日本血吸蟲新基因Sjnanos的克隆、表達及免疫保護效果評估*
2010-08-21 10:23:14郭凡吉彭金彪傅志強石耀軍林矯矯
中國人獸共患病學報 2010年7期
關鍵詞:小鼠
王 艷,郭凡吉,彭金彪,李 曄 ,洪 煬,陳 實,傅志強,石耀軍,林矯矯
日本血吸蟲新基因Sjnanos的克隆、表達及免疫保護效果評估*
王 艷,郭凡吉,彭金彪,李 曄 ,洪 煬,陳 實,傅志強,石耀軍,林矯矯
目的擴增、表達日本血吸蟲Sjnanos編碼基因并評估其重組蛋白誘導的免疫保護效果。方法從42d齡的日本血吸蟲成蟲中擴增了一個日本血吸蟲性別差異表達的基因,GenBank登錄號為AY814948,并對其進行生物信息學分析。將該基因亞克隆入原核表達載體pET28a,轉化至大腸埃希菌(E.coli)BL21,用異丙基-β-D硫代半乳糖苷(IPTG)進行誘導表達,用純化的重組蛋白免疫小鼠,ELISA檢測其血清特異性抗體效價,蛋白印跡分析檢測其抗原性,以看家基因NADH為內參,應用熒光定量PCR技術分析該基因在血吸蟲各個階段的表達狀況。結果同源性分析表明,該基因為日本血吸蟲的一個新基因,其編碼序列的開放閱讀框(ORF)為525bp,編碼174個氨基酸,理論相對分子量為19.9,PI為8.2。熒光定量PCR技術分析顯示該基因在7d、13d、18d、23d、32d、42d蟲體均有表達,但在18d和13d蟲體的表達量明顯高于其他天數的蟲體,在雄蟲的表達量高于雌蟲。成功構建了該基因的重組表達質粒pET28a(+)-Sjnanos,并在大腸埃希菌中獲得表達,Western-blot分析表明重組蛋白具有較好的免疫原性,動物免疫保護試驗獲得了31.4%的減蟲率和53.8%的肝臟減卵率。結論獲得日本血吸蟲新的抗原基因Sjnanos,該基因的重組蛋白在小鼠中誘導了部分免疫保護作用。
日本血吸蟲;nanos基因;抗原性;免疫保護
血吸蟲病(schistosomiasis)呈全球性分布,嚴重危害人畜健康,是一種重要的人畜共患病。至2007年底,全國12個血吸蟲病流行省(市、區)的448個流行縣(市、區)中仍有105個未達到疫情控制標準,主要集中在長江流域以及洞庭湖和鄱陽湖等湖沼地區。2007年全國日本血吸蟲病病人數為51.6萬,晚期病人約2.9176萬,釘螺面積約 38億m2。
血吸蟲生活史復雜,歷經幾個發育階段,每個階段均表現出各自不同的生物學特征。尤其是血吸蟲成蟲在吸蟲中獨特的雌雄異體表現形式,使得其在生殖生理方面表現出復雜的功能差異,這已越來越引起人們的關注〔1〕。血吸蟲致病的主要原因是性成熟雌蟲大量產卵且沉積于肝臟及結腸等組織,形成蟲卵肉芽腫,造成組織免疫病理損害〔2〕。雌雄蟲合抱是雌蟲成熟和產卵的前提和關鍵,沒有合抱的雌蟲發育不全,生長遲緩,生殖系統不成熟,不能產卵,已經發育成熟的雌蟲一旦與雄蟲分離也會退化至不成熟狀態。因此,深入研究血吸蟲性別差異表達分子及其作用機制,有助于探索控制血吸蟲雌蟲產卵的有效方法〔3〕。近幾年,對血吸蟲性別差異性基因及表達產物的功能研究,已引起人們極大關注。這一領域的研究不僅對闡明血吸蟲發育及生殖機制具有重要理論意義,而且對研制抗血吸蟲生殖免疫疫苗及有效防治新藥也具有重要的實踐意義。
本文所克隆的日本血吸蟲nanos基因(Sjnanos)屬于nanos基因家族的成員之一。Nanos基因廣泛存在于果蠅、線蟲、兩棲動物類等生物中,nanos基因家族產物與其他相關基因產物是調節細胞發育和生長的一個關鍵分子,對動物的發育特別是生殖系統的發育起著重要的調節作用,但國外對血吸蟲nanos基因的研究尚未有報道。日本血吸蟲nanos基因的成功克隆,不僅使我們對nanos這類鋅指蛋白基因家族有了初步的了解,更為以后研究日本血吸蟲發育、生殖機制奠定了基礎,具有重要的生物學意義。
