重組谷氨酸棒桿菌產蘇氨酸發酵培養基的優化*
2010-11-02 06:26:19呂揚勇伍展紅鄭穗平
食品與發酵工業 2010年12期
呂揚勇,伍展紅,鄭穗平
(華南理工大學生物科學與工程學院,廣東廣州,510006)
重組谷氨酸棒桿菌產蘇氨酸發酵培養基的優化*
呂揚勇,伍展紅,鄭穗平
(華南理工大學生物科學與工程學院,廣東廣州,510006)
通過基因敲除構建了蛋氨酸合成途徑關鍵酶基因metX缺失的重組谷氨酸棒桿菌R102ΔmetX。采用部分因子實驗設計考察發酵培養基中各組分對重組菌產蘇氨酸的影響。結果表明,葡萄糖、酵母粉和酶解酪素對產蘇氨酸影響顯著。繼而采用最陡爬坡路徑逼近最大響應區域,并結合中心組合實驗和響應面對3個顯著性因素進行分析,得到優化的發酵培養基組成:葡萄糖50.78g/L,酵母粉6g/L,酶解酪素0.99g/L,生物素80μg/L,VB12mg/L,CaCO320g/L,(NH4)2SO415.15g/L,MgSO4·7H2O 0.5g/L,KH2PO40.5g/L,K2HPO40.5g/L,Fe-SO4·7H2O 0.02g/L,MnSO4·7H2O 0.02g/L,接種量8%。采用該優化培養基,供試菌株的胞外蘇氨酸濃度為3.35g/L,較優化前提高了30%。
谷氨酸棒桿菌,L-蘇氨酸,響應面,培養基優化
L-蘇氨酸是人和動物體內一種必需氨基酸[1],同時也是一種重要的營養強化劑,已廣泛應用于飼料工業、保健食品和醫藥工業。自1935年 Rose和Meger在纖維蛋白水解物中分離并鑒定出蘇氨酸以來[2],已發現的能產生蘇氨酸的微生物主要有以下幾種:大腸桿菌(Escherichiacoli)、谷氨酸棒桿菌(Corynebacterium glutamicum)、粘質沙雷氏菌(Serratia marcescens)和黃色短桿菌(Brevibacterium flavum)。近年來隨著谷氨酸棒桿菌模式菌株ATCC 13032基因組測序的完成[3],利用基因工程改造谷氨酸棒桿菌成為現實[4]。要提高蘇氨酸產量,為進一步的育種工作提供好的基礎,除利用基因工程改造以外,發酵培養基的優化也較為重要。目前還未見針對重組谷氨酸棒桿菌發酵培養基響應面優化生產蘇氨酸的相關報道。
基于非完全平衡塊原理的Plackett-Burman(PB)法能通過對每個因子取2水平進行分析,通過比較各個因子兩水平的差異與整體的差異來確定因子的顯著性[5]。響應面法(Response Surface Methodology,RSM)可以建立連續變量曲面模型,并可以同時對影響產量的各因子水平及其交互作用進行優化與評價。因此PB設計及響應面法已廣泛應用于各類培養基的優化研究中[6-7]并取得了良好的效果。本研究利用PB設計法,對重組谷氨酸棒桿菌產蘇氨酸培養基的主要影響因子進行篩選,然后采用響應面法對主要影響因子間的交互作用進行了研究,以獲得產蘇氨酸的最優培養基,提高重組谷氨酸棒桿菌的產酸能力,為食品工業蘇氨酸育種提供基礎。
1 材料與方法
1.1 材料
1.1.1 供試菌株
重組谷氨酸棒桿菌R102ΔmetX(Corynebacterium glutamicum R102(AHVr)with deletion in the metX gene),為本實驗室構建。
1.1.2 培養基
種子培養基:種子培養基(g/L):葡萄糖40,蛋白胨20,KH2PO41.5,K2HPO40.5,MgSO4·7H2O 0.5,酵母粉 5,生物素 50μg/L。
發酵培養基(g/L):葡萄糖100,KH2PO40.5,K2HPO40.5,(NH4)2SO420,MgSO4·7H2O 0.25,Fe-SO4·7H2O 0.01,MnSO4·4H2O 0.01,CaCO320,酵母粉5,生物素 400μg/L,VB12mg/L,上述培養基均需要調整pH值為7.0。
1.2 試驗方法
1.2.1 菌種活化
將保存在-80℃凍存管中的菌種接于LB固體培養基上,30℃培養24 h,劃線轉接3次得到活化的菌種。
1.2.2 培養方法
將活化的菌種挑取単菌落接種于種子培養基中,于30℃,200 r/min培養24 h,以10%接種量接于裝有25mL發酵液的250mL的三角瓶中30℃,215 r/min發酵72 h,HPLC法檢測胞外蘇氨酸濃度。
1.2.3 發酵液中蘇氨酸的含量測定
采用高效液相分析系統(HPLC)柱前衍生測定。衍生方法:取發酵液1mL,12 000 r/min離心5 min,取上清液100μL加入1.5mL離心管中,加入已經配制好的衍生緩沖溶液(0.5mol/L NaHCO3溶液,pH9.0)100μL,搖勻后再加入衍生試劑溶液(1%2,4-二硝基氟苯的乙腈溶液)150μL,搖勻,將離心管置于60℃水浴中暗處恒溫加熱60 min后取出,冷卻溶液至室溫后,加入定容緩沖液(0.01mol/L KH2PO4,pH 7.0) 稀釋至1.2mL,用0.22 μm 針筒式過濾器過濾后,用于色譜分析。采用Agilent 1100高效液相色譜儀,Agilent 20RBAX SB-C18(4.6×150 nm,5 μm)色譜柱,紫外檢測器,360 nm 檢測。流動相分別為A:乙酸鈉緩沖溶液(pH 6.5,含10mL/L N,N-二甲基甲酰胺),B:60%乙腈。流動相速度為1.0mL/min;柱溫22℃。梯度洗脫程序見表1。

