交替假單胞菌LP621菌株產(chǎn)右旋糖苷酶的培養(yǎng)條件優(yōu)化
2010-09-25 09:30:44呂明生王淑軍房耀維焦豫良吳彬彬
微生物學雜志 2010年6期
關(guān)鍵詞:生長
呂明生,王淑軍*,房耀維,焦豫良,劉 姝,張 露,吳彬彬
(1.淮海工學院海洋學院,江蘇連云港 222005;2.江蘇省海洋資源開發(fā)研究院,江蘇連云港 222001)
交替假單胞菌LP621菌株產(chǎn)右旋糖苷酶的培養(yǎng)條件優(yōu)化
呂明生1,2,王淑軍1,2*,房耀維1,2,焦豫良1,2,劉 姝1,2,張 露1,吳彬彬1
(1.淮海工學院海洋學院,江蘇連云港 222005;2.江蘇省海洋資源開發(fā)研究院,江蘇連云港 222001)
從江蘇連云港海域分離和篩選到1株產(chǎn)右旋糖苷酶的海洋細菌交替假單胞菌Pseudoalterom onas tetraodonisLP621,通過單因素試驗和正交試驗對該菌株產(chǎn)右旋糖苷酶培養(yǎng)條件進行優(yōu)化。單因素試驗結(jié)果表明,最佳培養(yǎng)時間為24 h,最適產(chǎn)酶溫度為25℃;產(chǎn)酶pH范圍為5.0~11.0,最適產(chǎn)酶pH為6.0;產(chǎn)酶NaCl濃度范圍為1%~10%,NaCl濃度為4%時產(chǎn)酶較高;裝液量在25%。麥芽糖、胰蛋白胨和酵母膏促進產(chǎn)酶。利用響應(yīng)面方法對LP621產(chǎn)右旋糖苷酶的發(fā)酵條件進行優(yōu)化。選擇培養(yǎng)基pH、時間、麥芽糖濃度和裝液量4因素進行優(yōu)化,結(jié)果為pH 7.07,發(fā)酵時間21.94 h,麥芽糖濃度0.42%,裝液量為21.88%,酶活為270.1 U/mL。
右旋糖苷酶;交替假單胞菌屬;優(yōu)化
右旋糖苷酶(Dextranase,α-D-1,6-Glucan-6-DGlucanohydrolase,EC3.2.1.11),又叫α-葡聚糖酶,是專一切割右旋糖苷(Dextean)中α-1,6糖苷鍵的水解酶[1]。右旋糖苷酶在齲齒防治、制糖工業(yè)和藥用右旋糖苷的生產(chǎn)中具有重要作用[2]。丹麥Novo公司、美國Miles公司等都有商品右旋糖苷酶制品出售,在國內(nèi)此項工作開展甚少,也無該酶制劑的生產(chǎn)。目前國內(nèi)外報道的產(chǎn)右旋糖苷酶微生物主要為青霉(Penicillium)[3]、斯達油脂酵母(L ipom yces starkeyi)[4]、對角毛殼菌(Chaetom ium gracile)[5]、曲霉(Aspergillus ustus)[6]、芽胞桿菌(Bacillus)[1]、鏈球菌(Streptococcus)[7-8]等。據(jù)目前報道的右旋糖苷酶中,酶的最適作用溫度多在50℃左右[1,3,6-7],對熱較敏感,在50℃保溫30 min酶活力損55%[6]。海洋中蘊藏著巨大的微生物資源,海洋微生物長期生活在海水環(huán)境中,所產(chǎn)生的酶類比那些來自中溫物種的酶類在低溫條件下具有更高的催化效力。