16個魚腥草基因型遺傳多樣性的 SRAP分析
2010-12-05 02:59:36張丹丹曾維軍
作物研究 2010年1期
關鍵詞:體系
王 坤,鐘 軍,張丹丹,仇 萍,曾維軍
(1湖南農業大學生物科學技術學院,長沙 410128;2湖南農業大學農學院,長沙 410128;3湖南正清制藥集團股份有限公司,懷化 418000)
魚腥草(Houttuynia cordata Thunb)是三白草科蕺菜屬的宿根性多年生草本植物,在中國分布廣泛,種質資源十分豐富。巨大的種質資源數量使得人們很難對其進行深入研究并加以有效利用,且不清楚其資源遺傳多樣性的程度又影響了種質資源的育種利用[1]。
相關序列擴增多態性(sequence-related amplified polymorphism,SRAP)是一種新型的基于PCR的分子標記系統。其獨特之處在于其引物設計上,上游引物和下游引物分別包含17和18個堿基,它們能優先對基因組DNA開放閱讀框、內含子和啟動子區域進行特異擴增,因不同基因型啟動子、內含子和間隔序列長度不同而產生多態性[2],且該標記簡單、高效、高共顯性、易測序、擴增多態性高,所能提供的生物學信息更多,故而迅速在植物遺傳多樣性分析和比較基因組學研究等方面得到廣泛應用[3]。目前,SRAP標記在農作物(棉花[4]、玉米[5]、油菜[6])及蔬菜作物 (豌豆[7]、南瓜[8]、馬鈴薯[9]、蓮[10])上的應用報道較多;在果樹柿屬[11]植物、梨 屬[12]、柑 橘[13]、核 桃[14]、石 榴[15]上 的 SRAPPCR的擴增體系也有報道。但是關于SRAP標記在中草藥上的報道只有川黨參[16]、玄參[17]、黃芪[18]等少數幾種,而在魚腥草上應用研究的報道至今未見。筆者以16份不同魚腥草的資源為材料,在建立并優化其SRAP擴增反應的基礎上,利用篩選的引物對材料進行了遺傳多樣性分析,以期為揭示魚腥草資源間的系譜關系和遺傳多態性提供新的依據。
1 材料和方法
1.1 材料
