大腸埃希菌外排泵AcrAB-TolC研究相關進展
2010-09-07 06:30:46旭綜述黃永茂審校
重慶醫學 2010年19期
關鍵詞:耐藥
王 旭綜述,黃永茂審校
(瀘州醫學院附屬醫院感染科,四川瀘州646000)
大腸埃希菌外排泵AcrAB-TolC研究相關進展
王 旭綜述,黃永茂審校
(瀘州醫學院附屬醫院感染科,四川瀘州646000)
大腸埃希菌;耐藥;外排泵
細菌的外排系統是一種非特異性耐藥機制,是通過細菌外排泵將進入菌體內的藥物或其他底物排出膜外,它可以泵出多種對其自身有害的物質,包括喹諾酮類、氯霉素、紅霉素、四環素、青霉素、利福平等多種抗菌藥物、染料和去污劑等,從而加強細菌在藥物選擇壓力下的生存能力。目前,已在不同細菌上發現幾十種外排泵,依據氨基酸序列的同源性,將與抗菌藥物相關的膜外排泵分子分為5個主要超家族[1],包括主要易化子超家族(major facilitato r superfamily,M FS)、A TP結合盒(A TP binding cassette,ABC)超家族、耐藥節結化細胞分化(resistance nodulation division,RND)超家族、小多重耐藥性(smallmultidrug resistance,SMR)家族、多藥和有毒化合物排出(multidrug and toxic compound extrusion,MA TE)家族。按能量來源不同可分為兩大類:ABC型多藥外排系統和次級多藥外排系統。大多數抗菌藥物的外排系統都屬于次級外排系統,多數細菌通過質子驅動力來表達對不同結構化合物的耐藥。
1980年Bah和M cM urry在研究大腸埃希菌對四環素的耐藥性時發現了主動外排耐藥機制,此后人們對細菌特別是大腸埃希菌的外排耐藥系統有了進一步的認識。大腸埃希菌是已發現的主動外排泵最多的一種細菌,存在Bcr、Em rE、EmrAB、Em rD、QacE、AcrAB-TolC、AcrEF、AcrD、YdhV、M dfA等多種外排泵。RND中的AcrAB-TolC蛋白是大腸埃希菌多藥耐藥外排系統中目前研究比較清楚的一種細菌外排泵。用基因敲除技術人工去除AcrAB或TolC基因后,大腸埃希菌對上述藥物的敏感性明顯增加[2]。
1 外排泵AcrAB-TolC的結構和功能
AcrAB-TolC系統主要有3個部分:膜融合蛋白(AcrA)、外排轉運蛋白(AcrB)和外膜通道蛋白(TolC)。AcrA、AcrB、TolC以三聚體形式存在于大腸埃希菌細胞膜上,其中AcrA位于周質間隙中,兩端連接著AcrB和TolC;AcrB和TolC分別位于細胞內膜和外膜上。迄今所知,AcrAB-TolC系統可以提供2種途徑對藥物實現外排,一種是捕獲胞質內的藥物,直接穿越雙層膜將藥物排出,比如該系統介導對氯霉素和四環素等膜滲透性藥物的耐藥;另一種是捕獲周質間隙中的藥物分子,通過TolC將藥物轉運到外界[2]。
1.1 AcrA蛋白 AcrA是一個由398個氨基酸殘基組成的蛋白質,相對分子質量為41 kD,屬膜融合蛋白(membranefusion p rotein,M FP)超家族成員,其脂質化的N端錨釘在內膜上,C端伸展到周質[3]。在周質中,AcrA呈高度不對稱,蛋白質的N端和C端不相互接觸,而是分別位于兩端,從而連接AcrB和TolC形成復合物[4]。AcrA和AcrB位于同一操縱子,兩基因之間無終止信號。在大腸埃希菌中除AcrB外,AcrA可以與AcrD、AcrF和YhiV等多個RND家族蛋白作用。AcrA的C端區域290~357的氨基酸殘基對于它和AcrB間作用是必須的,這段區域的局部序列特征對決定AcrA與AcrB的作用非常重要[4]。AcrA除了作為橋梁連接AcrB和TolC外,也可以有效激活AcrB。Lobedanz等[4]發現,取代AcrA的357~397氨基酸殘基的編碼序列,仍然能夠產生一種功能性蛋白,但并不能使外排功能完全喪失,從而證實AcrA的290~357殘基區域在AcrB的激活。