1 材料與方法
1.1 菌種、質粒及實驗動物 日本血吸蟲成蟲cDNA文庫由本課題組構建并保存。質粒pET28a、大腸桿菌DH5α、BL21(DE3)也由本課題組提供。新西蘭白兔(雄性,2.5~3.0kg)購自上海羅涇飛達實驗動物養殖場。BALB/c小鼠購自中國科學院上海實驗動物中心,日本血吸蟲中國大陸株尾蚴由本所釘螺室提供。
1.2 主要試劑和酶 Trizol、GeneRacerTM Kit購自Invitrogen公司;Ex T aq DNA聚合酶、限制性內切酶EcoRI、SalI、T4 DNA 連接酶、RNA inhibitor、熒光實時定量PCR檢測試劑盒購自TaKaRa生物工程(大連)有限公司;DNA Marker DL2000、DL5000,小量質粒抽提試劑盒購自上海天根生物技術有限公司,瓊脂糖凝膠電泳DNA純化回收試劑盒均購自Qiagen公司,帶His標簽的蛋白純化試劑盒購自美國 Novenge公司,Agrose、DEPC購自上海生工生物工程公司;羊抗兔 IgG-H RP、羊抗鼠IgG一HRP購自西格瑪奧里奇(上海)貿易有限公司;硝酸纖維素膜(Whatman)購自北京經科宏大生物技術有限公司;其余試劑均為國產分析純。
1.3Sjnanos基因的克隆及生物信息學分析 從本課題組構建的7d齡童蟲抑制性消減cDNA文庫獲得一段童蟲期差異表達基因EST,將該EST片段在NCBI中進行同源性比對,(http://www.ncbi.nlm.nih.gov/blast);利用DNA Star軟件對該cDNA編碼的氨基酸組成、蛋白質相對分子質量以及該蛋白的抗原性進行預測,利用ClustalW軟件對不同物種nanos蛋白氨基酸序列進行多重比對。用GeneDoc軟件對比對結果進行編輯。以該EST序列設計上游引物P1:CGCAGGACT TTCAGACTC TAG;下游引物 P2:GTAGTGAAGTAGGCGTAATGAC,利用PCR技術擴增含ORF的cDNA片段和基因組序列片段。
1.4 實時定量PCR 分別提取日本血吸蟲7、13、18、23、32d蟲體及42d雄蟲與雌蟲的蟲體總RNA,消化基因組DNA,定量,反轉錄,純化回收。以純化的不同期別的日本血吸蟲cDNA為模板,設計引物:Sense primer:5'-ATGGCAACAAGGACCTGATG-3',Antisense primer:5'-TCACTAAGGCTGT TCCAATCG-3',用熒光染料法進行實時定量PCR檢測Sjnanos基因在不同時期蟲體的表達量,以日本血吸蟲NADH基因為內參,NADH引物Sense primer:CGAGG ACC TAA CAG CAG AGG,Antisense primer:TCCGA ACG AAC TT T GAA T TC,引物均由上海英俊生物技術有限公司合成,反應條件為:95℃10s,然后 95℃5s,55℃10s,72℃15s,40個循環,其中72℃15s結束時間為熒光信號檢測點,每個反應均做3孔重復。用cobert公司rotogene6.0軟件進行分析,得出相對每百萬持家基因的目的基因含量。
1.5 重組表達質粒的構建與表達