試驗材料于2009年8月取自湖南正清制藥集團股份有限公司懷化魚腥草種質資源圃(表1)。
1.2 儀器與試劑
UV-55紫外分光光度計(Bio-RAD),Gel Doc2000凝膠成像系統(Bio-RAD),Eppendorf Mastercycle gradient梯度 PCR儀 (Eppendorf),DYCZ-24B型電泳槽(北京市六一儀器廠),DYY-6C型電泳儀(北京市六一儀器廠),s700三星數碼相機。
瓊脂糖(進口分裝),SRAP引物(北京賽百盛基因技術有限公司 ),PVP(Sigma),β-巰基乙醇(Sigma),CT AB(Amresco),100 bpDNA laddermarker(TOYOBO),丙烯酰胺、甲叉-雙丙烯酰胺、硝酸銀、甲醛均為 BBI公司產品,T EMED、過硫酸銨為 Promega產品,Taq酶(TOYOBO),dNT Ps(TaKaRa),其余為國產分析純試劑。
1.3 方法
(1)DNA的提取。魚腥草葉片 DNA的提取采用鐘軍改良的CTAB法[19],并稍作改動。經 UV-55紫外分光光度計分析純度和產率,用0.8%瓊脂糖電泳檢測其質量。
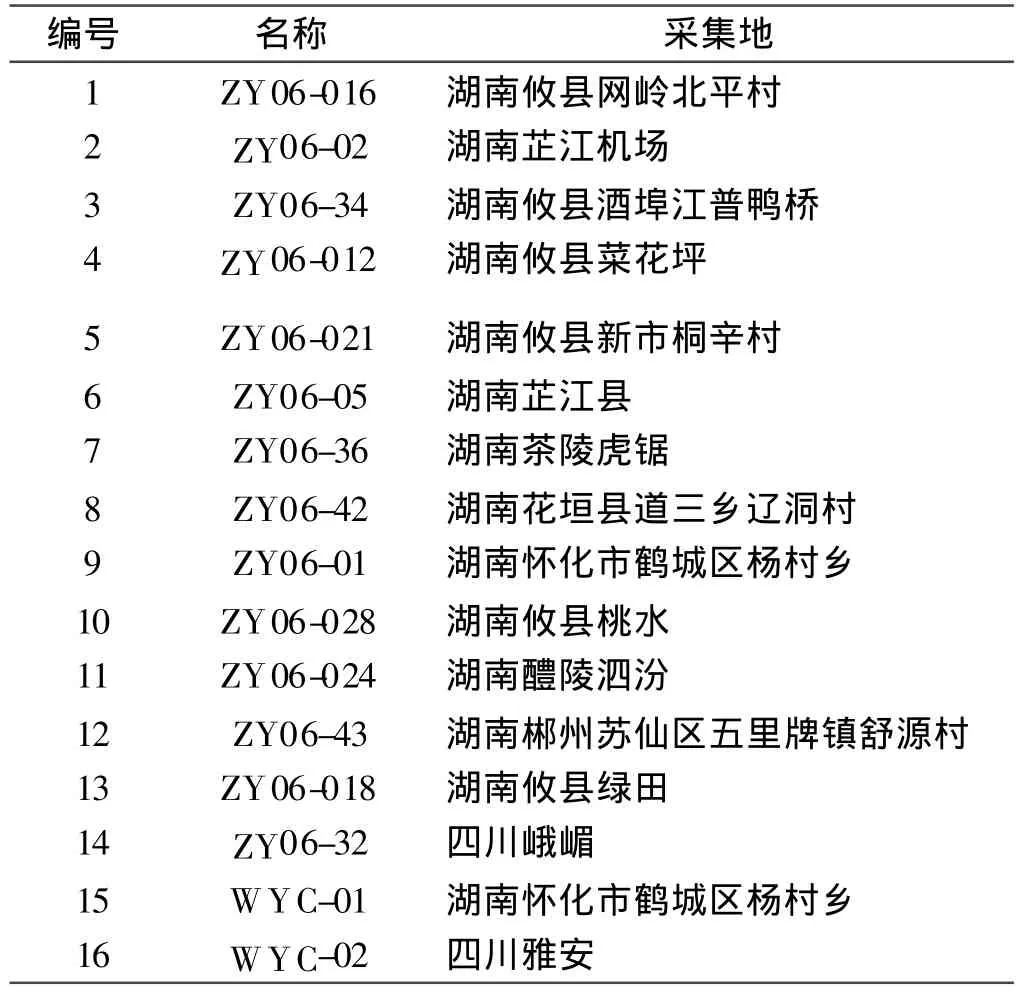
表1 魚腥草材料編號及來源
(2)SRAP反應體系的優化。每一組均以 ZY06-012 DNA為模板,以 Me5— Em4為引物組合,分別對魚腥草 PCR反應體系中5個主要影響因素(dNTPs的濃度、模板 DNA的濃度、引物濃度、Taq酶的用量和Mg2+濃度)和擴增體系中 2個主要影響因素(退火溫度和循環數)進行正交設計優化。正交試驗設計分別見表 2和表 3。
(3)篩選引物。選取已公布的 SRAP上游引物18條,下游引物20條(表4),組成360對引物組合,由北京賽百盛基因技術有限公司合成。篩選出擴增多態性好、條帶清晰的引物對進行遺傳多樣性分析。

表2 魚腥草SRAP-PCR反應體系正交試驗設計表L16(45)

