鱔藤等4種藤本植物耐鹽性試驗
2011-05-08 09:35:08杜月青劉建強陳立潮金水虎
浙江林業科技 2011年4期
杜月青,劉建強,陳立潮,金水虎
(浙江天泰園林建設有限公司,浙江 紹興 312000)
鱔藤等4種藤本植物耐鹽性試驗
杜月青,劉建強,陳立潮,金水虎*
(浙江天泰園林建設有限公司,浙江 紹興 312000)
摘要:以鱔藤(Anodendron affine)、山蒟(Piper hancei)、風藤(Piper kadsura)和厚藤(Ipomoea pes-caprae)為試材,研究在鹽脅迫下4種藤本植物形態特征和生理生化指標的變化。結果表明:4種藤本植物的細胞膜透性均隨鹽脅迫時間的延長而升高,同時鹽濃度越大的處理其細胞膜透性越高;山蒟、風藤和厚藤的POD活性隨鹽濃度的升高而升高,鱔藤的POD活性基本隨鹽濃度的升高和處理時間的延長而逐漸下降;風藤、鱔藤和厚藤的游離脯氨酸隨鹽脅迫時間的延長而升高,山蒟的游離脯氨酸隨鹽脅迫時間的延長呈先升后降的趨勢。4種藤本植物的可溶性蛋白含量基本隨鹽脅迫濃度的升高而呈先升高然后迅速下降并趨于穩定的趨勢,并且鹽脅迫時間越長其可溶性蛋白含量越低;4種植物的MDA含量隨鹽濃度的上升而增加,處理時間上以處理20 d的MDA含量最高。
關鍵詞:鹽脅迫;生理生化指標;鱔藤;山蒟;風藤;厚藤
藤本植物垂直綠化已經成為我國園林生態景觀建設的新熱點。特別在上海市,垂直綠化己經獲得成功。同時,垂直綠化在北京、深圳、南京等城市也有較快的發展,許多藤本植物在立體綠化中得到了較好的利用[1~2]。
我國東部大部分發達城市處在沿海地區,這些地區分布有大面積的濱海鹽漬土。淡水資源缺乏、土壤鹽堿化、植被覆蓋率底、環境質量需要改善是這些城市發展的重要問題。另外,在我國的一些內陸地區,尤其是東北、西北地區,由于對水土、草原、森林等資源的過度開發利用,致使某些地區沙漠化、鹽堿化問題特別嚴重,引種耐鹽堿植物,恢復地被成為這些地區的當務之急[3~5]。
1 材料與方法
1.1 材料與處理
試驗材料為鱔藤(Anodendron affine)、山蒟(Piper hancei)、風藤(Piper kadsura)和厚藤(Ipomoea pes -caprae)的一年生扦插苗和實生苗。