1.2 AcrB蛋白 AcrB由1 048個氨基酸殘基組成,相對分子質量為110 kD,屬RND家族。AcrB包括12個親脂區和2個親水片段;2個親水片段是AcrB轉運子所特有。射線衍射分析顯示,AcrB以同三聚體形式橫跨細胞內膜,在內膜的外側形成漏斗形結構,開口于細胞周質,能直接攝取細胞周質間隙中的藥物,在內膜的內側形成一個直徑為30?的中央腔,開口向細胞質,漏斗形結構和中央腔相連的部分是個狹窄或關閉的孔道[5]。在對AcrB氨基酸殘基的分析中,研究人員發現了與底物結合以及質子轉移供能相關的位點。在T(tight)狀態時, AcrB 2個周質結構域上的F136、178、610、615、617和628; V 139、612;I277、626和位于孔洞結構域的Y327構成的疏水區域,一些化學性質不同的物質如羅丹明6G和溴化乙錠(ethidium bromide)等可以進入結合洞穴,通過疏水鍵與中央洞穴結合[6]。
1.3 TolC蛋白 TolC由495個氨基酸組成的三聚體,上端為開放結構,以便給外排底物提供泵出寬闊通道;下端為封閉結構,其中包括一個10 nm的位于細胞周質的一個螺旋孔道和一個4 nm跨越外膜的筒狀結構。它是一個由TolC基因編碼的多功能的TolC,可以與多種不同的轉運蛋白相耦合,在轉運過程中幾乎不決定轉運的特異性和方向[7]。TolC為三聚體,蛋白質一部分以螺旋結構進入周質與AcrA連接,當內膜AcrB捕獲到細胞內的有害物質并與之結合時,TolC與AcrAB復合體連接,打開內在通道,將有害物質排出。基因研究證實, TolC是AcrAB實現其功能不可或缺的部分,TolC的作用主要是為藥物分子通過細菌外膜提供途徑(圖1)。
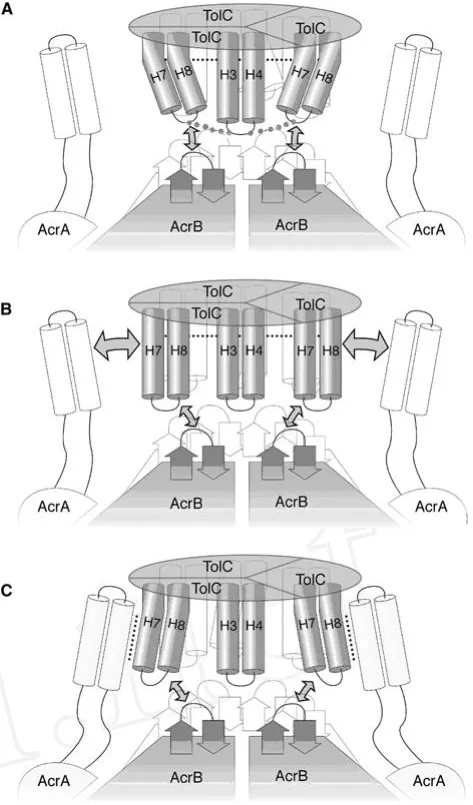
圖1 大腸埃希菌外排泵AcrAB-TolC結構和功能示意圖
2 AcrAB-TolC的基因調控
AcrAB的表達受多種調控因子的調節,包括局部和全局兩個層次調控。局部調控常常是在緊鄰其基因的位置上表達出轉錄抑制或激活蛋白,以調控該泵蛋白的表達。大腸埃希菌多藥外排泵AcrAB-TolC系統中,AcrA和AcrB蛋白的基因位于同一個操縱元上,AcrA上游存在著一個調控基因,AcrA屬于局部調控基因[8-10]。全局性的調控則往往是由細胞內一些整體性調控因子參加的與細胞整體代謝變化相關聯的調控機制。
2.1 局部調控因子 局部調控因子AcrR位于轉運子基因上游,其編碼產物為N末端含有超螺旋結構阻遏調控蛋白。AcrR為負調控蛋白,AcrR作用可以阻遏AcrAB的外排作用,其基因突變可以使AcrAB表達增強,同時AcrR也可以抑制自身的啟動子而調控自己的表達,從而使耐藥性增強[11]。Webber等[10]對于不同來源的抗氟喹諾(nuoroquinolone)大腸埃希菌突變型進行分析發現,它們的AcrB過量表達并且AcrR第45位氨基酸有突變,利用野生型AcrR對此種突變型進行互補實驗發現,互補會導致菌株對環丙沙星(cip rofloxacin)和溴化乙錠敏感性增加。在人工突變研究中,Ma將大腸埃希菌Ac-rR插入失活后,AcrAB的轉錄水平隨之提高;將AcrR連接到pACYC177上讓其過量表達,會抑制AcrAB的轉錄。
2.2 整體調控因子 整體調控因子負責誘導這個外排泵系統,增加AcrAB和TolC的轉錄,屬于正調控。目前發現,參與正調控的因子有MarA、Rob、SoxS和Fis等[12]。
2.2.1 全局性調控蛋白(MarA) MarA是多重抗菌藥物耐性(multiple antibio tic resistance,M ar)操縱元的轉錄激活蛋白,由125個氨基酸組成的正調控因子。它可結合到AcrAB啟動子附近,增強RNA聚合酶與該啟動子的親合力,促進AcrAB的轉錄,它也能以同樣的方式提高TolC的表達。MarA亦可提高自身的轉錄水平,MarA作為一個單體可與MarRAB啟動子上方MarO連結,激活MarRAB的轉錄,提高MarA在細胞內的表達量,隨后與MarA調節基因啟動子鄰近的AcrAB和TolC結合,促進其轉錄[13]。