1.5.1 pET28a(+)-Sjnanos重組質粒的構建及鑒定 根據Sjnanos基因的cDNA序列設計引物,Sense Primer:5'-GC GAAT TC ATG ACT TCA ATC GAC-3';Antisense Primer:5'-GTCGAC CTA AAA T TG CCT GGG-3',分別引入限制性酶切位點EcoR I、SalI(劃底線部分),引物由上海英俊生物技術有限公司合成,通過PCR擴增得到含完整ORF的Sjnanos基因序列,該序列與經過同樣雙酶切的載體pET28a(+)連接,將該序列定向克隆于原核表達載體pET28a(+)的多克隆位點區,構建重組質粒 pET28a(+)-Sjnanos,并轉化表達菌BL21(DH5α)。將陽性重組菌接種 LB中擴大培養,提取重組質粒,進行PCR雙酶切和DNA序列鑒定。
1.5.2 在大腸桿菌中的表達 將測序正確的表達重組質粒接種于含100μ g/mL的卡那霉素LB液體培養基,37℃震蕩培養,當OD600=1.0時加入終濃度為 1mmol/L 的異丙基-β-D-硫代半乳糖苷(IPTG)進行誘導表達,分別收集誘導前及誘導后1、2、3、4、5、6 、7h 的菌液以分析確定最佳表達時相。同時以載體pET28a(+)誘導前后及未誘導重組質粒作為對照,SDS-PAGE電泳,凝膠經過考馬斯亮藍染色、脫色后分析表達結果。
1.6 重組Sjnanos蛋白的純化及復性 pET28a(+)-Sjnanos按上述方法進行大量誘導表達,然后對誘導菌作如下處理:10 000 g離心10 min,每100 mL菌液用4 mL 1×PBS重懸。-80℃反復凍融3次后,冰浴超聲30min,其間每超 5s停 10s。10 000 g離心 15 min,棄上清,沉淀用20mL 8mol/L尿素溶解。10 000 g離心15 min,收集上清即為包涵體蛋白。上清用0.45μ m 的濾膜過濾,按照Ni-NTA agarose包涵體純化試劑盒說明純化重組蛋白,SDS-PAGE電泳結果顯示,重組蛋白純度可達90%以上,過柱純化后的蛋白經透析復性后,紫外分光光度計測定蛋白濃度,-80℃保存備用。
1.7 純化重組蛋白Sjnanos的抗原性鑒定 將純化的重組蛋白經SDS-PAGE電泳,然后電轉移(280mA不超過3h)至硝酸纖維素膜(NC膜),電轉結束后,取出NC膜放入封閉液(5%脫脂奶粉/PBST),室溫 2h或 4℃過夜,用經 pET28a(+)/BL21大腸桿菌蛋白吸附的日本血吸蟲成蟲抗原免疫兔血清做一抗,辣根過氧化物酶標記的羊抗兔IgG做二抗,最后加入沉淀型TMB顯色液進行顯色,拍照保存。
1.8 小鼠免疫保護實驗及血清采集 購置6-8w齡(SPF級)BALB/c小鼠 30只,隨機分成 3組,每組10只,分別為免疫組、佐劑對照組和空白對照組。免疫組每次每只注射含206佐劑和27μ g純化的Sjnanos重組蛋白的乳化液;佐劑對照組每次每只注射相同體積的206佐劑和PBS的乳化液;空白對照組每次每只注射相同體積的PBS。免疫間隔時間為2w,每次免疫后1w尾靜脈采血,收集血清,共免疫3次。第3次免疫后2w,每只小鼠經腹部皮膚貼片攻擊感染40±1條日本血吸蟲尾蚴。攻蟲42d后,剖殺小鼠,收集血清,以肝門靜脈灌注法收集蟲體,計數蟲體,計算減蟲率,肝臟減卵率和糞便減卵率。
1.9 ELISA試驗檢測特異性IgG抗體 將免疫前、每次免疫后1w及剖殺的實驗血清作1∶100稀釋后做常規 ELISA實驗〔4-5〕,檢測Sjnanos特異性抗體。
2 結 果