用20 cm×20 cm底部有孔的營養缽,下面設托盤,裝入濕度和重量基本一致的基質(基質為珍珠巖、泥炭和蛭石按2:3:1的比例配制),選擇生長良好、長勢基本一致的植株進行移栽;每種藤本植物選擇15盆,置于智能溫室內緩苗一個月后開始進行鹽脅迫處理,處理鹽(NaCl)濃度設定為:0、0.3%、0.6%、0.9%和1.2%,每隔4 d定時定量澆灌一定濃度的鹽溶液;于第5天、第10天、第15天和第20天分別測定相關生理生化指標。試驗重復3次。按時均勻取樣,葉片剪后置于冰袋中,樣品均當天處理分析完畢。
1.2 方法
電導儀法測定細胞膜透性、愈瘡木酚比色法測定過氧化物酶(POD)活性、考馬斯亮藍G-250染色法測定可溶性蛋白含量、茚三酮染色法測定游離脯氨酸含量、硫代巴比妥酸法測定丙二醛含量[6]。
試驗數據用Excel2003和SAS9.0進行分析,并進行差異性比較,采用模糊數學的隸屬函數法綜合評價4種藤本植物的耐鹽能力。
2 結果與分析
2.1 鹽脅迫對4種藤本植物細胞膜透性的影響
細胞受鹽害后,細胞膜遭受傷害,透性變大。大量研究表明,植物在逆境脅迫或衰老過程中,細胞內活性氧代謝的平衡被破壞而有利于自由基的超量積累,過剩自由基的毒害之一就是引發或加劇膜脂過氧化作用,造成細胞膜系統和代謝過程的損傷。在鹽分脅迫下,相對電導率增幅小,表明植物細胞膜系統結構受損程度低,從而其耐鹽性就高[7~9]。
由圖1、圖2、圖3、圖4表明,4種藤本植物的相對電導率均隨脅迫時間的延長而升高,同時鹽濃度越大的處理其相對電導率也越高,到第20天時,山蒟、風藤、鱔藤和厚藤在不同鹽濃度處理的相對電導率平均分別是ck的2.55倍、2.75倍、3.11倍、2.30倍。
采用多重比較分析可知:山蒟、風藤和厚藤之間差異不顯著,鱔藤和其它植物之間差異極顯著;4種植物的相對電導率均隨鹽濃度的提高和處理時間的延長而增加。
根據上述分析可知,4種藤本植物的耐鹽能力強弱依次為:厚藤 > 山蒟 > 風藤 > 鱔藤。
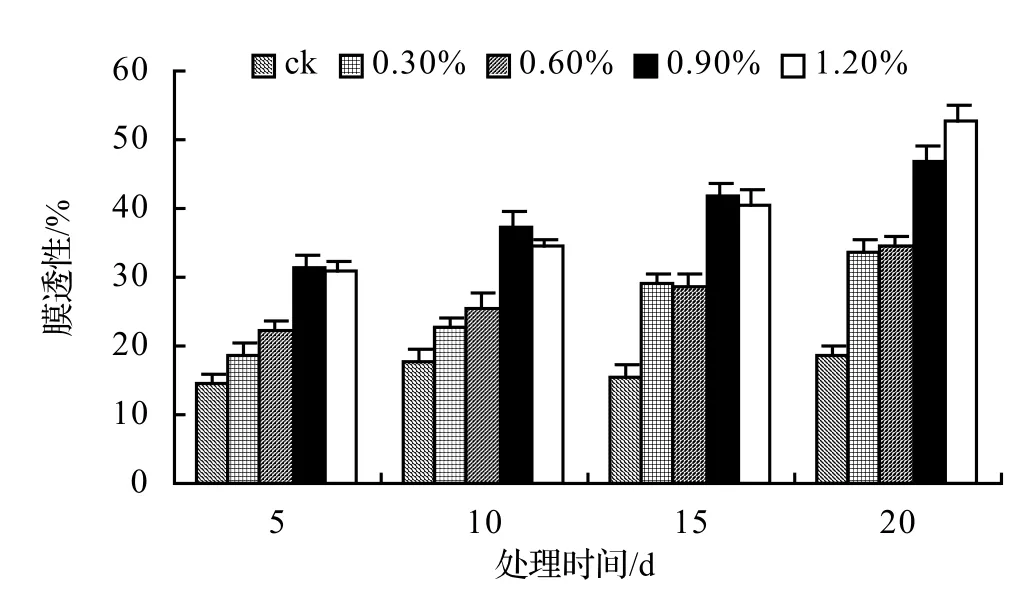
圖1 鹽脅迫對山蒟相對電導率的影響Figure 1 Effect of salt stress on relative conductivity of P. hancei
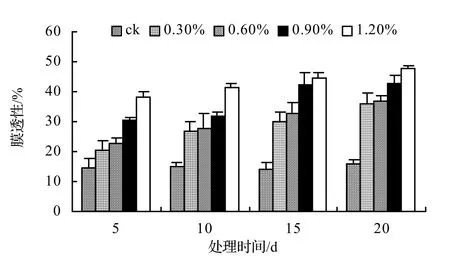
圖2 鹽脅迫對風藤相對電導率的影響Figure 2 Effect of salt stress on relative conductivity of P. kadsura
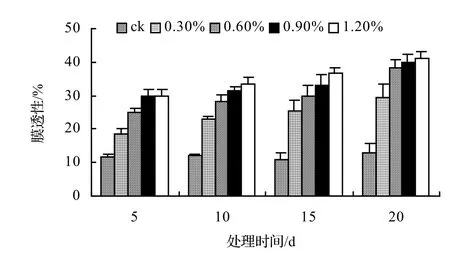