表1 流動相梯度程序
測定完畢以后利用標準曲線計算發酵液中蘇氨酸的含量。
1.3 試驗設計
1.3.1 PB實驗
本試驗選取12個因子(變量)進行試驗,各參數所代表因子及其水平見表2。每個試驗均進行2次,對應的響應值取2次試驗結果的平均值,評價指標(響應值)為發酵液中蘇氨酸濃度。
1.3.2 最陡爬坡試驗
根據PB試驗結果得到的一階模型按其回歸系數的符號和大小,設計顯著因子的最陡爬坡路徑和步長,找出峰值,從而快速接近最大響應區域。
1.3.3 響應面分析
由最陡爬坡試驗逼近最大響應區域后,采用旋轉中心組合設計(central composite rotatable design,CCD)法對其進行進一步研究,以獲得最佳發酵培養基。用多項式回歸對試驗數據進行擬合,得到一個描述響應值(蘇氨酸濃度)與自變量(發酵條件)關系的一個二次多項式方程(以3因素為例):

表2 Plackett-Burman試驗設計的因子水平及其編碼

式中:X1、X2、X3為由部分因子實驗確定對響應值影響顯著的3種培養基組分的編碼值,Y為預測響應值,即蘇氨酸濃度(g/L),b0、b1、b2、b3、b11、b12、b13、b22、b23、b33為方程系數。上述每個試驗均進行 2次,對應的響應值取2次試驗結果的平均值。實際考察的變量及其試驗水平編碼見表3。
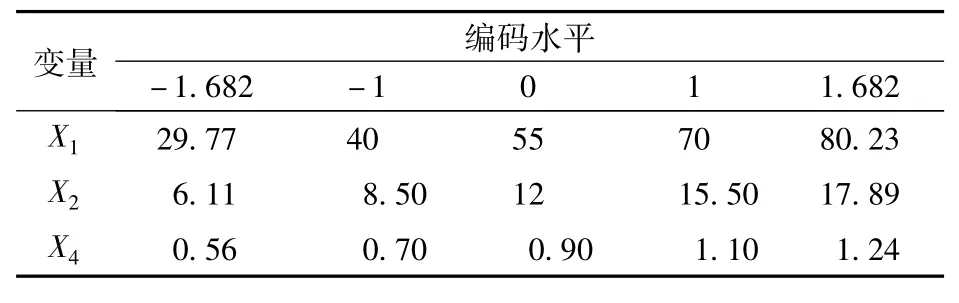
表3 旋轉中心組合設計試驗的因素水平及其編碼
2 結果與分析
2.1 PB篩選影響蘇氨酸產量的顯著因子
PB試驗結果見表4。通過minitab 15軟件對表4試驗數據進行方差分析,結果見表5,各因子顯著性分析見表6