2.1Sjnanos基因的克隆及生物信息學分析 從本課題組構建的7d童蟲消減cDNA文庫中,獲得一EST序列,利用NCBI的Blast軟件對該基因進行同源性搜索,結果顯示該基因與血吸蟲其他已知基因無顯著同源性,為日本血吸蟲的新基因,氨基酸序列相似性比較表明該基因編碼蛋白的結構域與多個物種的ZF-nanos蛋白結構功能域相似,命名為Sjnanos。DNA Star分析表明該序列長721bp,其中含ORF為525bp,編碼175個氨基酸,理論相對分子量為 19 905.71,理論等電點 PI為 8.2,Sjnanos蛋白的N端不具有信號肽,無跨膜區,但疏水性分布較均勻,有較強的抗原性,見圖3。選擇分別來自曼氏血吸蟲(GenBank登錄號 ref|XP_002576493)、渦蟲(GenBank登錄號:AB052809)、家蠶(GenBank登錄號:ref|NP_0010987021)、水螅屬(GenBank登錄號:ref|XP_002159764.1)、耳鮑(GenBank登錄號:ACT35656.1)、小家鼠(Gen-Bank登錄號:AAH56473.2)和人(GenBank登錄號:ref|NP_955631.1)7個物種的nanos蛋白進行氨基酸序列的多重比對,結果顯示其編碼的氨基酸序列與曼氏血吸蟲Nanos的相似性為67%,與人的相似性為 56%,與家蠶、水螅、小鼠、渦蟲等的相似性分別為 58.0%、40.0%、53%、45.0%等,見圖 1。
2.2 重組原核表達質粒 pET28a(+)-Sjnanos的構建經 PCR、雙酶切鑒定(圖 4)和測序,證明pET28a(+)-Sjnanos重組表達質粒構建成功。
2.3Sjnanos基因的期別、性別差異表達分析 Real-time PCR分析表明,Sjnanos基因在童蟲及成蟲階段均有表達,其中在18d蟲體的表達量最高,隨后依次為13d、23d、42d蟲體,7d和32d蟲體表達量較少。
以 42d雌、雄蟲cDNA為模板進行real-time PCR,結果表明,Sjnanos在雌、雄蟲中均有表達,但在雄蟲中的表達量較高,說明Sjnanos為雄蟲高表達基因。

圖1不同物種nanos蛋白氨基酸序列的多重比對(clustal W)100%同源的用 *表示,80%同源的用 :表示Fig.1 Aminoacid sequence alignment of nanos orthologues(clustalW)100%identical residues are marked with*,80%identical with:

圖2 不同物種nanos的氨基酸序列構建的系統進化樹Fig.2 The Phylogenetic tree analysis of Sjnanos with the homologues(the accession numbers of the other members are cited in the legend of fig.1).
2.4Sjnanos在大腸桿菌中的表達和重組蛋白的純化 SDS-PAGE結果顯示,Sjnanos在BL21中獲得表達,融合蛋白的相對分子量為21kD,誘導時間4h表達量達到最大(圖8),該重組蛋白以包涵體形式存在,溶解于8mol/L尿素,經NTA-Ni樹脂純化,獲得純化后的重組蛋白(圖9)。
2.5 表達產物的抗原性分析 重組蛋白SDSPAGE電泳后,經電轉移至NC膜上,用日本血吸蟲成蟲抗原免疫兔血清作為一抗,辣根過氧化物酶標記的抗兔IgG做二抗,Western blot檢測,結果有一明顯的識別條帶(圖10),表明重組蛋白具有良好的抗原性。

圖3 Sjnanos蛋白的疏水性分析Fig.3 Analysis the hydrophobicity of Sjnanos protein

2.6 動物保護實驗 與空白對照組相比,重組蛋白在BALB/c小鼠中誘導了31.4%減蟲率,t檢驗分析表明免疫組和空白對照組差異顯著(P=0.00761)。肝臟經消化處理后,在顯微鏡下進行蟲卵計數,與空白對照組相比,肝臟減卵率為53.8%,t檢驗分析表明,免疫組與空白對照組差異極顯著(P=0.00227),與空白對照組相比,佐劑組雖也一定的減蟲及減卵效果,但其P值分別為0.071484、0074063,都大于0.05,與空白組差異不顯著。