已發(fā)現(xiàn)的海洋微生物酶包括有溶菌酶、唾液酸酶、氫化酶、谷氨酰胺酶、葡萄糖脫氫酶、甲基化酶、脂肪酶、DNA聚合酶、木聚糖酶、環(huán)糊精酶、纖維素酶、甘露聚糖酶、果膠裂解酶、氨單價氧化酶等。本課題組已從海洋環(huán)境中篩選了10株具有產(chǎn)生右旋糖苷酶的海洋細菌,對其中酶活較高的菌株621進行了形態(tài)特征、分子鑒定以及酶性質(zhì)研究,初步鑒定菌株LP621為交替假單胞菌Pseudoalterom onas tetraodonis。該菌生長溫度范圍為4~37℃,最適生長溫度為25℃;在pH 6.0~11.0范圍內(nèi)生長良好,最適生長pH為10.0;在0.5%~12%的NaCl濃度范圍內(nèi)可以生長,最適生長的NaCl濃度為7%,無NaCl不能生長,是1株低溫嗜鹽嗜堿菌(另文報道)。該菌產(chǎn)生的右旋糖苷酶作用溫度低(30℃),耐熱性較好,具有較好的應(yīng)用前景。目前國內(nèi)外尚無交替假單胞菌產(chǎn)右旋糖苷酶的報道。本文主要對該菌產(chǎn)低溫右旋糖苷酶的培養(yǎng)條件進行優(yōu)化研究,為低溫右旋糖苷酶的進一步研究和開發(fā)奠定基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 菌株來源 1株分離自江蘇連云港海域的交替假單胞菌Pseudoalterom onas tetraodonis LP621(在GenBank的登錄號為EU84912),由淮海工學院海洋學院實驗室分離和保藏。
1.1.2 培養(yǎng)基 種子培養(yǎng)基:蛋白胨0.5%,酵母粉0.1%,陳海水配制,pH 8.0;產(chǎn)酶培養(yǎng)基:酵母膏0.5%,蛋白胨0.5%,乳糖0.5%,右旋糖苷1%,陳海水配制,pH 8.0;陳海水配置人工海水(g/L):VOSO4·2H2O 0.005,LiCl 0.05,H3BO30.1,NiCl2·6H2O 0.01,BaCl2·2H2O 0.005, CuSO4·5H2O 0.01,ZnSO4·7H2O 0.1,CoCl2· 6H2O 0.005,MnCl2·4H2O 0.2,Na2MoO4·2H2O 0.1,KBr 0.05,KI 0.05,NaF 0.05,Al2(SO4)30.05,H2WO40.005,SrCl2·6H2O 0.005。
1.2 方法
1.2.1 產(chǎn)酶發(fā)酵方法 接種交替假單胞菌(P. tetraodonisLP621,以下簡稱LP621)斜面活化后的菌種于種子培養(yǎng)基,搖床振蕩培養(yǎng),25℃、180 r/ min,培養(yǎng)16 h,得種子液。將種子液以2%的接種量接種于產(chǎn)酶培養(yǎng)基(300 mL三角瓶裝產(chǎn)酶培養(yǎng)基75 mL),180 r/min,25℃振蕩培養(yǎng)24 h。培養(yǎng)結(jié)束后取出培養(yǎng)液,以10000 r/min離心5 min,取沉淀。加3 mL緩沖液溶解沉淀,裝入10 mL離心管中,冰水浴中超聲波破碎5 min。12000 r/min離心10 min,測定上清液的酶活力。