2.2.2 其他正調控蛋白(SoxS) SoxS是整體超氧化反應(superoxide response,SOX)調控元SoxRS的效應蛋白,是由107個氨基酸組成的小分子蛋白質。蛋白組學和遺傳學常規分析結果顯示SoxS可激活17個基因或操縱子的表達,過氧化物(O2-)可以使SoxR效應蛋白轉變成它的活化形式(SoxR*),活化的SoxR進而會促進SoxS的產生[13];Rob是由289個氨基酸組成的能結合低分子量效應基因單獨區域的較大蛋白質,大腸埃希菌中Rob的無意義突變可增加其對有機溶劑的敏感性,過量表達則增加其耐受性[14]。輔助激活蛋白Fis參與到重組和DNA修復過程,穩定某些啟動子的局部DNA結構,對應不同的生長條件而改變其轉錄活性,它也可以促進AcrAB轉錄。
3 大腸埃希菌外排泵抑制劑的探索
隨著抗菌藥物的大量使用,大腸埃希菌在選擇性壓力下不斷發展其耐藥機制,造成日益嚴重的耐藥問題。探索解決其耐藥方法已經刻不容緩,根椐對大腸埃希菌外排系統的了解和研究,可以從以下2個方面著手。
3.1 干擾外排泵組裝的抑制劑 Globomycin是一種鏈霉菌來源的環肽結構的抗菌藥物,它是脂蛋白信號肽酶LspA的抑制劑,通過抑制LspA從而抑制含有L spA剪接位點的膜融合脂蛋白前體的加工。大腸埃希菌和產氣腸桿菌組成表達RND型外排泵AcrAB-TolC的組件AcrA前體且含有L spA的剪接位點,在Globomycin的作用下不能形成成熟的AcrA,也不能和外排泵的其他組分結合形成外排泵,抑制了外排泵的作用。文獻報道75μmol/L Globom ycin能夠使表達AcrAB-TolC的產氣腸桿菌的氯霉素最低抑菌濃度(M IC)降低至原值的1/4,其外排泵抑制能力與羰酰氰間氯苯腙(carbonyl cyanidemchlorophenylhydrazone,CCCP)(100μmol/L)相同,低于PAβN (100μmol/L,M IC降低至原值的1/8)。
3.2 阻斷外排泵能量來源的抑制劑 大腸埃希菌的外排泵主要由質子動力勢驅動。對細菌多重耐藥外排泵抑制劑的研究發現,以阻斷外排泵能量來源為機制的外排泵抑制劑主要針對質子動力勢[15]。例如CCCP其分子在解離狀態下,導致轉運蛋白失去能量供應,破壞外排系統的主動外排作用,使藥物在細菌體內的累積量增加,恢復細菌對藥物的敏感性,因此,它是很強的解耦聯劑。CCCP作為外排泵抑制劑對大腸埃希菌AcrAB和AcrEF、產氣腸桿菌AcrAB-TolC、空腸彎曲桿菌CmeABC等均有抑制作用。例如,100μmol/L的CCCP可以使表達AcrAB-TolC的產氣腸桿菌的氯霉素M IC降低至原值的1/4[16]。
4 展 望
目前,對大腸埃希菌的外排泵的結構、調控方式及如何抑制它的外排泵有了一定的認識和了解,但是研究僅限于體外實驗,并且這些抑制劑大多對人體有不良反應。許多在體外活性良好的外排泵抑制劑由于安全性、特異性、機制明確性等諸多問題未能用于臨床治療。外排泵抑制劑不僅可以提高具有外排泵介導耐藥性的病原細菌的藥物敏感性,恢復抗菌藥物的抗菌活性,還有利于減少由外排作用促進的耐藥突變株的產生。除了對外排泵抑制劑的研究還需要臨床工作者注意合理使用抗菌藥物。(1)根據體外檢測藥敏結果使用抗菌藥物,對患者實行個體化用藥;(2)抗菌藥物的劑量要適當,注意由于劑量不足而造成耐藥性的產生,療程應盡量短;(3)根據細菌耐藥和發展趨勢,有計劃地將抗菌藥物分批分期交替使用可能是一項重要措施;(4)在醫院內嚴格執行消毒隔離制度,以防耐藥菌的交叉感染[17]。因此,尋找有應用前景的外排泵抑制劑并合理使用抗菌藥物對于細菌性感染的治療有著深遠的意義。
[1]Poole K.Efflux-mediated antimierobial resistance[J].J Antimicrob Chemother,2005,56(1):20.