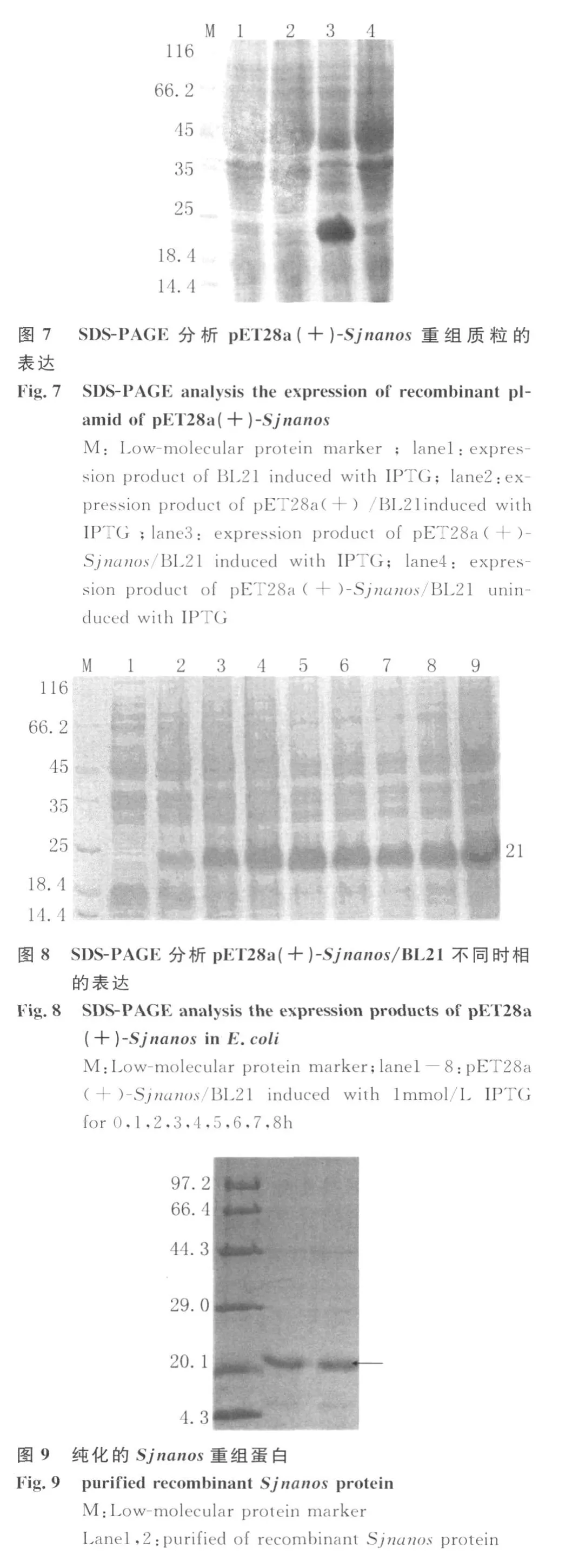

圖10 pET28a(+)-Sjnanos重組蛋白western bloting分析Fig.10 Western blotinganalysis of pET28a(+)-Sjnanos recombinant protein
2.7 特異性IgG抗體檢測 各組小鼠免疫血清中抗Sjnanos特異性IgG抗體的動態變化如圖11,第二次免疫后7d,特異性抗Sjnanos IgG抗體明顯上升,第三次免疫后7d,抗體滴度達到最高,而空白對照組及佐劑對照組特異性IgG抗體在整個免疫過程中一直維持在較低水平,無顯著變化。

圖11 Balb/c小鼠血清抗Sjnanos抗原特異性IgG抗體水平檢測結果Fig.11 Specific IgG level against Sjnanos by ELISA

表1 BALB/c小鼠免疫保護試驗結果Tab.1 Worm and egg counting reduction rate induced by Sjnanos in Balb/c mice
3 討 論
血吸蟲病是一種分布廣泛,危害嚴重的人畜共患病。血吸蟲為吸蟲中罕見的雌雄異體,性別的分化對血吸蟲的發育及致病起著至關重要的作用。雌雄蟲合抱是血吸蟲雌蟲發育和成熟、產卵的關鍵〔6〕。雖然蟲體在動物體內不能夠增殖,但雌蟲成熟后產生大量蟲卵寄生在人體及動物肝臟和腸組織形成的肉芽腫是導致發病的主要原因〔7〕。因此,加強對血吸蟲性別相關基因的研究具有重要的理論意義和實際應用價值,可為血吸疫苗的研究提供線索和理論依據。
Nanos是最先在果蠅的生殖顆粒中被鑒定的一個母源效應組分,其編碼一種鋅指結構蛋白,鋅指蛋白是指一類含有Zn2+的“手指”結構的一類蛋白質,最初于1983年在非洲爪蟾卵母細胞的轉錄因子TFⅢA 中被發現〔8-9〕,廣泛分布在動物、植物和微生物中,人類基因組中可能有將近1%的序列編碼的是含有鋅指結構的蛋白質。由于鋅指蛋白自身的結構特點,使其在基因表達調控、細胞分化、胚胎發育、增強植物抗逆性等方面具有重要作用〔10〕。該蛋白的C端含有兩個高度保守的由CCHC組成的鋅指結構,在這個結構域外幾乎不含保守區〔11〕,此蛋白可以特異性地結合RNA,nanos家族蛋白的主要功能是抑制和調節生殖細胞的分化、發育和成熟〔10-11〕,在昆蟲胚胎后部分區域的特殊分化中起重要作用。經研究證實,nanos蛋白是生殖細胞發育和再生所必須的。同時,有學者推斷,在果蠅中,從早期的卵裂期到成蟲的整個發育過程中,nanos蛋白與vasa蛋白都在生殖細胞中表達,共同調節個體的發育與生殖細胞的成熟。nanos蛋白在早期胚胎中有兩個功能:第一,nanos蛋白與 RNA結合蛋白pumilio一起抑制母源 mRNA hunchback的翻譯〔13-14〕。第二,nanos蛋白對原始生殖細胞的發育是必不可少的,母源nanos蛋白的缺失導致原始生殖細胞不能正常移動到性腺,從而產生無功能的生殖細胞〔15〕。
研究結果表明,Sjnanos在 13d、18d、23d的表達量顯著高于7d、32d、42d,這與日本血吸蟲生長發育過程是相吻合的。日本血吸蟲從14d開始合抱,合抱以后睪丸和卵巢逐漸發育和成熟,nanos基因可以調節個體發育及生殖細胞的形成,該基因在13d、18d、23d等高表達可能在日本血吸蟲的生長發育,特別是生殖器官的發育成熟中起到重要的作用。先前的研究表明,應用RNAi的技術敲除nanos基因,進行有性生殖的渦蟲不能產生和形成性腺〔16〕。同樣,在進行無性生殖(分裂生殖)的渦蟲體內通過RNAi手段敲除nanos基因后,分裂生殖的渦蟲不能正常生殖,渦蟲可能不能形成正常的生殖細胞,進而影響了生殖。