1.2.2 單因素產(chǎn)酶條件的研究 ①培養(yǎng)時間對產(chǎn)酶的影響:將LP621菌株接種到產(chǎn)酶培養(yǎng)基中,培養(yǎng)48 h,其他同1.2.1,每隔4 h取出,測定OD600和酶活力;②溫度對產(chǎn)酶影響:分別在不同溫度(15、20、25、30℃)下進行產(chǎn)酶發(fā)酵實驗,其他同1.2.1;③培養(yǎng)基初始pH值對產(chǎn)酶的影響:按照實驗標準配方配制1 L基礎(chǔ)培養(yǎng)基,配制培養(yǎng)基時需添加緩沖液(醋酸∶硼酸∶磷酸=1∶1∶1),添加濃度為10 mmol/L。加NaOH或HCl分別調(diào)pH至5.0~11.0,其他同1.2.1;④NaCl濃度對產(chǎn)酶的影響:用人工海水替代陳海水配置不同NaCl濃度(1%~10%)的產(chǎn)酶培養(yǎng)基,其他同1.2.1;⑤裝液量對產(chǎn)酶的影響:搖瓶裝液量分別為10%、15%、20%、25%、30%、40%進行產(chǎn)酶試驗,其他同1.2.1;⑥碳、氮源對產(chǎn)酶的影響:去除產(chǎn)酶培養(yǎng)基中的乳糖,分別在培養(yǎng)基加入終濃度為0.5%的各種碳源(分別為可溶性淀粉、葡萄糖、麥芽糖、乳糖、右旋糖苷、蔗糖、纖維素)進行碳源對產(chǎn)酶影響的發(fā)酵試驗。去除發(fā)酵培養(yǎng)基中的酵母膏和蛋白胨,分別加入終濃度為0.5%的各類氮源(分別為蛋白胨、酵母膏、硫酸氨、胰蛋白胨、酪蛋白、玉米粉)進行氮源對產(chǎn)酶影響的發(fā)酵試驗,測酶活力。
1.2.3 響應(yīng)曲面法(RS M)優(yōu)化發(fā)酵條件 在上述各單因素實驗的基礎(chǔ)上,主要選取了pH、發(fā)酵時間、麥芽糖濃度、裝液量4個因素作為自變量,以所產(chǎn)的低溫右旋糖苷酶的相對酶活為響應(yīng)值,對海洋交替假單胞菌產(chǎn)右旋糖苷酶的搖瓶培養(yǎng)條件進行優(yōu)化,利用DX6Trial軟件設(shè)計實驗方案。
1.2.4 酶活力測定 將100μL酶液加入到100 μL 1%的右旋糖苷緩沖液中,在30℃水浴反應(yīng)15 min,用3,5-二硝基水楊酸(DNS)法測定還原糖量。酶活力單位定義:在上述反應(yīng)條件下,每分鐘催化產(chǎn)1μg麥芽糖的酶量為1個活力單位。
2 結(jié) 果
2.1 單因素培養(yǎng)條件的研究
2.1.1 培養(yǎng)時間對產(chǎn)酶的影響 將LP621菌株接入到培養(yǎng)基中,在25℃下靜置培養(yǎng)48 h,每4 h取出測酶活力及LP621菌株生長量(即OD600)。由圖1可知,右旋糖苷酶在24 h之前,產(chǎn)酶量上升速度較快,到發(fā)酵24 h時(即菌體生長到穩(wěn)定期)產(chǎn)酶量最高,隨時間增加,酶活力逐漸下降。
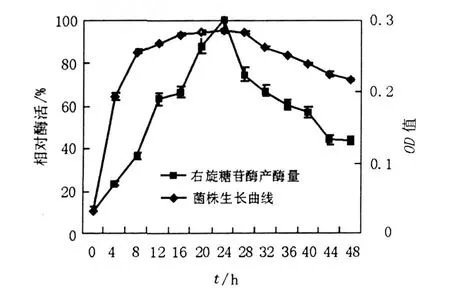