表3 魚腥草SRAP-PCR擴增體系正交試驗設計表L9(34)
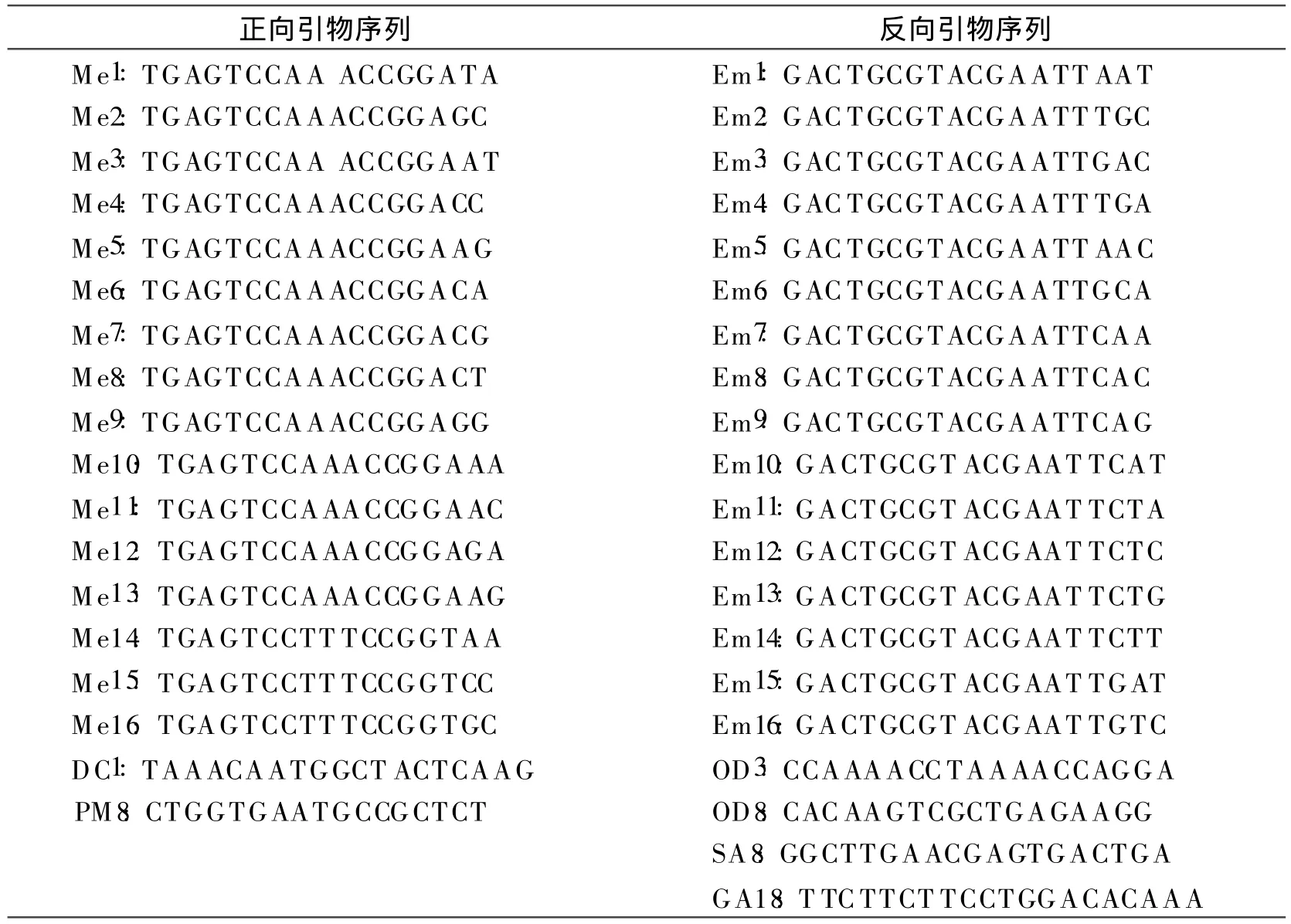
表4 試驗中使用的SRAP引物
(4)SRAP擴增。以優化后的最佳反應體系和擴增體系進行,最后取2μL的擴增產物,8%丙烯酰胺凝膠電泳,電壓 150 V,電泳1.5 h,然后固定、染色、顯影、停顯[20],最后用數碼相機拍照并記錄。
1.4 統計分析
選取同一引物在相同基因位點擴增重復性強和清晰的條帶,根據條帶的有無分別記做“1”和“0”。不同引物擴增的結果構成原始的“0,1”二元數據矩陣,用DPS分析軟件計算魚腥草各基因型之間的 Nei’s遺傳距離,用 UPGMA程序構建聚類圖。
2 結果與分析
2.1 SRAP-PCR反應體系及擴增體系正交試驗優化分析
從圖1中可以看出,在 16組反應中只有第 4組的電泳結果最為清晰可辨且擴增條帶數最多,第 3組稍次之,即適合于魚腥草 SRAP最優的反應體系為:25 μL的反應體系中,模板 DNA量 40 ng、Mg2+濃度 2.0 mmol/L、上下游引物 37.5 ng/L、dNTPs0.1 mmol/L以及 2 U Taq酶。從圖 2結果來看,第 9組的條帶比其他組更清楚,條帶數更多,所以適合于魚腥草SRAP最優的擴增體系為第 9組擴增體系,即退火溫度 53℃,循環數為 35。
2.2 遺傳多樣性分析
從360對引物組合中篩選出118對擴增條帶清晰、重復性好的引物組合進行 PCR擴增,共擴增出 7 582個條帶,其中多態性條帶 6 590個,多態率 86.92%。SRAP標記的多態性見表5,擴增產物的分子量為50~2 500 bp。 其中引物組合 Me9-GA18共擴增出 79個基因位點,多態性位點63,多態位點比率為79.75%;引物組合Me16-Em9共擴增出基因位點105個,其中多態性位點73個,多態位點比率為 69.52%。
2.3 魚腥草基因型SRAP特異性缺失標記及特異性標記