圖3 鹽脅迫對鱔藤相對電導率的影響Figure 3 Effect of salt stress on relative conductivity of A. affine
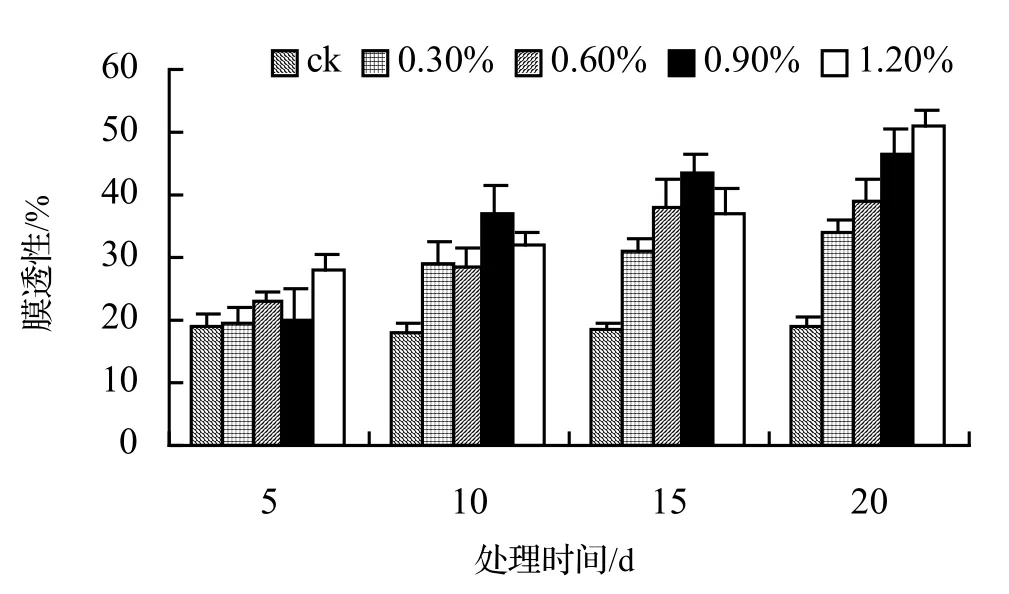
圖4 鹽脅迫對厚藤相對電導率的影響Figure 4 Effect of salt stress on relative conductivity of I. pes-caprae
2.2 鹽脅迫對4種藤本植物POD活性的影響
在鹽脅迫下POD活性變化目前尚無定論,鹽脅迫后POD活性表現出既有升高也有降低,并且耐鹽品種比鹽敏感品種有較高的POD活性[10],還有一些植物體內POD活性隨鹽濃度的增大而增強[11~12];而有些植物在不同濃度鹽脅迫下POD活性隨鹽濃度增加呈波動性變化[13]。
由圖5、圖6、圖7、圖8表明,山蒟、風藤和厚藤的POD活性隨鹽濃度的升高而升高,并且在脅迫處理第10天時達到最大值,然后迅速下降;鱔藤的POD活性基本隨鹽濃度的升高和處理時間的延長而逐漸下降。處理第10天時,山蒟在0.3%、0.6%和1.2%鹽脅迫濃度下的POD活性達到最大值,其中1.2%鹽濃度下其POD活性是ck的4.90倍;風藤在鹽脅迫處理第5天時的POD活性迅速上升,0.3%、0.6%和1.2%鹽濃度處理在第10天時達到最大值,其中1.2%鹽濃度下其POD活性是ck的4.89倍;鱔藤在0.3%鹽濃度處理第5天時達最大值,隨后迅速下降,到第20天時下降了72.44%,1.2%鹽濃度處理在第5天時達最大值,隨后迅速下降,到第20天時下降了66.66%;厚藤4個鹽濃度處理在第5天時同時達到最大值,鹽濃度越高其POD活性也越高,隨后迅速下降。
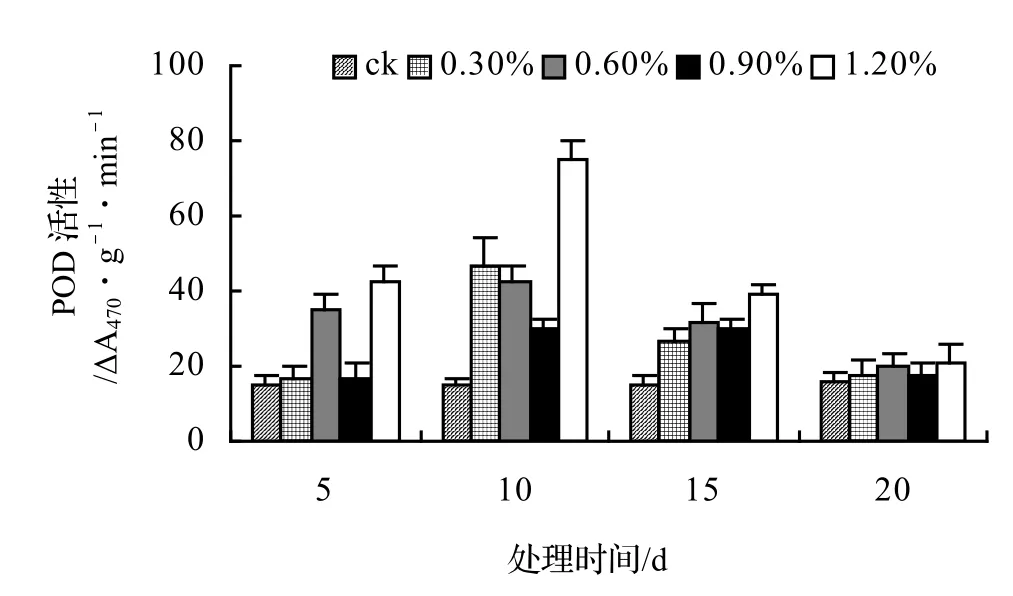
圖5 鹽脅迫對山蒟POD活性的影響Figure 5 Effect of salt stress on POD of P. hancei
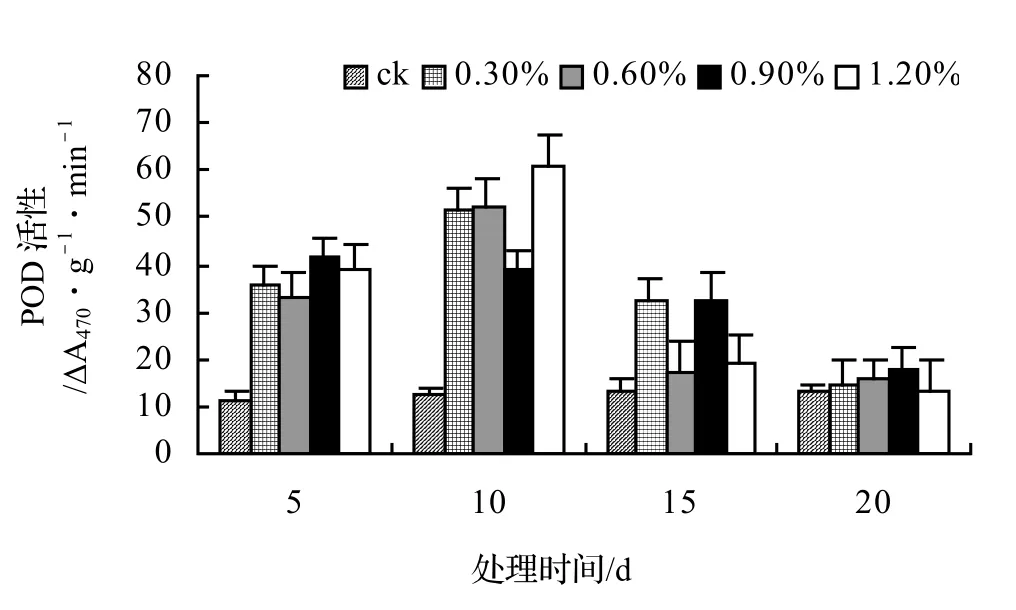
圖6 鹽脅迫對風藤POD活性的影響Figure 6 Effect of salt stress on POD of P. kadsura
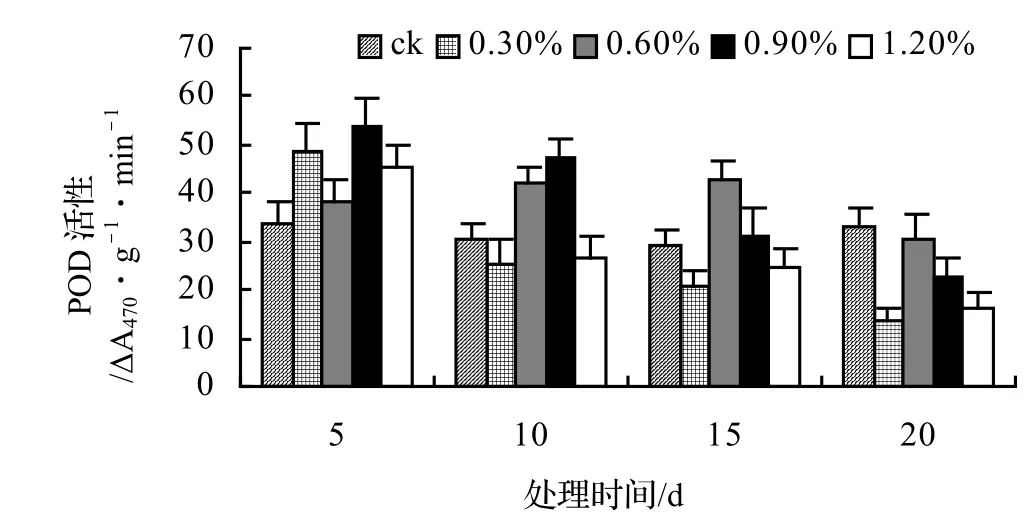