圖1 培養(yǎng)時間對菌株生長和產(chǎn)酶的影響Fig.1 The effect of time on growth of LP621 and production of dextranase
2.1.2 溫度對產(chǎn)酶的影響 LP621菌株能在4~37℃下生長,最適生長溫度為25℃,屬于低溫菌(另文報道),因此重點研究了15~30℃溫度下對菌株產(chǎn)酶的影響(圖2),20~25℃時右旋糖苷酶產(chǎn)量達到最高,低于20℃、高于25℃的情況下產(chǎn)酶均迅速下降。15℃和30℃時分別只有最高產(chǎn)酶量60%和73.1%的產(chǎn)酶量。
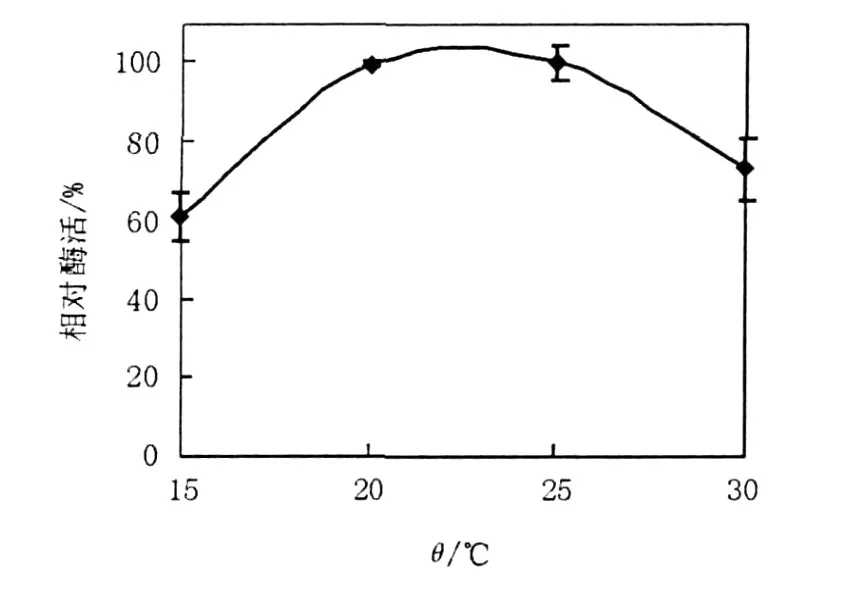
圖2 溫度對右旋糖苷酶產(chǎn)生的影響Fig.2 The effect of temperature on production of dextranase
2.1.3 pH對產(chǎn)酶的影響 LP621菌株生長pH范圍為6.0~11.0,最適生長pH為10.0,屬于嗜堿菌(另文報道),因此重點研究了pH 5.0~11.0對菌株產(chǎn)酶的影響(圖3),隨著pH的升高,酶產(chǎn)量逐漸增加,當pH達到6.0時產(chǎn)酶量達到最大; pH繼續(xù)升高,在pH 7.0~9.0之間酶產(chǎn)量下降較小,趨勢平穩(wěn),酶活相當穩(wěn)定;自pH值為9后,酶產(chǎn)量迅速下降。在低于pH 5.0或高于pH 10.0時酶產(chǎn)量均很低。
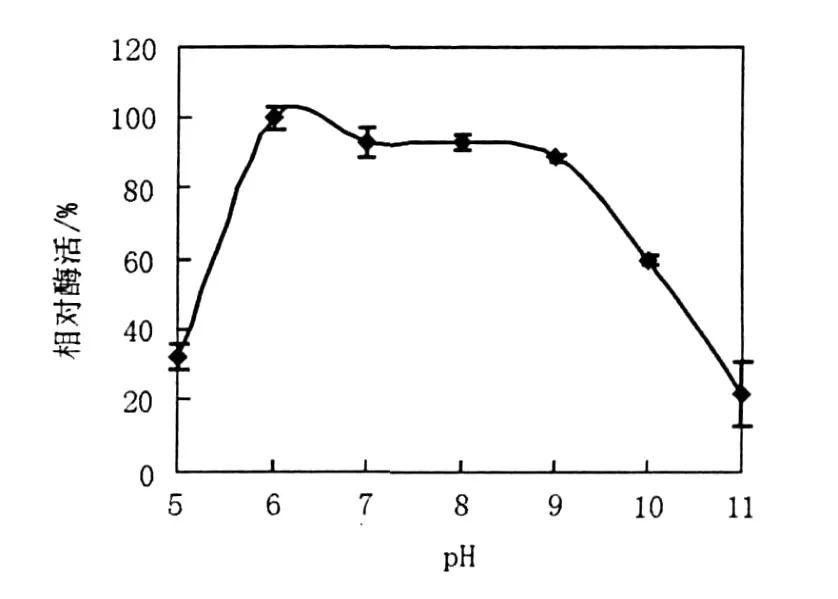
圖3 pH對右旋糖苷酶產(chǎn)生的影響Fig.3 The effect of pH on production of dextranase
2.1.4 不同NaCl濃度對產(chǎn)酶的影響 如圖4所示,產(chǎn)右旋糖苷酶的量隨NaCl濃度的增加而逐漸增加,當培養(yǎng)基NaCl濃度升至4%時,產(chǎn)右旋糖苷酶最高,進一步升高NaCl濃度則會對LP621菌體生長和右旋糖苷酶酶活積累產(chǎn)生一定的抑制作用。

圖4 NaCl濃度對右旋糖苷酶產(chǎn)生的影響Fig.4 The effect of NaCl concentration on production of dextranase
2.1.5 裝液量對產(chǎn)酶的影響 通過控制裝液量來控制發(fā)酵液的溶氧情況,由圖5可知,裝液量太少溶氧過大,對LP621菌體造成傷害不利于產(chǎn)酶,裝液量在25%時產(chǎn)酶適宜。
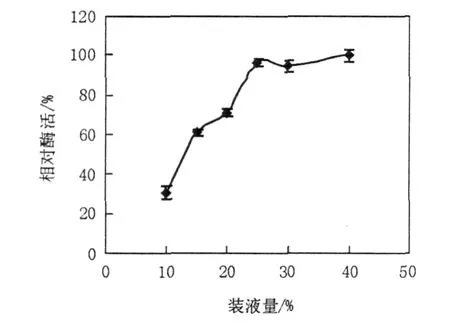
圖5 裝液量對右旋糖苷酶產(chǎn)生的影響Fig.5 The Effect of medium amount on production of dextranase
2.1.6 不同碳氮源對產(chǎn)酶的影響 通過在培養(yǎng)基中添加不同的碳源和氮源進行發(fā)酵培養(yǎng),結(jié)果見表1。麥芽糖最能促進LP621產(chǎn)酶,乳糖和可溶性淀粉次之,葡萄糖、纖維素不利于產(chǎn)酶,培養(yǎng)基中添加不同碳源對產(chǎn)酶的影響較大。胰蛋白胨促進產(chǎn)酶,酵母膏和酪蛋白、蛋白胨、玉米粉次之,無機氮源不利于產(chǎn)酶。