在 16個魚腥草基因型中,ZY06-028魚腥草在Me9-GA18(圖3)引物對擴增時缺失一條250 bp左右擴增帶,可以作為ZY06-028魚腥草品種鑒定的特異標記。而 ZY06-016和 ZY06-028魚腥草約在550 bp處共同缺失一條擴增帶,其余品種均無缺失。在 Me8-Em10引物擴增產物中出現了兩個特異性標記的條帶,分別是 ZY06-42在 300 bp處的一條帶和 ZY06-01在 450 bp處的一條帶(圖4)。這些特異性缺失標記和特異性標記都可以作為魚腥草品種鑒定和形態學分類的重要依據。
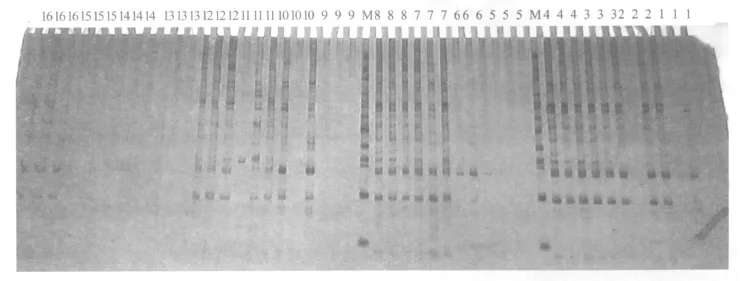
圖1 魚腥草SRAP-PCR反應體系正交試驗設計優化結果
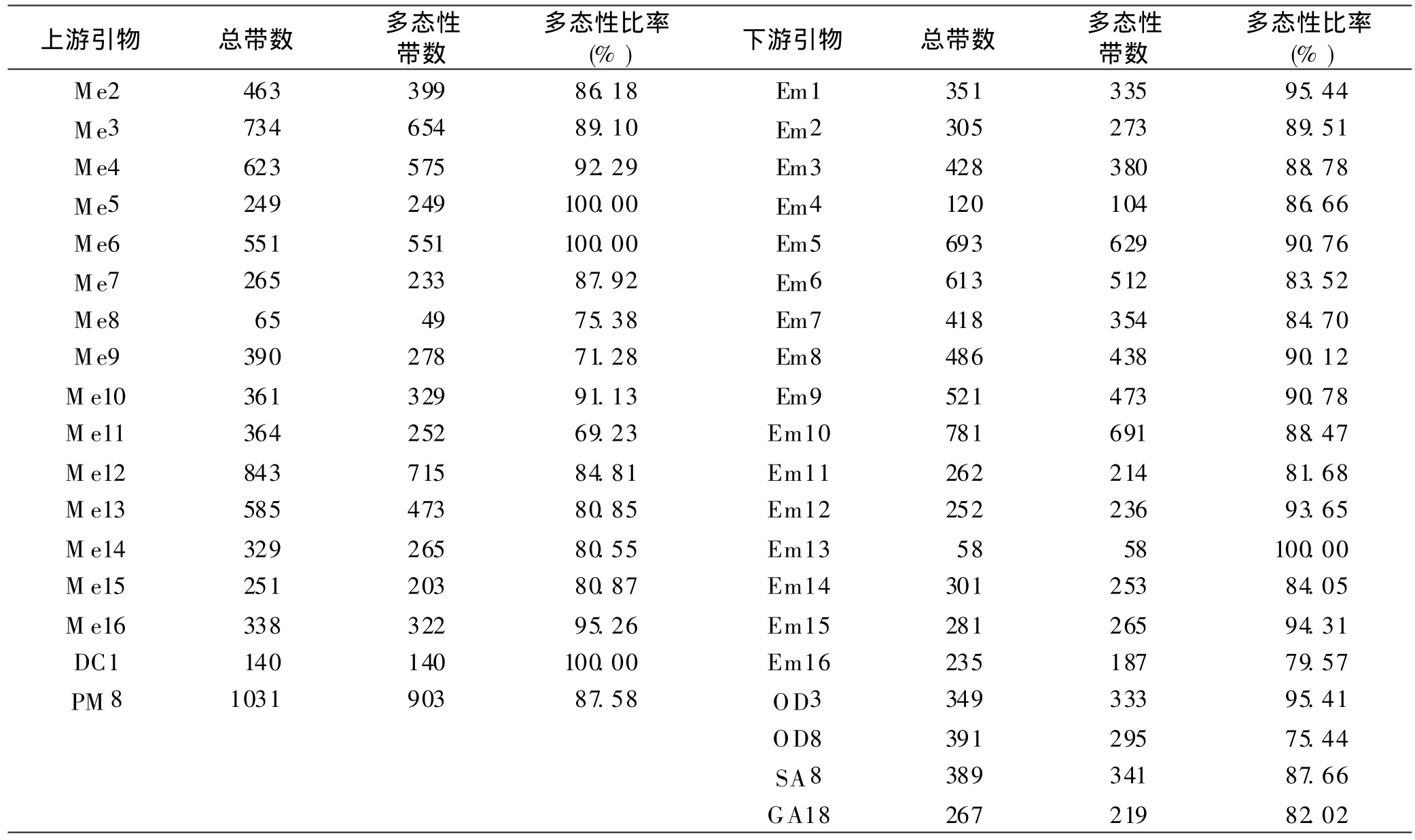
表5 SRAP上、下游引物、擴增總帶數及多態性帶數
2.4 基因型間的親緣關系
應用 DPS軟件,根據“0,1”數據矩陣,計算的遺傳距離在 0.0536~0.4040。其中 ZY06-021和 ZY06-05之間的遺傳距離最小為 0.0536,ZY06-016和 ZY06-024之間的遺傳距離最遠為0.4040。ZY06-016為在湖南攸縣網嶺北平村采集的材料,ZY06-024為在湖南醴陵泗汾采集的材料。圖5為應用UPGMA程序建立的16個魚腥草基因型的聚類圖。根據聚類圖可看出,在遺傳距離 0.31處,可將 16份魚腥草材料分為三大類,其中ZY06-024魚腥草為第一類,ZY06-01為第二類,第三類包括了其余的14份材料,這14份材料包括了兩個亞類,第一亞類中包含有 ZY06-016,ZY06-02,ZY06-021,ZY06-05,ZY06-36,ZY06-42,ZY06-028,共 7份材料;第二亞類中包含有 ZY06-34,ZY06-012,ZY06-43,ZY06-018,ZY06-32,WYC-01,WYC-02,也是 7份材料。