圖7 鹽脅迫對鱔藤POD活性的影響Figure 7 Effect of salt stress on POD of A. affine

圖8 鹽脅迫對厚藤POD活性的影響Figure 8 Effect of salt stress on POD of I. pes-caprae
采用多重比較分析可知:山蒟和風藤之間差異不顯著,厚藤和鱔藤之間差異顯著,其中厚藤具有較高的POD活性,風藤最低;POD活性隨鹽濃度的升高而升高,0.3%和0.6%鹽濃度之間差異不顯著,0.9%和1.2%鹽濃度之間差異不顯著。
根據上述分析可知,4種藤本植物的耐鹽能力強弱依次為:厚藤 > 鱔藤 > 山蒟 > 風藤。
2.3 鹽脅迫對4種藤本植物游離脯氨酸含量的影響
目前把游離脯氨酸積累的作用大致歸結為以下幾個方面:可以作為細胞的有效調節物質[14];具有保護酶和膜的作用[15];可以直接利用的無毒形式的氮源,作為能量和呼吸底物,參與葉綠素的合成等,從脯氨酸在逆境條件下積累的途徑來看,它既可能有適應性的意義,又可能是細胞結構和功能受損傷的表現,是一種傷害反應[16~17]。
由圖9、圖10、圖11、圖12表明,風藤、鱔藤和厚藤的游離脯氨酸隨鹽脅迫時間的延長而升高,山蒟的游離脯氨酸隨鹽脅迫時間的延長而呈現出先升高后降低的趨勢。山蒟在各鹽濃度處理至第15天時其游離脯氨酸含量達到最大值,其中0.3%為最高值,是ck的3.15倍,風藤游離脯氨酸在1.2%鹽濃度處理至第20天時其游離脯氨酸含量為各處理中的最大值,是ck的3.07倍,鱔藤在鹽脅迫初期各鹽濃度之間差異不顯著,到第20天時,其游離脯氨酸含量迅速增加,其中1.2%鹽濃度處理為最高值,是ck的3.61倍,厚藤游離脯氨酸含量隨鹽脅迫的時間延長而升高,到第20天時,0.6%鹽濃度處理達到最高值,是ck的3.76倍。

圖9 鹽脅迫對山蒟游離脯氨酸含量的影響Figure 9 Effect of salt stress on free proline of P. hancei
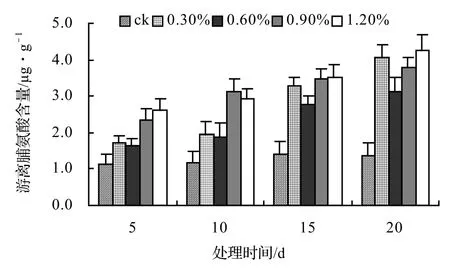
圖10 鹽脅迫對風藤游離脯氨酸含量的影響Figure 10 Effect of salt stress on free proline of P. kadsura
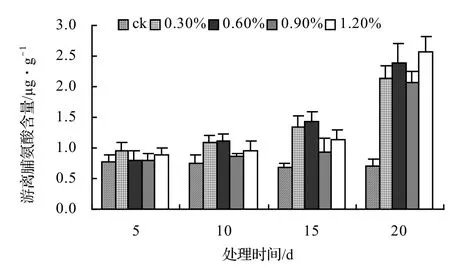
圖11 鹽脅迫對鱔藤游離脯氨酸含量的影響Figure 11 Effect of salt stress on free proline of A. affine