2.2 響應(yīng)曲面法(RSM)優(yōu)化發(fā)酵條件
2.2.1 響應(yīng)面設(shè)計實驗及其結(jié)果 在以上單因素發(fā)酵實驗的基礎(chǔ)上,選取了pH、時間、麥芽糖和裝液量4個因素為自變量,以所產(chǎn)的低溫右旋糖苷酶的相對酶活為響應(yīng)值,采用響應(yīng)面軟件設(shè)計實驗方案,分別發(fā)酵測定酶活力,結(jié)果見表2。從表2可以看出,在培養(yǎng)基pH為7.0、時間為24 h、麥芽糖濃度為1%、裝液量為25%時,測得的酶活最高。但4個因素具體的值還要通過響應(yīng)面軟件進一步分析。
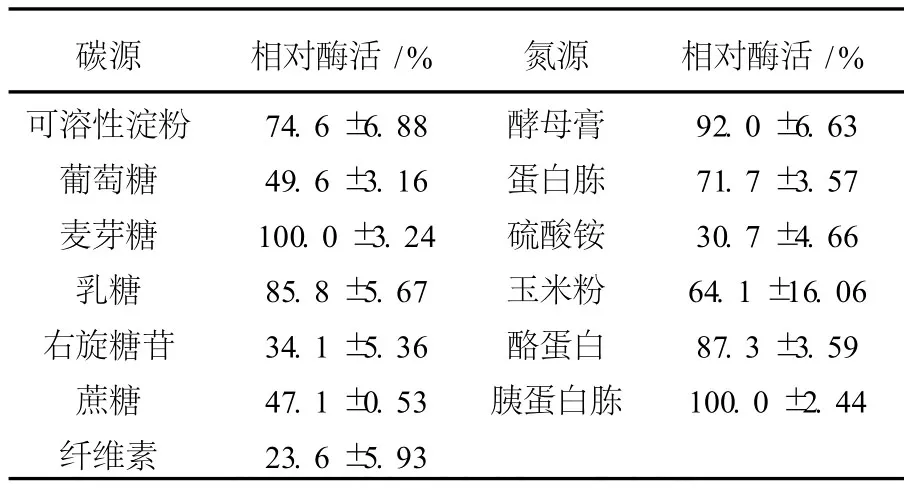
表1 碳氮源對右旋糖酐酶產(chǎn)生的影響Table1 Effect of carbon and nitrogen source on production of dextranase
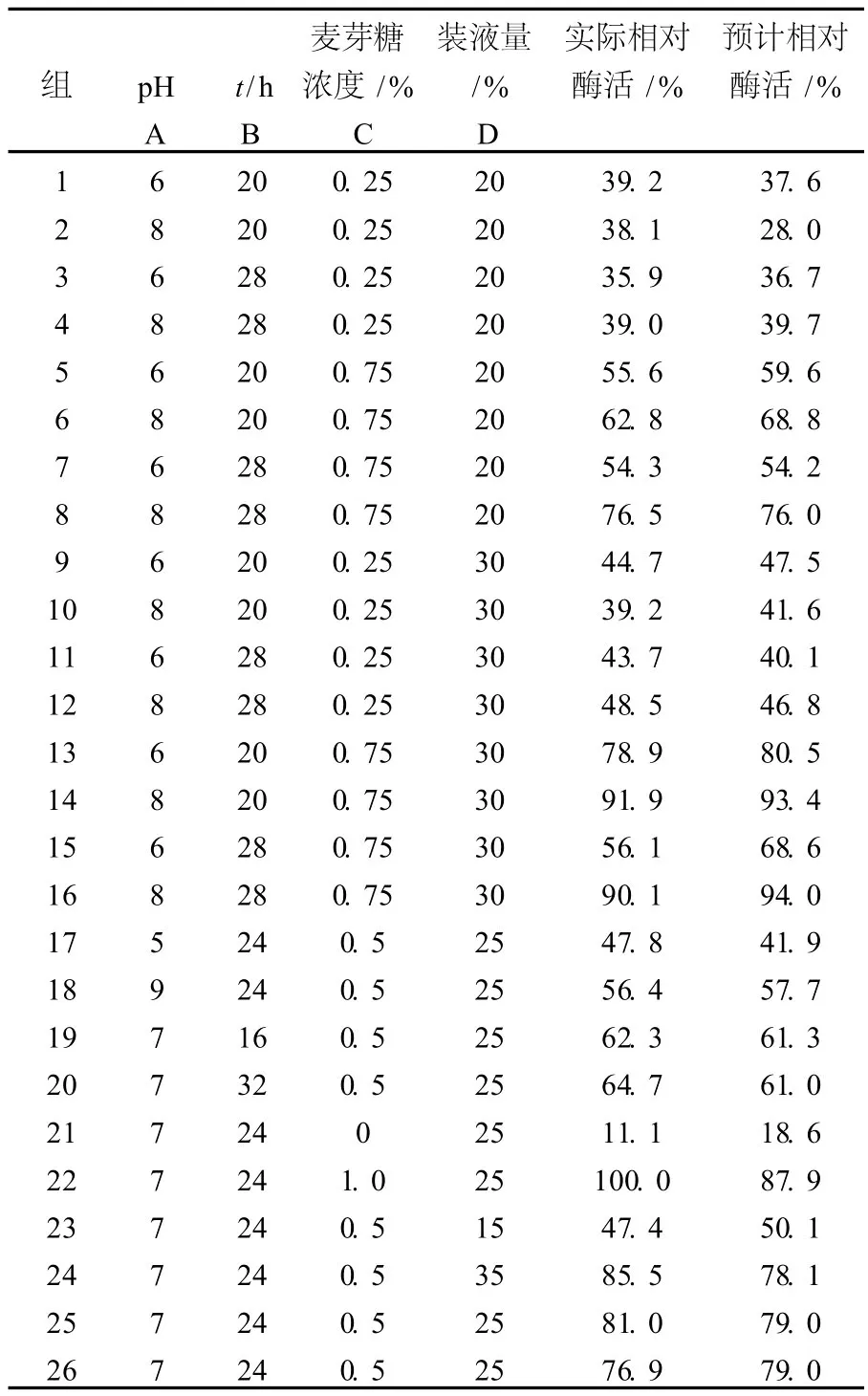
表2 響應(yīng)面實驗設(shè)計及其實驗結(jié)果Table 2 Experimental design and results response analysis
2.2.2 二次回歸擬合以及方差分析 以相對酶活為響應(yīng)值,經(jīng)回歸擬合后各試驗因子對響應(yīng)值的影響可以用以下函數(shù)表示:Y=a0+a1A +a2B+a3C+a4D+a5A2+a6B2+a7C2+ a8D2+a9AB+a10AC+a11AD+a12BC+ a13BD+a14CD。應(yīng)用響應(yīng)面軟件對26個響應(yīng)值進行分析所得到的擬合全變量二次回歸方程各變量的偏回歸系數(shù)估計值及方差分析見表3。由表3可知,決定系數(shù)R2=0.9013,說明回歸方程的擬合程度較好,因此可以用該回歸方程代替試驗真實點對試驗結(jié)果進行分析。由響應(yīng)面軟件可以得出全變量二次回歸方程:Y= 79.30+3.95A-0.063B+17.32C+7.00D-7.38A2-4.53B2-6.52C2-3.79D2+3.16AB +4.69AC+0.93AD-1.13BC-1.64BD+ 2.74CD。
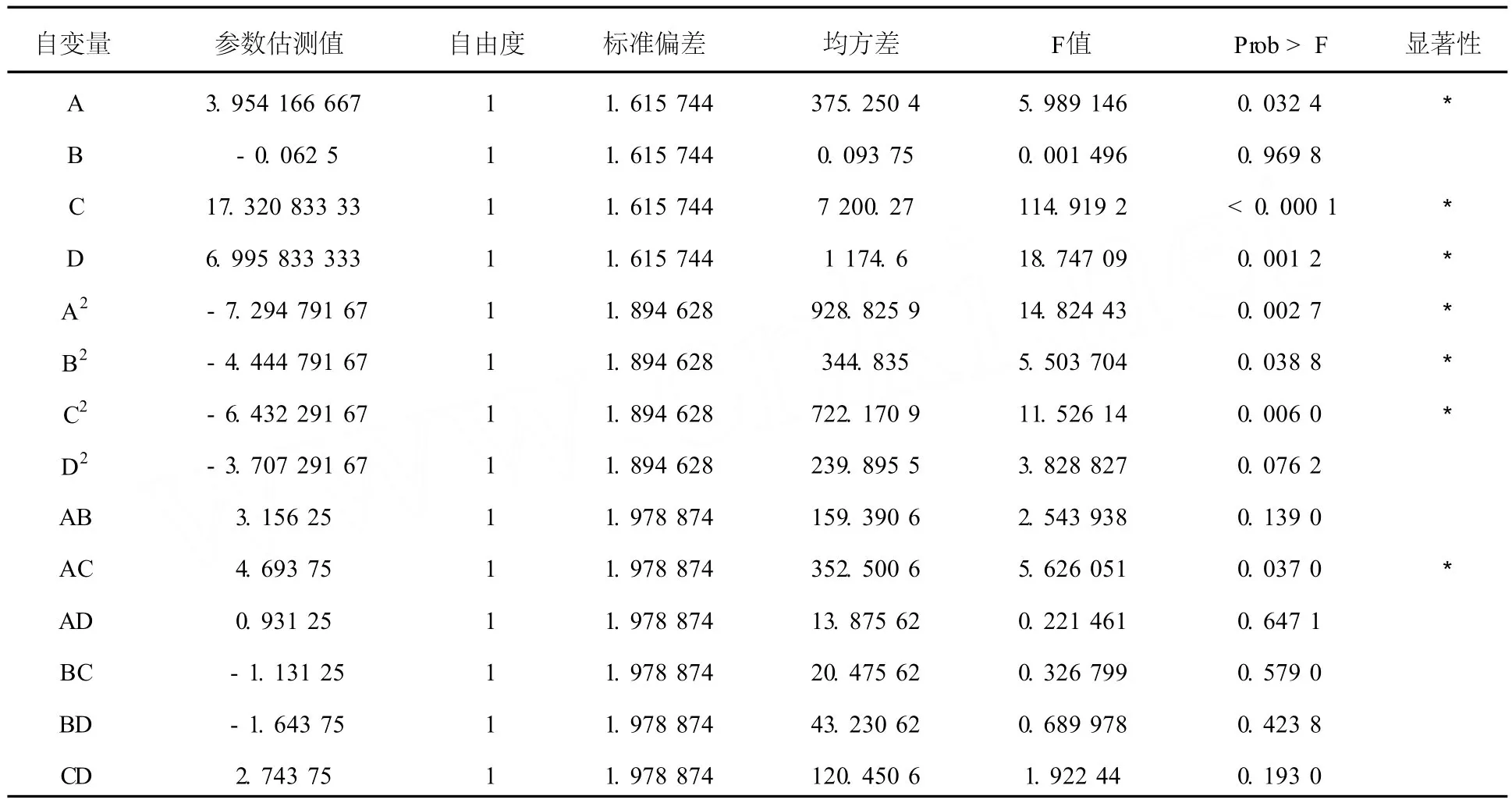
表3 回歸分析結(jié)果Table 3 Results of the regression analysis
2.2.3 發(fā)酵條件的最適組合 在獲得非線性回歸模型和響應(yīng)面之后,為了求得最佳發(fā)酵條件,對所得的回歸擬和方程分別對各自的變量求一階偏導數(shù),求解此方程組可以得到最大酶活時的最佳條件:pH 7.07,發(fā)酵時間21.94 h,麥芽糖濃度0.42%,裝液量為21.88%,在此條件下理論預(yù)測最大相對酶活為99.7%,酶活為270.1 U/mL。