在16份材料中遺傳關系較近的有 ZY06-021和ZY06-05,ZY06-43和 ZY06-018,WYC-01和 WYC-02。從此結果可以看出,同一地區的品種,如湖南攸縣的 4份資源:ZY06-016,ZY06-012,ZY06-021,ZY06-018并沒有聚到一類,且同為紅稈的ZY06-05和ZY06-01的遺傳關系還比較遠。這或許能夠說明魚腥草基因型間的遺傳關系很復雜。

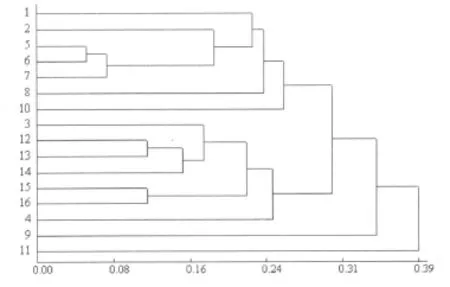
圖5 16份魚腥草品種資源的SRAP聚類圖
3 討 論
本試驗中選擇118對引物組合,對 16個魚腥草品種資源進行了多樣性分析,其揭示的多態性位點數高達 86.92%。由此可見,作為一種新型的分子標記,SRAP技術在魚腥草資源的系統分類、親緣關系鑒定、品種鑒別、親本選擇等方面具有潛在的應用前景。
SRAP標記是一種基于 PCR技術的分子標記,擴增結果易受到反應條件及擴增條件的影響,因此針對不同的試材及實驗條件需要優化反應體系。本試驗進行了不同影響因子的正交試驗設計優化,試驗結果表明,適合于魚腥草 SRAP最優的反應體系為:25μL的反應體系中,模板DNA量40 ng,Mg2+濃度 2.0 mmol/L,上下游引物 37.5 ng/L,dNTPs0.1 mmol/L以及 2 U Taq酶;適合于魚腥草SRAP最優的擴增體系為:退火溫度 53℃,循環數為 35。此結果與南瓜[8]、柿子[11]、梨[12]等建立的 SRAP標準反應體系中的各參數相比,存在一定的差別。其原因可能是不同物種的基因組存在很大的差別,所需的反應體系也會有所不同,也可能是不同公司的藥品儀器導致的差別。
