高時,極易產生非特異性擴增。如圖5所示,5個處理均有擴增產物,但當Taq DNA聚合酶濃度為0.5×16.67 nkat時,擴增條帶很弱;濃度0.8×16.67,1.0×16.67,1.5×16.67 nkat時,譜帶亮且清晰;2.0×16.67時,條帶反而變弱。因此,確定Taq DNA聚合酶適宜用量為16.67 nkat。
2.6 退火溫度的影響
退火溫度的高低與PCR擴增特異性有顯著的相關性,而退火溫度主要取決于引物的解鏈溫度(melting temperature,Tm)值、引物與模板的配對程度等因素。當溫度較低時,非特異性條帶增加,退火溫度較高時,條帶變少(圖6)。所以體系選用的退火溫度比在日本白樺(其引物退火溫度為56℃)中提高了1℃。
2.7 歐洲白樺和日本白樺引物通用性檢驗

圖6 部分引物的在不同退火溫度下擴增產物在80 g·L-1聚丙烯酰胺凝膠上電泳檢測圖Figure6 Amplified product of some pairs of primer at different annealing temperature detecting in 80 g·L-1PAGE

表2 引物AF310851,AB084479,AB084480序列在GeneBank比對結果Table2 Blast results of AF310851,AB084479,AB084480 in GeneBank
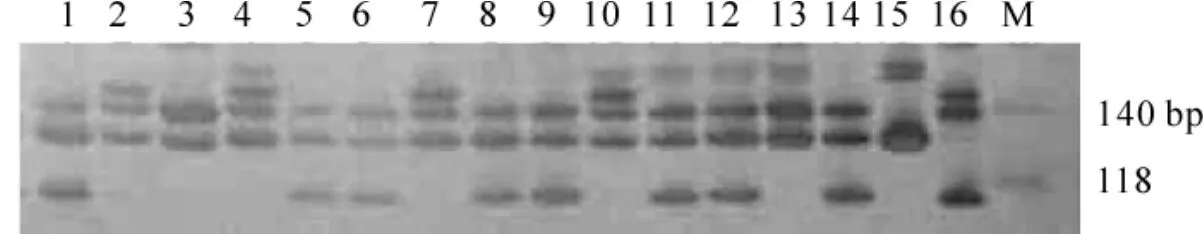
圖7 引物AB084473擴增產物在80 g·L-1聚丙烯酰胺凝膠上電泳檢測圖Figure7 Amplified product of AB084473 detecting in 80 g·L-1PAGE
克隆序列經比對后,與原物種序列相似度(max ident值)都在95%以上(表2),說明歐洲白樺和日本白樺的引物可以應用于光皮樺。體系建立后用引物AB084473對光皮樺16個植株的DNA進行擴增,得到較好的擴增結果(圖7)。
3 結論與討論
通過單因素法確定了光皮樺SSR反應體系中模板DNA用量、Mg2+濃度、dNTPs濃度、引物濃度、Taq DNA聚合酶濃度,建立光皮樺SSR反應體系。在體系建立過程中,當各個因素水平較低或較高時,條帶都比較弱,這與柳曉磊等[17]對椰子Cocos nucifera的研究結果一致。其中退火溫度是影響PCR反應的重要因素之一,實驗結果表明:較小的溫度變化對最后的擴增結果影響不明顯,但較大的溫度跨度,影響擴增條帶的多寡及其清晰程度。所選的SSR引物各解鏈溫度值相差很大,最大跨度達10℃,特別需要對每一對引物進行梯度退火實驗,以期得到各引物最佳退火溫度。
SSR分子標記因具有諸多優點,成為現代分子生物學最理想的標記之一,但其昂貴的引物開發費用,令人望而卻步。而微衛星序列的又具有保守性,使其引物在近緣中可以通用。同時在植物分類學上,光皮樺、日本白樺和歐洲白樺同屬于樺木科樺木屬,親緣關系較近,微衛星序列也趨于一致,故日本白樺和歐洲白樺的SSR引物在光皮樺中應具有較高通用性。本研究結果也表明:3個隨機選取的SSR引物在光皮樺中的擴增序列,與原物種白樺的擴增序列經比對,最大相似度值均達到95.0%以上。
[1]鄭萬鈞.中國樹木志:第2卷[M].北京:中國林業出版社,1985:2124-2131.
[2]謝一青,李志真,黃儒珠,等.光皮樺基因組DNA提取方法的比較[J].浙江林學院學報,2006,23(6):664-668.XIE Yiqing,LI Zhizhen,HUANG Ruzhu,et al.Comparison of methods of extracting genomic DNA from Betula luminifera [J].J Zhejiang For Coll,2006,23 (6):664-668.