本研究大量表達純化Sjnanos融合蛋白,并進行了小鼠日本血吸蟲病免疫保護實驗,初步的實驗結果表明,融合蛋白Sjnanos在BALB/c小鼠中誘導了31.4%減蟲率和53.8%的肝臟減卵率,且與空白對照組相比差異極顯著,說明Sjnanos蛋白作為候選疫苗分子可誘導產生部分的保護效果,但這只是初步試驗結果,有待于進一步重復驗證。實際上影響動物保護效果的因素除了抗原本身的性質,還與免疫劑量、注射途徑以及佐劑量、佐劑種類等因素相關〔17-18〕,如本次試驗的佐劑組的結果稍稍偏高,所以在佐劑種類的選擇及佐劑的量方面還需進一步摸索。同時由于該蛋白是日本血吸蟲童蟲期特異表達產物,產生的抗體可能對成蟲期的影響不大,成蟲可在宿主體內生長發育。
本研究首次克隆獲得了日本血吸蟲nanos基因,為探索nanos基因家族在血吸蟲生長發育特別是生殖器官發育中過程中的作用及開發高效的抗血吸蟲病的疫苗和篩選新型的抗血吸蟲藥物奠定基礎。
〔1〕Schussler P,Grevelding CG,Kunz W.Identification of Ras,and aGAP protein inSchistosoma mansoniby immunoblotting and their putative involvement in male2female interaction〔J〕 .Parasitology,1997,115:629-634.
〔2〕Basch PF.Cultivation ofSchistosoma mansoni in vitroⅡproduction of infertile eggs by worm pairs from cercariae〔J〕 .Parasitol,1981,67:186-193.
〔3〕Hirai,H.M.T anaka,T.L.Philip,Schistosoma mansoni:chromosomal location of female-specific gene and a female-specific DNA element〔J〕 .Ex perimental Parasitol,1993,76:175-181.
〔4〕薩姆布魯克,弗里奇,曼尼阿蒂斯,分子克隆實驗指南〔M〕.2版.北京:科學出版社,2002.
〔5〕ZhuY,RenJ,Ham DA,et al.Protective immunity induced with 23 kDa membrane Protein DNA vaccine ofSchistosoma japonicumChinese strain in infected C57BL/6 mice〔J〕.Southeast Asian Trop M ed Pub Hehh,2003,34(4):697-701
〔6〕Popiel I,Basch PF.Reproductive development of femaleSchistosoma mansoni(Digenea:Schisto-somatidae)following bisexual pairing of worms and worm segments〔J〕.T he Journal of Experimental Zoology,1984,232(1):141-150.