2.2.4 各因素交互作用的響應(yīng)面圖和等高線圖
和單因素試驗相比響應(yīng)曲面優(yōu)化培養(yǎng)條件的優(yōu)勢在于響應(yīng)面優(yōu)化法可以考察因素間的交互作用。等高線的形狀可反映出交互效應(yīng)的強弱大小,橢圓形表示兩因素交互作用顯著,而圓形則與之相反。根據(jù)響應(yīng)面軟件繪出的等高線和響應(yīng)面立體分析圖(圖6~圖11),可以得出培養(yǎng)基pH、麥芽糖濃度和裝液量對產(chǎn)酶的影響較大,之間的交互作用都很顯著,這與方差分析中顯著性的分析結(jié)果相符。
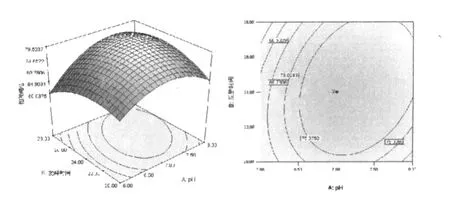
圖6 發(fā)酵時間和pH對產(chǎn)右旋糖苷酶影響的響應(yīng)面立體分析圖和等高線Fig.6 Response surface plot and contour plot of dextranase production showing interaction between time value and pH value at the central value of the other parameter
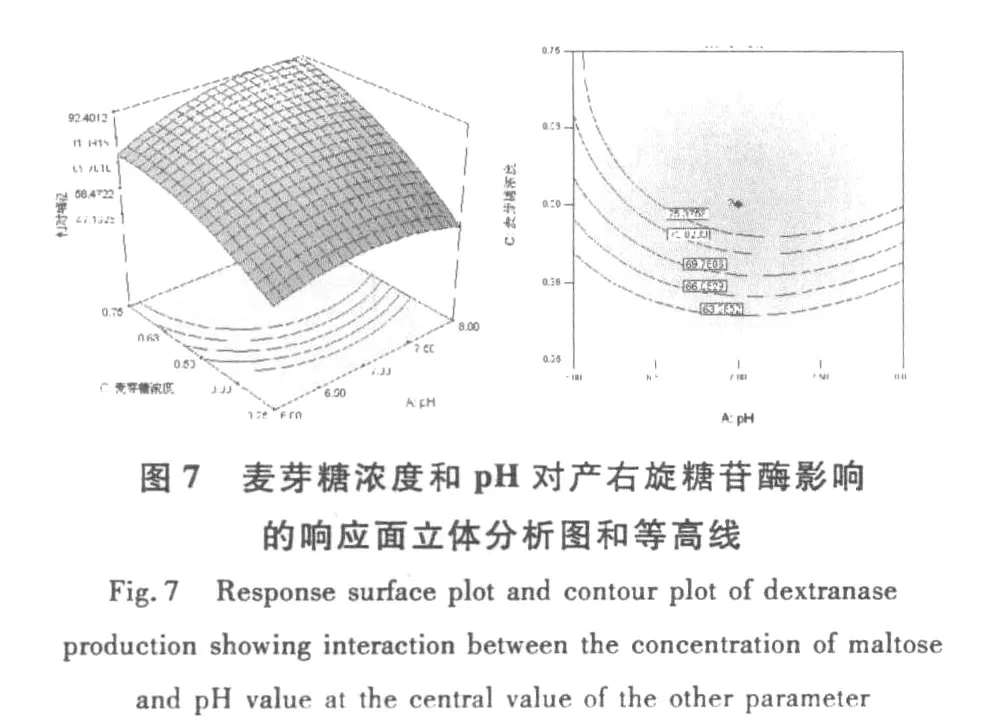
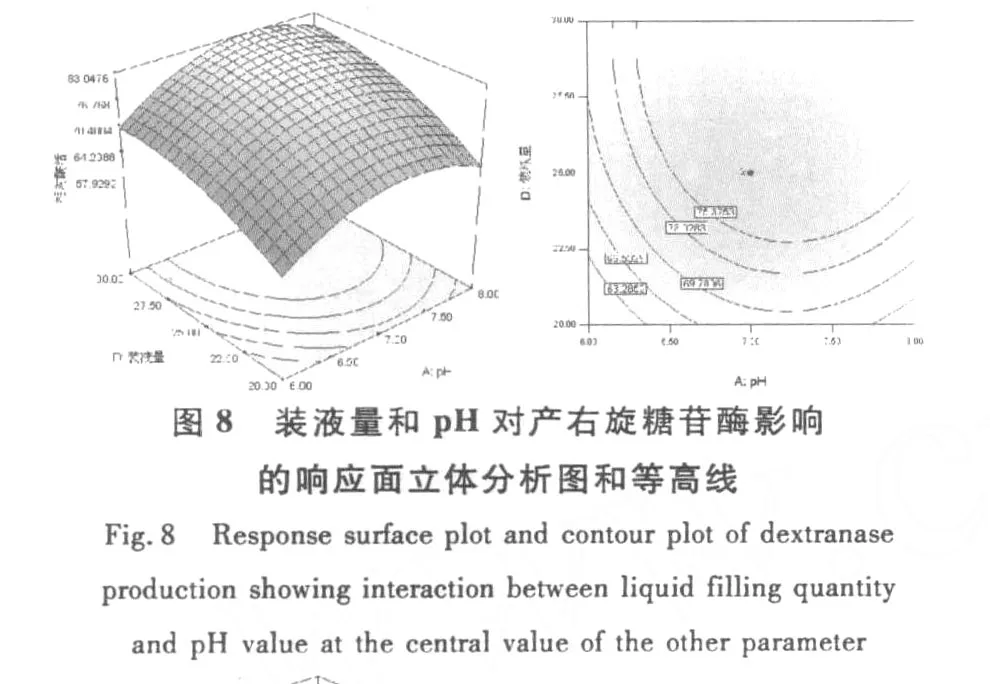

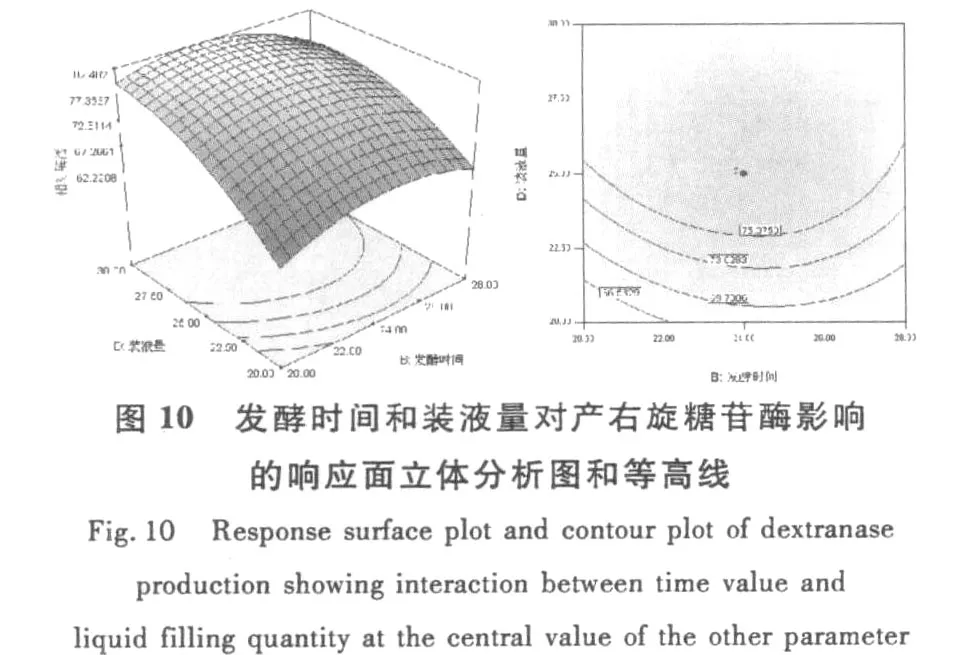
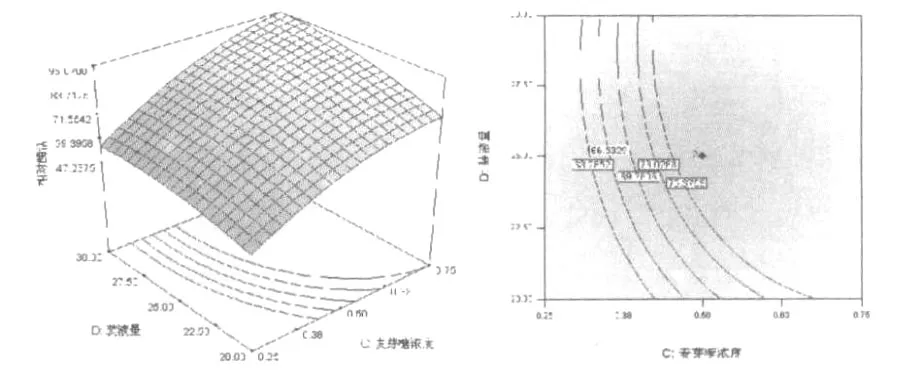
圖11 麥芽糖濃度和和裝液量對產(chǎn)右旋糖苷酶影響的響應(yīng)面立體分析圖和等高線Fig.11 Response surface plot and contour plot of dextranase production showing interaction between he concentration ofmaltose and liquid filling quantity at the central value of the other parameter
2.2.5 驗證實驗 在優(yōu)化后的培養(yǎng)基及培養(yǎng)條件下進行5組平行試驗,所得酶活分別為270.1、270.1、270.1、270.1、270.1 U/mL,平均值為270.1 U/mL。回歸方程所得到的納他霉素產(chǎn)量的預(yù)測值(2.19 g/L)與驗證試驗的平均值(2.14 g/L)相接近,說明回歸方程能夠較真實地反映各篩選因素對發(fā)酵產(chǎn)酶的影響。
3 討 論
交替假單胞菌屬(Pseudoalterom onas)是Gauthier G等[9]在1995年提出的一個新建立的海洋細菌屬。目前在世界范圍海洋中,包括南極洲嚴寒海洋環(huán)境中分離到多株交替假單胞菌,已經(jīng)鑒定到種的有P.tunicata、P.agarolytic和P.bacteriolytica等30多種,每年都有新種發(fā)現(xiàn)[10-13]。