[2]Touze T,Eswaran J,Bokma E,et al.Interactions underlying assembly of the escherichia coli AcrAB-TolC multidrug efflux system[J].JMol M icrobiol,2004,53(2):697.
[3]M ikolosko J,Bobyk K,Zgurskaya H I,et a1.Confo rmational flexibility in the multidrug efflux system p ro tein AcrA[J].Structure,2006,14(3):577.
[4]Lobedanz S,Bokma E,Symmons M F,et al.A perip lasmic coiled-coil interface underlying TolC recruitment and the assembly of bacterial drug efflux pumps[J].Proc Natl Acad Sci USA,2007,104:4612.
[5]Deniaud A,Goulielmakis A,Covès J,et al.Differences between CusA and AcrB crystallisation highlighted by p rotein flexibility[J].PLoSOne,2009,4(7):e6214.
[6]Seeger MA,Schiefner A,Eicher T,et a1.Structural asymmetry of AcrB tfimer suggests a peristaltic pump mechanism[J].Science,2006,313:1295.
[7]Higgins CF.Multip le molecular mechanism s fo r multidrug resistance transporters[J].Nature,2007,446:749.
[8]Ramos JL,Martlnez,Bueno M,Molina-Herares AJ,et al. The TetR family of transcrip tional rep resso rs[J].M icrobiol Mol Biol Rev,2005,69(2):326.
[9]Li M,Gu R,Su CC,et a1.Crystal structure of the transcriptional regulato r AcrR from Escherichia coli[J].JMol Biol,2007,373:591.
[10]Webber MA,Talukder A,Piddock LJV.Contribution of mutation at amino acid 45 of AcrR to AcrB exp ression and cipmfloxacin resistance in clinical and veterinary Escherichia coli isolates[J].Antimicrob Agents Che, 2005,49(10):4390.
[11]Herve N,Vincent P,Stuart B.Levy,increased genome instability in escherichia coli ion mutants:relation to emergence of multip le-antibio tic-resistant(Mar)mutants caused by insertion sequence elements and large tandem [J].Antimic Agents Chem,2007,51(4):1293.
[12]Eiji Nikaido,Akihito Y,Kunihiko N.AcrAB m ultidrug efflux pump regulation in salmonella enterica serovar ty-phimurium by RamA in response to environmental sig-Snals[J].JBio Chem,2008,283(35):24245.
[13]David K,A lexey R.MarA-mediated overexp ression of the AcrAB efflux pump results in decreased suscep tibility to tigecycline in Escherichia coli[J].J Antimicrobial Chemotherapy,2008,61:46.
[14]Taketo K,Kameino L,Fujisawa I.High hydrostatic p ressure treatment impairs AcrAB-TolC pump resulting in differential loss of deoxycholate Tolerance in Escherichia coli[J].J Bio Bioeng,2005,100:6.
[15]Paix?o L,Rodrigues L,Couto I,et al.Fluorometric determination of ethidium bromide efflux kinetics in Escherichia coli[J].J Biol Eng,2009,3:18.
[16]Mallea M,Chevalier J,Eyraud A,et a1.Inhibito rsof antibiotic efflux pump in resistant Enterobacter aerogenesstrains[J].Biochem Biophys Res Commun,2002,293 (5):1370.
[17]周安宇.746例痰標本中的病原菌分布及耐藥性分析[J].重慶醫學,2008,37(11):1327.
book=6,ebook=307
10.3969/j.issn.1671-8348.2010.19.057
R378.21;R969.3
A 文獻標識碼:1671-8348(2010)19-2670-04
2010-04-10
2010-05-28)
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