表4 Plackett-Burman設計及試驗結果
采用minitab 15對表4中的蘇氨酸產量數據進行逐步回歸分析,剔除掉不顯著因子,得到以蘇氨酸產量為響應值的一階回歸方程:


表5 Plackett-Burman試驗回歸方程方差分析
回歸方程方差分析(表5)表明,所得方程達到極顯著(P=0.009),說明該回歸模型在被研究的整個回歸區域擬合的較好。相關系數R2=0.92,說明相關性較好,校正決定系數AdjR2=0.80,表明80%的試驗數據的變異性可用此回歸模型來解釋。

表6 Plackett-Burman試驗回歸方程系數的顯著性檢驗
由表6可以看到,影響R102ΔmetX發酵產蘇氨酸的重要因素為:X1、X2、X4,即葡萄糖、(NH4)2SO4和酶解酪素。3者對蘇氨酸產量的影響均為負效應,在后續最陡爬坡試驗中應該降低3者數值。不顯著因素中 MgSO4·7H2O、FeSO4·7H2O、MnSO4·7H2O為正效應,在后續試驗應取其高水平;其余均為負效應,在后續響應面試驗中應取其低水平。
2.2 最陡爬坡試驗
最陡爬坡試驗設計及結果見表7。由表7可知,X1、X2和X43個因子的濃度在試驗4附近時蘇氨酸濃度最高,故選擇試驗4中的各因子的濃度,即葡萄糖55g/L、(NH4)2SO412g/L、酶解酪素0.9g/L為后續旋轉組合試驗設計的中心點。

表7 爬坡實驗設計及結果
2.3 發酵培養基的響應面分析
2.3.1 二次回歸模型擬合及檢驗
以2.2中確定的葡萄糖、酵母粉和酶解酪素3個因子的濃度為中心點,以蘇氨酸濃度Y為響應值,設計旋轉中心組合試驗。根據試驗設計表進行了20組試驗,每組試驗重復2次,取其平均值為響應值,結果見表8。

表8 旋轉中心組合設計及其試驗結果
對表8試驗數據進行二次多項回歸擬合,得二次多項式回歸方程為:

式中:Y為蘇氨酸濃度預測值,X1、X2和X4分別為葡萄糖、(NH4)2SO4和酶解酪素的編碼值。對得到的模型(上述方程)進行方差分析和系數顯著性檢驗,方差分析結果表見表9,顯著性檢驗結果見表10。
由表9可看出,該模型極顯著(P <0.000 1),失擬項不顯著(P=0.212 1>0.1)。經分析計算,該模型的確定系數R2=0.969 1,表明模型與實際情況符合很好,該模型可用于蘇氨酸濃度的預測。