其獨特的生物學特性引起了越來越多研究者的關(guān)注,該屬菌能夠產(chǎn)生多種活性物質(zhì),表現(xiàn)出多種生物活性。交替假單胞菌能分泌多種胞外活性物質(zhì),包括胞外酶、胞外毒素、抗生素和胞外多糖等,這些物質(zhì)表現(xiàn)出抗菌、溶菌、殺菌、分解半乳糖、降解纖維素和果膠以及軟化瓊膠等多種生物活性,有助于交替假單胞菌獲取營養(yǎng)和競爭生存空間等,具有很重要的生態(tài)學作用[14]。目前交替假單胞菌產(chǎn)生的低溫酶主要有脂肪酶、瓊脂糖酶、α-半乳糖酶、DNA連接酶、蛋白酶、纖維素酶和淀粉酶等[15-20]。
菌株LP621為交替假單胞菌,是本課題組從海洋環(huán)境中篩選的1株交替假單胞菌屬(Pseudoalterom onas),并從培養(yǎng)液中檢測到了右旋糖苷酶的活性,但活性不高,為了獲得大量的酶,以便于酶性質(zhì)的研究和酶基因的克隆表達,采用單因素和響應(yīng)面方法對菌株產(chǎn)酶條件進行了優(yōu)化研究。目前國內(nèi)外沒有交替假單胞菌屬產(chǎn)右旋糖苷酶的研究報道。
菌株LP621是需氧的革蘭陰性桿菌,最適生長溫度為25℃,生長溫度范圍為4~37℃;最適生長pH為10.0,在pH 6.0~11.0范圍內(nèi)生長良好;最適生長的NaCl濃度為7%,在0.5%~12%的NaCl濃度范圍內(nèi)可以生長,無NaCl不能生長,是1株低溫嗜鹽嗜堿菌。該菌株生長4 h即進入對數(shù)期,培養(yǎng)24 h菌株為平衡期時,菌株產(chǎn)酶到達高峰。菌株低于20℃時生長緩慢,此時產(chǎn)酶量也相應(yīng)降低;而25℃時,菌體生長最快,此時也是產(chǎn)酶的最佳溫度,這與文獻[21]中報告的產(chǎn)酶最佳溫度與菌株生長溫度一致。LP621菌株生長和產(chǎn)酶的pH和NaCl濃度范圍與文獻中報告基本一致,但最適生長和產(chǎn)酶的pH和NaCl濃度不一致。最適生長pH為10.0,最適產(chǎn)酶pH為6.0,而到pH為10.0酶活下降;最適生長的NaCl濃度為7%,但NaCl濃度為4%時產(chǎn)酶較高。
[1]Khalikova E,Susi P,Usanov N,et al.Purification and properties of extracellular dextranase from a Bacillus sp.[J].J.Chromatogr B,2003,796(2):315-326.
[2]Khalikova E,Susi P,Korpela T.Microbial dextran-hydrolyzing enzymes:fundamentals and applications[J].Microbiol Mol Biol Rev,2005,69(2):306-325.
[3]Aslan Y,Tanriseven A.Immobilization of Penicillium lilacinum dextranase to produce isomaltooligo saccharides from dextran [J].Biochem Eng,J,2007,34(1):8-12.