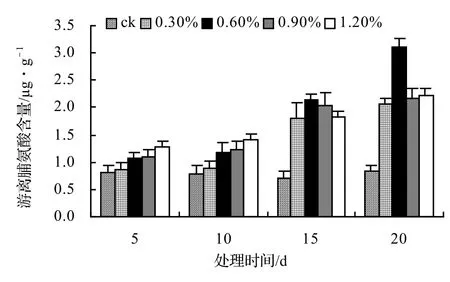
圖12 鹽脅迫對厚藤游離脯氨酸含量的影響Figure 12 Effect of salt stress on free proline of I. pes-caprae
采用多重比較分析可知:0.3%和0.6%鹽濃度處理之間差異不顯著,0.9%處理和其它處理之間差異極顯著,各鹽濃度處理之間以1.2%鹽濃度處理最高,0.9%最低。
根據上述分析可知,4種藤本植物的耐鹽能力強弱依次為:厚藤 > 鱔藤 > 山蒟 > 風藤。
2.4鹽脅迫對4種藤本植物可溶性蛋白含量的影響
逆境下植物可溶性蛋白的變化與植物的抗逆性有一定的關系。許多資料表明,在鹽脅迫下,植物體內的可溶性蛋白含量隨著鹽濃度的增加呈下降趨勢[18]。徐云嶺卻發現可溶性蛋白并不隨鹽處理濃度的增大而呈直線變化;可溶性蛋白在不同的樹種、不同發育階段起的作用是不同的[19]。
由圖13、圖14、圖15、圖16表明,4種藤本植物的可溶性蛋白含量基本上隨鹽脅迫濃度的升高而呈現出先升高后下降并趨于穩定的趨勢,并且鹽脅迫時間越長其可溶性蛋白含量也越低,在后期其不同鹽濃度之間表現不顯著。山蒟在第10天時達到最大值,隨后開始下降,到第20天時各鹽濃度處理平均比ck下降43.53%;風藤在第10天時為最大值,隨后便迅速下降,到第20天時各鹽濃度處理平均比ck下降43.59%;鱔藤在第10天時為最大值,隨后便迅速下降,到第20天時各鹽濃度處理平均比ck下降39.29%;厚藤在第10天時為最大值,隨后便迅速下降,到第20天時各鹽濃度處理平均比ck下降37.06%。
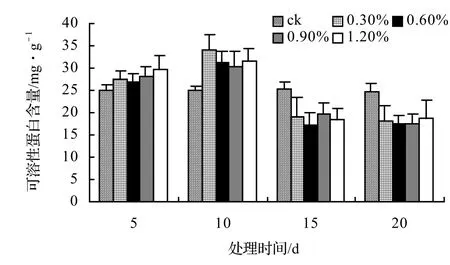
圖13 鹽脅迫對山蒟可溶性蛋白含量的影響Figure 13 Effect of salt stress on soluble protein conten of P. hancei

圖14 鹽脅迫對風藤可溶性蛋白含量的影響Figure 14 Effect of salt stress on soluble protein conten of P. kadsura

圖15 鹽脅迫對鱔藤可溶性蛋白含量的影響Figure 15 Effect of salt stress on soluble protein conten of A. affine