本實驗在對 16個魚腥草品種的 SRAP分析中發現,有些品種用特定的引物對擴增以后,會出現一些特異性缺失的條帶及特異性的條帶,這些特異性缺失條帶和特異性的條帶可為魚腥草的品種鑒定提供必要的依據。例如,引物對組合Me9-GA18-250 bp缺失條帶是 ZY06-028魚腥草的特異缺失標記,Me9-GA18-550 bp缺失條帶是ZY06-016和ZY06-028魚腥草的特異缺失標記,在引物對組合Me8-Em10中ZY06-42和 ZY06-01分別在 300 bp和 450 bp處出現了一條特異性的條帶。
16個魚腥草基因型中 ZY06-016和 ZY06-024的遺傳關系最遠,ZY06-021和 ZY06-05,ZY06-43和ZY06-018,WYC-01和 WYC-02遺傳關系較近,同時可在遺傳距離0.31處分為3個類群,其中 ZY06-024和ZY06-01魚腥草與其它基因型顯著不同,分別單獨聚為一類,其余的 14個魚腥草品種聚為一大類,而這一大類又可在遺傳距離0.28處分為兩個亞類。從聚類結果中可以發現,同一地區的品種(如湖南攸縣的 4個品種:ZY06-016,ZY06-012,ZY06-021,ZY06-018)沒有聚到一起,不僅如此,與形態學指標莖稈顏色、莖稈長度等分類也有很大不同。吳衛[1]等人對四川地區92份魚腥草材料進行研究,發現魚腥草染色體數目的變化范圍很大,在 36~126之間變化,且即使在同一地區不同村鎮上的魚腥草染色體數目亦有很大差別。例如,同是四川劍閣的魚腥草材料,但染色體數目就有 72條,82條,84條,86條之別。這一結論或許能對上述結果做出較好的解釋,即魚腥草基因型間的遺傳關系非常復雜。
4 結 論
SRAP分子標記能夠很好的用于魚腥草基因型遺傳多樣性分析。 ZY06-024和 ZY06-01與其他 14份材料的基因型顯著不同,且這 16份材料的分子標記結果表明,在同一地區引進的材料并沒有很好聚到一類中,這或許能夠說明魚腥草的基因型受生態環境的影響比較小。同時各品種間聚類結果與按照莖稈顏色、莖稈長度等形態學指標分類也不太一致(未發表),表明了魚腥草各基因型遺傳關系的復雜性。
[1]吳 衛.魚腥草種質資源研究 [D].雅安:四川農業大學,2002.