[4]Chen L,Zhou X,Fan W,Zhang Y,et al.Expression,purification and characterization of a recombinant Lipomyces starkey dextranase in Pichia pastoris[J].Protein Expr Purif,2008,58 (1):87-93.
[5]Hattori A,Ishibashi K,Minato S.The purification and characterization of the dextranse of Chaetom ium gracile[J].Agri Bio Chem,1981,45(11):2409-2416.
[6]程秀蘭,孫晉武,王海燕,等.焦曲霉右旋糖苷酶的純化和性質(zhì)[J].微生物學報,1992,32(3):218-226.
[7]蔣丹,仇元新,胡濤,等.口腔鏈球菌右旋糖苷酶分子結(jié)構(gòu)和功能的研究進展[J].國際口腔醫(yī)學雜志,2008,35(3):249-251.
[8]Kim YM,Seo M Y,Kang H K,et al.Construction of a fusion enzyme of dextransucrase and dextranase:Application for one step synthesis of isomalto-oligosaccharides[J].Enzy Micro Tech,2009,44(3):159-164.
[9]Gauthier G,Gauthier M,Christen R.Phylogenetic analysis of the genera Alteromonas,Shewanella,and Morltella using genes coding for small-subunit rRNA sequences and division of the genus Alteromonas into two genera,Alteromonas(emended)and Pseudoalteromonas gen.nov,and proposal of twele new species combinations[J].Int J Syst Bacteriol,1995,45:755-761.
[10]Carolina CR,Rowley AF.Effect of extracellular products of Pseudoalterom onas atlantica on the edible crab Cancer pagurus [J].Appl Environ Microbiol,2004,70(2):729-735.
[11]Park YD,Baik KS,Yi H,et al.Pseudoalterom anas byunsanensis sp.nov.,isolated from tidal flat sediment in Korea[J].Int J Syst Evol Microbiol,2005,55:2519-2523.
[12]Al Khudary R,St?sser N I,Qoura F,et al.Pseudoalterom onas arcticasp.nov.,an aerobic,psychrotolerant,marine bacterium isolated from Spitzbergen[J].Int J Syst Evol Microbilo,2008, 58(9):2018-2024.
[13]Yu Y,Li H,Zeng Y,et al.Extracellular enzymes of cold-adapted bacteria from Arctic sea ice,Canada Basin[J].Polar Biology, 2009,32(10):1539-1547.
[14]席宇,朱大恒,劉紅濤,等.假交替單胞菌及其胞外生物活性物質(zhì)研究進展[J].微生物學通報,2005,32(3):108-112.
[15]Vera J,Aluarez R,Murano E,et al.Identification of a Marine Agarolytic Pseudoalterom onas Isolate and Characterization of Its Extracellular Agarase[J].Appl Emirom Microbiol,1998,64 (11):4378-4383.
[16]Bakunina IY,Sova VV,Nedashkovskaya O I,et al.Alpha-galactosidase of the marine bacterium Pseudoalterom onas sp. K MM701[J].Biochemisty(Masc),1998,63(10):1209-1215.
[17]Ceorlette D,Jonsson ZO,Van Petegem F,et al.A DNA ligase from the psychrophile Pesudoalterom onas haloplankitis gives insights into the adaptation of proteins to low temperatures[J]. Eur J Biochem,2000,267(12):3502-3512.
[18]李丹,陳麗,李富超,等.一株海洋細菌Pseudoalteromonas flavipulchraHH407產(chǎn)低溫堿性蛋白酶的篩選與生長特性的研究[J].食品與生物技術(shù)學報,2007,26(6):74-80.
[19]呂明生,呂鳳霞,房耀維,等.低溫纖維素酶產(chǎn)生菌的篩選、鑒定及酶學性質(zhì)初步研究[J].食品科學,2007,28(12): 235-239.
[20]劉紅飛,呂明生,王淑軍,等.產(chǎn)低溫淀粉酶海洋細菌Pseudoalteromonas sp.GS230發(fā)酵條件與酶學性質(zhì)的研究[J].中國釀造,2008,13:13-18.
[21]程秀蘭,孫晉武,楊敬,等.淡紫擬青霉右旋糖苷酶的形成條件[J].微生物學報,1992,32(5):334-339.
Culture Optimization of Dextranase-Producing Pseudoalterom onas tetraodonis LP621
LV Ming-sheng1,2,WANG Shu-jun1,2,FANG Yao-wei1,2, JIAO Yu-liang1,2,LIU Shu1,2,ZHANGLu1,WU Bin-bin1
(1.Coll.of Marine Sci.and Technol.,Huaihai Inst.of Technol., Lianyungang,222005; 2.Jiangsu Marine Res.Devel.Res.Inst.,Lianyungang, 222001,China)
A dextranse-producing marine bacteria Pseudoalterom onas tetraodonisLP621 was screened and isolated from the sea area of Lianyungang,Jiangsu.Its dextranase production conditions was optimized through single-factor and orthogonal experiments.The results of single factor experiment indicated that the most suitable culture condition was 24 hours,and the most suitable dextranse producing temperature was 25℃at pH 6.0 with ranging 5.0~11.0, and the range of the concentration of NaCl for the dextranse production was 1%~10%,and the yield of dextranase was fairly high at 4%of NaCl with filling volume rate at 25%.Maltose,tryptone,and yeast extract enhanced the enzyme production.Response surface method was adopted to optimize the enzyme production fermentation conditions. Medium pH,time,maltose,and filling volume four factors were selected for the optimization.And the results were pH 7.07,21.94 h,maltose 0.42%,and filling volume 21.88%,the enzyme activity was 270.1 U/mL.
dextranase;Pseudoalterom onas tetradonis;optimization
Q939;TS275.4
A
1005-7021(2010)06-0011-07
國家自然科學基金(40746030);江蘇省科技廳工業(yè)支撐計劃項目(BE2009095);江蘇省海洋生物技術(shù)重點建設(shè)實驗室開放課題項目(2008HS008);連云港市自然科學基金(ZH200801)
呂明生 男,副教授。研究方向為海洋生物技術(shù)。Tel:0518-85895421,E-mail:mingshenglu@hotmail.com
*通訊作者。E-mail:shujunwang86@163.com
2010-11-01;
2010-11-21
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