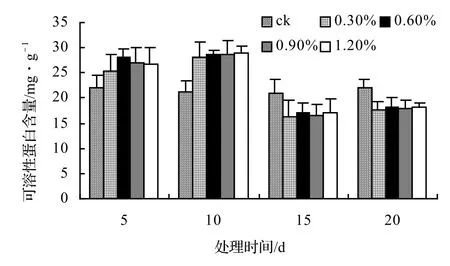
圖16 鹽脅迫對厚藤可溶性蛋白含量的影響Figure 16 Effect of salt stress on soluble protein conten of I. pes-caprae
采用多重比較分析可知:風藤、鱔藤和山蒟、風藤之間差異不顯著,山蒟和厚藤之間差異極顯著;各鹽濃度之間差異不顯著,可溶性蛋白隨鹽濃度的升高而降低;鹽脅迫處理第10天達到最大值,第5天和第10天、15天之間差異極顯著,第15天和第20天之間差異不顯著。
根據上述分析可知,4種藤本植物的耐鹽能力強弱依次為:厚藤>鱔藤>山蒟>風藤。
丙二醛(MDA)被認為是膜脂過氧化的產物,其含量與細胞膜系統的傷害程度密切相關[20~21]。隨著鹽脅迫強度的加大,MDA含量逐漸升高,MDA的含量大小可衡量膜質過氧化程度。在鹽分脅迫下,MDA含量增幅小,表明植物抗氧化脅迫的能力強,從而其耐鹽性就強。
由圖17可知:山蒟的MDA含量在鹽脅迫第5天時達到最大值,隨后便開始下降,各鹽脅迫處理的平均MDA含量約是ck的1.78倍。由圖18可知:風藤的平均MDA含量約是ck的1.45倍。由圖19可知:鱔藤各處理的平均MDA含量約是ck的1.19倍。由圖20可知:厚藤各處理的平均MDA含量約是ck的1.17倍。
采用多重比較分析可知:不同植物之間差異極顯著,其MDA總量大小為:風藤 > 鱔藤 > 山蒟 > 厚藤;其中0.6%和0.9%鹽濃度處理之間差異不顯著,0.3%和1.2%鹽濃度處理之間差異極顯著;處理時間以處理第20
2.5 鹽脅迫對4種藤本植物丙二醛含量的影響天的MDA含量最高,第15天最小,第10天和第20天之間差異不顯著,第5天和第15天差異不顯著。
根據上述分析可知,4種藤本植物的耐鹽能力強弱依次為:厚藤 > 鱔藤 > 風藤 > 山蒟。
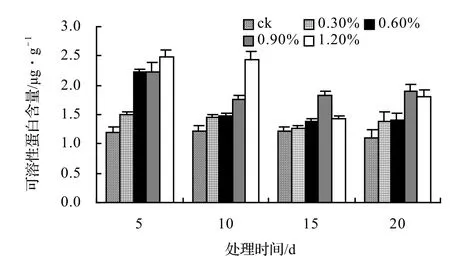
圖17 鹽脅迫對山蒟丙二醛含量的影響Figure 17 Effect of salt stress on MDA conten of P. hancei
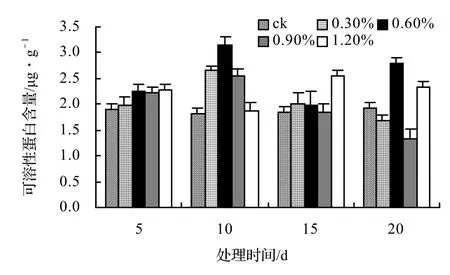
圖18 鹽脅迫對風藤丙二醛含量的影響Figure 18 Effect of salt stress on MDA conten of P. kadsura