[3]謝一青,黃儒珠,李志真,等.光皮樺RAPD分析體系優化設計方案比較[J].福建林學院學報,2006,26(1):73-77.XIE Yiqing,HUANG Ruzhu,LI Zhizhen,et al.Comparison between single factor design and uniform design to optimize the reaction system for RAPD of Betula luminifera [J].J Fujian Coll For,2006,26 (1):73 - 77.
[4]陳偉.光皮樺天然群體遺傳多樣性研究[J].北京林業大學學報,2006,28(6):28-34.CHEN Wei.Genetic diversity of the natural populations of Betula luminifera [J].J Beijing For Univ,2006,28 (6):28-34.
[5]尤衛艷,黃華宏,童再康,等.光皮樺AFLP分子標記體系的建立[J].生物技術,2008,18(6):42-45.YOU Weiyan,HUANG Huahong,TONG Zaikang,et al.Establishment of AFLP molecular labeling technique system for Betula luminifera [J].Biotechnology,2008,18 (6):42 - 45.
[6]FRAGEAU C J,FOURNEY R M.DNA typing with short tandem repeat:A sensitive and accurate approach to human identification[J].Biotchniques,1993,15:100 - 119.
[7]SHARON D,ADATO A,MHAMEED S,et al.DNA fingerprint in plant using simple sequence repeat and minisatellite probes [J].HortScience,1995,30 (1):109 - 112.
[8]宋健,韓明玉,趙彩平,等.桃‘秦光2號’ב曙光’F1代SSR遺傳連鎖圖譜的構建[J].西北植物學報,2008,28(5):895 - 900.SONG Jian,HAN Mingyu,ZHAO Caiping,et al.Construction of a general genetic linkage map for peach using a‘Qinguang 2’ × ‘Shuguang’ F1 progeny by SSR markers [J].Acta Bot Boreal-Occident Sin,2008,28 (5):895 - 900.
[9]徐小林,徐立安,黃敏仁,等.栓皮櫟天然群體SSR遺傳多樣性研究[J].遺傳,2004,26(5):683-688.XU Xiaolin,XU Li’an,HUAN Minren,et al.Genetic diversity of microsatellites (SSRs) of natural populations of Quercus variabilis [J].Hereditas,2004,26 (5):683 - 688.
[10]黃烈健,蘇曉華,張香華.與楊樹木材密度、纖維性狀相關的SSR分子標記[J].遺傳學報,2004,31(3):299-304.HUANG Liejian,SU Xiaohua,ZHANG Xianghua,et al.SSR molecular markers related to wood density and fiber traits in poplar [J].Acta Gen Sin,2004,31 (3):299 - 304.
[11]艾呈祥,張力思,魏海蓉.甜櫻桃品種SSR指紋圖譜數據庫的建立[J].中國農學通報,2007,23(5):55-58.AI Chengxiang,ZHANG Lisi,WEI Hairong,et al.Construction of molecular fingerprinting database for sweet cherry using SSR markers [J].Chin Agric Sci Bull,2007,23 (5):55 - 58.
[12]李菊芬,許玲,馬國斌.應用SSR分子標記鑒定甜瓜雜交種純度[J].農業生物技術學報,2008,16(3):494-500.LI Jufen,XU Ling,MA Guobin.Identification of melon hybrid purity by SSR markers [J].J Agric Biotechnol,2008,16 (3):494 - 500.
[13]WU B,LIAN C,HOGETSU T.Development of microsatellite markers in white birch(Betula platyphylla var.japonica)[J].Mol Ecol Notes,2002,2:413 - 415.
[14]KULJU K K,PEKKINEN M,VARVIO S.Twenty-three microsatellite primer pairs for Betula pendula (Betulaceae)[J].Mol Ecol Notes,2004,4:471 - 473.
[15]胥猛,李火根.鵝掌楸EST-SSR引物開發及通用性分析[J].分子植物育種,2008,6(3):615-618.XU Meng,LI Huogen.Development and characterization of microsatellite markers from expressed sequence tags for Liriodendron [J].Mol Plant Breed,2008,6 (3):615 - 618.
[16]李明芳.荔枝SSR標記的研究及其對部分荔枝種質的遺傳多樣性[D].儋州:華南熱帶農業大學,2003.LI Mingfang.Development of SSR Markers in Litehi chinensis and Application for Analysis of Genetic Diversity within Litchi Varieties [D].Danzhou:South China University of Tropical Agriculture,2003.
[17]柳曉磊,湯華,李東棟,等.椰子SSR反應體系的建立和優化[J].華中農業大學學報,2007,26(5):676-679.LIU Xiaolei,TANG Hua,LI Dongdong,et al.Optimization and establishment of SSR analysis method in coconut [J].J Huazhong Agric Univ,2007,26 (5):676 - 679.