[2]Li G,Quiros CF.Sequence-related amplified polymorphism(SRAP),a new marker system based on a simple PCR reaction:Itsapplication tomapping and gene tagging inBrassica[J].Theor Appl Genet,2001,103:455-460.
[3]李 莉,彭建營,白瑞霞,等.SRAP與 TRAP標記及其在園藝植物研究中的應用[J].西北植物學報,2006,26(8):1749-1753.
[4]李 武,林忠旭,張獻龍.海島棉遺傳多樣性的 SRAP標記分析 [J].作物學報,2007,34(7):634-638.
[5]姜樹坤,馬 慧,劉 君,等.利用 SRAP標記分析玉米遺傳多樣性[J].分子植物育種,2007,5(3):412-415.
[6]Riaz A,Li G,Quresh Z,et al.Genetic diversity of oilseed Brassica napus inbred lines based on sequence-related amplified polymorphism and its relation to hybrid performance[J].Plant Breeding,2001,120(5):411-415.
[7]Espostio M A,Martin EA,Cravero V P,et al.Characterization of pea accessions by SRAP markers[J].Scientia Horticulture,2007,113(4):329-333.
[8]Ferriol M,Pioo B,Nuez F.Genetic diversity of a germplasm collection of Cucurbita pepousingSRAP and AFLP markers[J].Theor Appl Genet,2003,107:271-276.
[9]何鳳發,楊志平,張正圣,等.馬鈴薯遺傳資源多樣性的SRAP分析 [J].農業生物技術學報,2007,15(6):1001-1006.
[10]瞿 楨,魏英輝,李大威 ,等.蓮品種資源的 SRAP遺傳多樣性分析[J].氨基酸和生物資源,2008,30(3):21-24.
[11]郭大龍,羅正榮.部分柿屬植物 SRAP-PCR反應體系的優化[J].果樹學報,2006,23(1):138-142.
[12]張妤艷,吳 俊,張紹鈴.梨 SRAP-PCR反應體系的建立與優化 [J].農業生物技術學報,2007,15(5):909-914.
[13]吳 鑫,雷天剛 ,何永睿,等.柑桔 SRAP和 ISSR分子標記技術體系的建立與優化 [J].分子植物育種,2008,6(1):170-175.
[14]馬 明,楊克強,劉曉菊 ,等.核桃 SRAP標記反應體系建立的研究[J].山東農業大學學報,2007,38(2):189-194.
[15]張四普,汪良駒,曹尚銀 ,等.23個石榴基因型遺傳多樣性的 SRAP分析 [J].果樹學報,2008,25(5):655-660.
[16]陳大霞,彭 銳 ,李隆云,等.利用 SRAP和 ISSR標記分析川黨參的遺傳多樣性 [J].中國中藥雜志,2009,34(3):257-262.
[17]陳大霞,李隆云,彭 銳,等.玄參3種栽培類型遺傳關系和遺傳多樣性的 SRAP研究[J].中國中藥雜志,2009,34(2):140-146.
[18]錢 丹,黃璐琦,崔光紅,等.不同產地蒙古黃芪遺傳關系的 SRAP分析 [J].中國中藥雜志,2009,34(4):384-389.
[19]鐘 軍.蕓芥抗油菜菌核病分子基礎的研究 [D].長沙:湖南農業大學,2003.
[20]趙 培,王振英,彭永康,等.瓊脂糖和聚丙烯酰胺凝膠電泳技術檢測小麥基因組 DN ARAPD擴增產物的方法學比較[J].中國生物工程雜志,2003,23(8):99-104.
猜你喜歡
商品與質量(2021年43期)2022-01-18 05:31:22
杭州(2020年23期)2021-01-11 00:54:42
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
中國衛生(2015年12期)2015-11-10 05:13:40
現代企業(2015年1期)2015-02-28 18:43:18
汽車零部件(2014年5期)2014-11-11 12:24:28
新高考·高一物理(2014年1期)2014-09-18 01:26:07
浙江人大(2014年1期)2014-03-20 16:19:53
終身教育研究(2012年4期)2012-03-25 10:41:11