〔7〕Muschamp JW,Fong PP.Effects of the serotonin receptor ligand methiothepin on reproductive behavior of the freshwater snail Biomphalaria glabrata:reduction of egg laying and induction of penile erection〔J〕.The Journal of Experimental Zoology,2001,289(3):202-207.
〔8〕Miller J,McLaehlan A D,Klug A.Repetitiv zinc-binding domains in the protein transcription factorⅢA from Xenopus oocytes〔J〕.EMBO J,1985,4:1609-1614.
〔9〕Lee M S,Gippert G P,Soman K V,et al.T hree-dimensional solution structure of a single zinc finger DNA-binding domain〔J〕.Science,1989,245:635-637.
〔10〕Calvo E,Walter M,Adelman ZN,et al.Nanos(nos)genes of the vector mosquitoes,Ancphelesgambiae,AnophelesstephensiandAedes aegy pti〔J〕.Insect Biochem,2005.
〔11〕Curtis D,T reiber DK,T ao F,et al.A CCHC metal-binding domain in Nanos is essential fortranslational regulation〔J〕.EMBO J,1997,16:834-843.
〔12〕Extavour CG,PangK,Matus DQ,MartindaleM Q.vasa and nanos expression patterns in a sea anemone and the evolution of bilaterian germ cell specification mechanisms〔J〕.Evol Dev,2005,7:201-215.
〔13〕Hulskamp M,Schroder C,Pfeifle C,et al.Posterior segmentation of the Drosophila embryo in the absence of a maternal posterior organizer gene〔J〕 .Nature,1989,338(6217):629-632.
〔14〕Irish V,Lehmann R,Akam M.The Droscphila posterior-g roup gene nanos function by rep-ressing hunckback activity〔J〕.Nature,1989,338(6217):646-648.
〔15〕Kobay ashi S,Yamada M,Asaoka M,et al.Essential role of the posteriormo rphogen nanos for germline development in Droscphila Nature,1996,380(6576):708-711.
〔16〕Wang Y Y,Zay as R M,Guo TX,et al.Newmark nanos function is essential fo r development and regeneration of planarian germ cells〔J〕 .PN AS,2007,104(14):5901-5906.
〔17〕鄭武飛.醫學免疫學〔M〕.北京:人民衛生出版社,1989:18-33.
〔18〕沈關心,周汝麟.現代免疫學實驗技術〔M〕.湖北科學技術出版社,1998:16-39.
Cloning,expression and characterization of a novel gene ofSchistosoma japonicum
WANG Yan,GUO Fan-ji,PENG Jin-biao,LI Ye,HONG Yang,CHEN Shi,FU Zhi-qiang,SHI Yao-jun,LIN Jiao-jiao
(Shanghai Veterinary Research Institute,Chinese Academy of Agricultural Science,Key Laboratory for Animal Parasitology,Ministry of Agriculture,Shanghai200241,China)
To clone and express the gene(nanos)ofSchistosoma japonicum,and study the antigenicity and immunogenicity of the protein,the nanos gene amplified by PCR from worms'cDNA was cloned into pronucleus expression vector pET28a(+)to construct pET28a(+)-Sjnanosrecombinant plasmid and transformed intoE.coliBL21(DE3).In the presence of IPTG,the 21KD protein was expressed and purified under denaturing conditions to immune the BALB/c mice.The gene sequencing data showed that length of the whole ORF was 525bp and encoded 174 amino acids with a prediction of 21KD.Results of Western blot and ELISA revealed that the fusion protein had good immunogenicity.Real-time PCR analysis showed the mRNA transcription level of nanos was much higher in 13d,18d and 23d within Schistosomula than in other stages.After immunizing BALB/c mice with fusion protein,the percentage of worm and liver egg reduction was 31.4%and 53.8%,respectively.The gene ofSjnanoswas cloned for the first time,and the successful expression and purification of pET28a(+)-Sjnanoswould be helpful for the further protection in animals.
Schistosoma japonicum;nanos;clone;fusion protein
R383.2
A
1002-2694(2010)07-0631-07
*國家自然科學基金(30671581),973課題(2007CB513108)和863計劃(2006AA10A207)聯合資助
林矯矯,Email:wangyan0516@163.com
中國農業科學院上海獸醫研究所,農業部動物寄生蟲學重點開放實驗室,國家防治動物血吸蟲病專業實驗室,上海 200241;
2009-11-25;
2010-03-22
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