圖19 鹽脅迫對鱔藤丙二醛含量的影響Figure 19 Effect of salt stress on MDA conten of A. affine

圖20 鹽脅迫對厚藤丙二醛含量的影響Figure 20 Effect of salt stress on MDA conten of I. pes-caprae
2.6 4種藤本植物耐鹽性綜合評價
植物的耐鹽性是一個受多種因素影響的較為復雜的綜合性狀,多種因素的綜合作用才促進耐鹽性的形成,因此對于植物耐鹽性的評價應該用盡可能多的指標來綜合評價,從而彌補與緩和單個指標對于評定植物耐鹽性所造成的片面性。本文選取細胞膜透性、POD活性、游離脯氨酸含量、丙二醛含量、可溶性蛋白含量5個指標,采用模糊數學的隸屬函數法對4種藤本植物的耐鹽性進行綜合評價[22~25]。
由表1可知,4種藤本植物的耐鹽能力大小依次為:厚藤 > 鱔藤 > 山蒟 > 風藤。
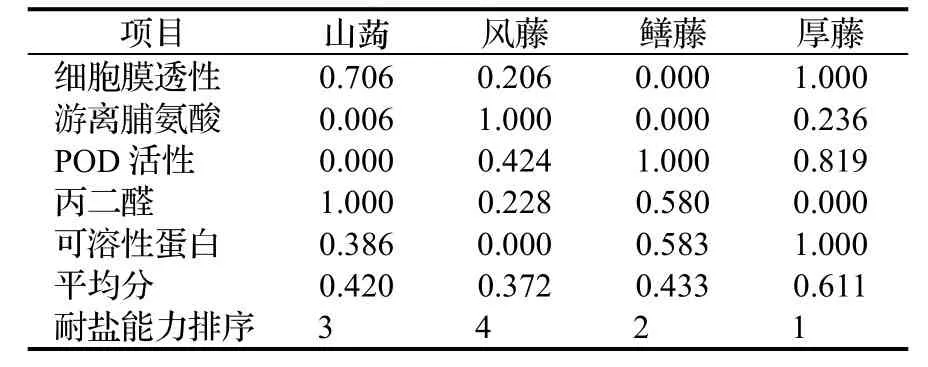
表1 4種藤本植物耐鹽能力綜合評價(隸屬函數法)Table 1 Integrated evaluation of salt-resistance of 4 tested plants (by subordinate function)
3 結論與討論
植物在受到逆境脅迫時,其植株的水分、激素含量等因子會發生一系列的變化,外觀表現為葉片發黃、萎蔫、脫落,嚴重時將致其死亡等特征,具體表現為體內的一些酶發生改變,影響植物體內的同化作用。不同植物在不同的階段有著不同的反應,植物對逆境脅迫下的生理調整會因脅迫方式、強度不同而有所變化,即存在生理調整的多樣性。通過測定其植物體內的激素含量的變化來了解植物體內受迫害程度,從而得知植物體內受逆境脅迫的最大能力,在植物配置時做到適地適栽,為其園林推廣應用提供理論依據。
厚藤在野外主要生長在沙灘上,耐鹽性較強,植株根系極深,可作海灘固沙或覆蓋植物,其四季常綠,葉形奇特,生長勢強,花果期較長,葉花果具有較高的觀賞價值。鱔藤在野外主要生長在巖石邊上,具有一定的耐鹽能力,園林上可借助強大的攀援能力充分覆蓋裸露的山體及墻面等;山蒟和風藤也具有一定的耐鹽性,其四季常綠,枝葉繁茂,葉有辣味不宜感染病蟲害,具氣生根,可攀援與巖石、墻面、樹干、籬笆上生長,其優良的攀援能力可以形成別具特色的立題景觀。
參考文獻:
[1] 唐麗. 論藤本植物的造景功能[J]. 林業調查規劃,2006,2(1):150-153.