表9 蘇氨酸濃度預測回歸模型的方差分析
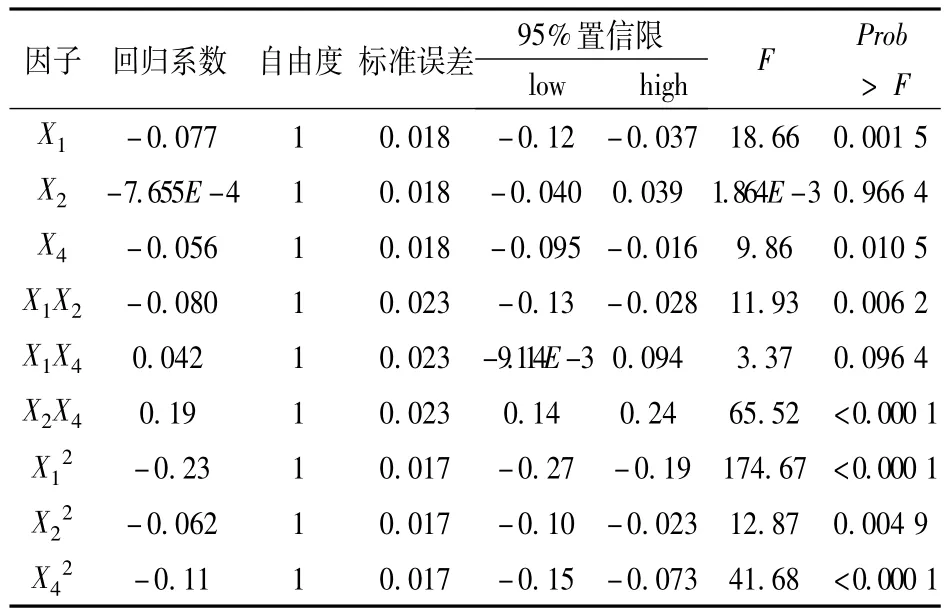
表10 蘇氨酸濃度預測回歸方程系數的顯著性檢驗
表10說明,X1對蘇氨酸濃度Y的影響達到極顯著水平,即葡萄糖濃度是對試驗結果影響較大的因子,同時X1與X2、X2與X4對蘇氨酸濃度Y的交互影響均達極顯著水平;X22、X42對酶活力Y的影響也達極顯著水平,說明葡萄糖、酶解酪素對蘇氨酸濃度的影響是二次關系。
2.3.2 響應面分析及最佳培養基成分的確定
圖1、圖2、圖3是上述二次多項式擬合出的響應曲面圖及其對應的等高線圖。
比較3組圖可知,葡萄糖濃度和(NH4)2SO4濃度、(NH4)2SO4濃度和酶解酪素濃度之間的交互作用顯著。葡萄糖濃度和酶解酪素濃度對蘇氨酸濃度的交互作用不顯著。這些直觀的分析結果與表10的分析一致,說明能真實地反應變量之間的交互作用。
圖1顯示在酶解酪素濃度一定的情況下,葡萄糖和硫酸銨對蘇氨酸濃度的交互影響效應。隨著葡萄糖濃度和硫酸銨濃度的不斷增加,蘇氨酸產量開始不斷提高,后期呈現小幅下降。葡萄糖含量在48~60g/L,酶解酪素在10.25~13.75g/L時,蘇氨酸濃度達到最高。說明一定程度上提高培養基中葡萄糖的濃度,有利于菌體生長并提高胞外蘇氨酸含量,但過高的濃度對菌體產生高滲作用,不利于菌株生長,從而影響蘇氨酸的分泌量。圖2表示在(NH4)2SO4濃度一定的情況下葡萄糖和(NH4)2SO4對蘇氨酸濃度的影響,當葡萄糖濃度為47.2~55.2g/L、酶解酪素濃度為0.78~0.85g/L時,蘇氨酸濃度最高;圖3表示在葡萄糖濃度一定情況下(NH4)2SO4和酶解酪素對蘇氨酸含量的影響,當硫酸銨濃度8.5~13g/L,酶解酪素為0.7~0.9g/L時,蘇氨酸濃度最高。

圖1 酶解酪素濃度一定時,葡萄糖和(NH4)2SO4對蘇氨酸的交互影響作用

圖2 (NH4)2SO4濃度一定時,葡萄糖和酶解酪素濃度對蘇氨酸濃度的影響

圖3 葡萄糖濃度一定時,(NH4)2SO4和酶解酪素對蘇氨酸含量的影響
方程二次項系數為負數,說明此方程有極大值,其表征的拋物面開口向下(圖1、圖2和圖3),二次回歸方程分別對X1、X2和X4求一階偏導,整理得如下三元一次方程:

解得:X1=-0.281,X2=0.899,X4=0.468;即對應的非編碼水平上3者的條件為:葡萄糖50.78g/L、(NH4)2SO415.15g/L、酶解酪素0.99g/L,此時蘇氨酸濃度預測值為3.38g/L。
2.3.3 模型驗證
以2.3.2確定的主要因子濃度配制發酵培養基,重復3次,然后測定胞外蘇氨酸含量,同時將初始發酵培養基發酵所得結果作為對照。結果顯示,蘇氨酸濃度由優化前的2.58g/L上升到3.35g/L,提高了30%,與預測值較為接近。說明回歸模型能夠真實地反映各篩選因子對發酵產蘇氨酸的影響。
3 討論
本文首先采用PB實驗設計方法,篩選出了影響重組谷氨酸棒桿菌R102ΔmetX產蘇氨酸發酵過程中的3個顯著因素:葡萄糖、硫酸銨和酶解酪素。通過最陡爬坡接近最大響應區域后,建立了以蘇氨酸濃度為響應值的二次回歸方程模型,最終得到R102ΔmetX產蘇氨酸的最優發酵條件為:葡萄糖50.78g/L,酵母粉6g/L,酶解酪素0.99g/L,生物素80μg/L,VB12 mg/L,CaCO320g/L,(NH4)2SO415.15g/L,MgSO4·7H2O 0.5g/L,KH2PO40.5g/L,K2HPO40.5g/L,FeSO4·7H2O 0.02g/L,MnSO4·7H2O 0.02g/L,接種量8%。在此條件下,實測蘇氨酸濃度為3.35g/L,與預測值3.38g/L接近,說明預測模型可靠性較高,表明PB試驗和響應面相結合的試驗方法用于優化R102ΔmetX產蘇氨酸發酵條件,不僅科學合理而且準確有效。
目前國內外利用谷氨酸棒桿菌發酵生產蘇氨酸的產量普遍較低,2009年解曉鵬等通過誘變得到一株產蘇氨酸的谷氨酸棒桿菌,其發酵產量為0.31g/L[9],Ramon等通過基因工程增強了1株谷氨酸棒桿菌產蘇氨酸的能力,達到6g/L左右[10]。本試驗僅僅對發酵培養基進行了優化,要進一步提高重組谷氨酸棒桿菌產蘇氨酸的能力,還需還可以優化發酵條件,并通過基因工程方法進一步對該重組菌加以改造獲得高產菌株。
[1]Vladimir G Debabov.The threonine story [J].Adv Biochem Eng Biotechnol,2003,79:113-l36.
[2]賈冬舒.蘇氨酸市場現狀及發展前景[J].飼料廣角,2006(1):28-29.
[3]Jorn Kalinowski,Brigitte Bathe,Daniela Bartels,et al.The complete Corynebacterium glutamicum ATCC 13032 genome sequence and its impact on the production on of L-aspartate-derived amino acids and vitamins[J].J Biotechnol,2003,104:5-25.
[4]Bastian Blombach,Mark E.Schreiner,Matthias Moch,et al.Effect of pyruvate dehydrogenase complex deficiency on L-lysine production with Corynebacterium glutamicum[J].Appl Microbiol Biotechnol,2007,76:615-623.
[5]A Miller,R R Sitter.Using the folded-over 12-run Plachett-Burman design to consider interactions[J].Technometrics,2001,43(1):44-55.
[6]王普,孫立明,何軍邀.響應面法優化熱帶假絲酵母104菌株產羰基還原酶發酵培養基[J].生物工程學報,2009,25(6):863-868.
[7]趙世光,王詩然,薛正蓮,等.Ganoderma lucidum UIM2281產漆酶發酵條件的篩選及響應面優化[J].食品與發酵工業,2009,35(11)72-77.
[8]解曉鵬,張力,楊曉志,等.L-蘇氨酸產生菌的選育[J].安徽農業科學,2009,37(22):10 417-10 419.
[9]Ramon Diesveld,Nadine Tietze,Oliver Fürst,et al.Activity of Exporters of Escherichia coli in Corynebacterium glutamicum,and their Use to Increase L-Threonine Production[J].J Mol Microbiol Biotechnol,2009,16(34):198-207.
Medium Optimization Studies for Threonine Production by Recombinant Corynebacterium glutamicum Using Response Surface Methodology
Lv Yang-yong,Wu Zhan-hong,Zheng Sui-ping
(School of Bioscience and Bioengineering,South China University of Technology,Guangzhou 510006,China)
The recombinant Corynebacterium glutamicum R102ΔmetX(R102 with deletion in the metX gene)was constructed.Fractional factorial design was used to evaluate the effects of medium components on the production of threonine.Glucose,yeast extract and enzymatic casein were the most important factors among twelve tested variables that influence the production of threonine.Based on the experimental results,the path of steepest ascent was undertaken to approach the optimal region of these factors.Central composite design and response surface analysis were subsequently employed for further optimization.The optimal medium for threonine synthesis was composed of:glucose 50.78g/L,yeast extract 6g/L,enzymatic casein 0.99g/L,biotin 80μg/L,VB12mg/L,CaCO320g/L,(NH4)2SO415.15g/L,MgSO4·7H2O 0.5g/L,KH2PO40.5g/L,K2HPO40.5g/L,FeSO4·7H2O 0.02g/L,MnSO4·7H2O 0.02g/L,inoculation quantity 8%。Under the optimum conditions,the threonine yield of the recombinant strain was 3.35g/L,which was 30%higher than the original medium components.
Corynebacterium glutamicum,L-threonine,response surface methodology,medium optimization
碩士研究生(鄭穗平副教授為通訊作者)。
*國家科技支撐計劃(2007BAK36B03),我國優勢傳統食品制造業關鍵技術研究與應用(谷氨酸發酵過程代謝流分析及菌種選育)項目資助。
2010-05-17,改回日期:2010-09-24