[2] 董延龍. 攀緣植物在垂直綠化中的應用[J]. 黑龍江農業科學,2007(2):63-64.
[3] 馬興旺. 干旱區沙漠化土地治理與保護性耕作[J]. 新疆農業科學,2004,41(3):138-142.
[4] 山侖,鄧西平,康紹忠. 我國半干旱地區農業用水現狀及發展方向[J]. 水利學報,2002(9):27-31.
[5] 郭然,王效科,歐陽志云,等. 中國土地沙漠化,水土流失和鹽漬化原因和驅動力總體分析[J]. 自然資源學報,2004,19(l):119-127.
[7] 白玉娥,易津,谷安琳,等. 鹽脅迫對幾種禾本科牧草根系細胞膜透性的影響[J]. 干早地區資源與環境,2001,15(5):51-54.
[8] 孫國榮,閻秀峰. 鹽脅迫對星星草幼苗保護酶系統的影響[J]. 草地學報,2001,9(l):34-38.
[9] 楊傳平,焦喜才,劉文祥,等. 樹木的細胞膜透性與抗鹽性[J]. 東北林業大學學報,1997,25(l):1-3.
[10] Sreenivasulu N,Ramanjulu S,Ramachandra K K, et al. Total peroxidase activity and peroxidase isoforms as modified by salt stress in two
cultivars of fox-tail millet with differential salt tolerance[J]. Plant Sci,1999,141(1):1-9.
[11] 金贊敏,王長海,劉兆普. 鹽脅迫對蘆薈幾項生理生化指標的影響[J]. 食品與發酵工業,2004,30(10):1-4.
[12]張云起,劉世琦,楊鳳娟,等. 耐鹽西瓜砧木篩選及其耐鹽機理的研究[J]. 西北農業學報,2003,12(4):105-108.
[13] 盧靜君,多立安,劉祥君. 鹽脅迫下兩草種SOD和POD及脯氨酸動態研究[J]. 植物研究,2004,24(1):115-119.
[14] Watad A A,Reinhold L,Lerner H R. Comparision between as table NaCl selected nicotianacell lieand wild type[J]. Plant Physiol,1983(73):624-632.
[15] 湯章城. 逆境條件下植物脯氨酸累積及其可能的意義[J]. 植物生理學通訊,1984(l):15-21.
[16]Hanson A D,Nelsen C E,Everson E H. Evalution of free proline ac-cumulation as an index of drought resistance using two contrasting barley cultivars[J]. Crop Sci,1977(17):720-734.
[17] Stewar C R,Lee J A. Therole of proline accumulation ihalophytes[J]. Planta,1974,120(3):279-289.
[18]郭房慶,周建明,湯章城. NaCl脅迫下小麥突變體和野生型葉片中一些有機溶質積累和基因表達差異[J]. 植物生理學報,1999,25(3):263-268.
[19] 徐云嶺,余叔文. 植物鹽脅迫蛋白[J]. 植物生理學通訊,1989(2):12-16.
[20]王愛國,邵從本. 丙二醛作為植物膜質過氧化指標的探討[J]. 植物生理學通訊,1986(2):55-57.
[21] 賈娜爾·阿汗,楊春武,等. 鹽生植物堿地膚對鹽堿脅迫的生理響應特點[J]. 西北植物學報,2007,27(1):79-84.
[22] 高建社,劉玉媛,符毓秦,等. 不同種源青楊幼樹的生長特性[J]. 浙江林學院學報,2004,21(l):11-118.
[23] 蒲光蘭,周蘭英,胡學華,等. 土坡干旱脅迫對杏樹滲透調節物質的影響[J]. 北方園藝,2005(2):50-51.
[24] 楊敏生,裴保華,朱之梯. 水分脅迫下白楊無性系生理和生長的數量遺傳分析[J]. 北京林業大學學報,1997,19(2):50-56.
[25] 鄭書星,樊軍鋒,蘇小華. 歐洲黑楊無性系抗旱性綜合鑒定研究[J]. 西北林學院學報,2005,20(1):57-64.
中圖分類號:S731.2
文獻標識碼:A
文章編號:1001-3776(2011)04-0051-07
收稿日期:2011-02-27;修回日期:2011-04-05
作者簡介:杜月青(1968-),女,浙江紹興人,工程師,從事園林綠化工作;*通訊作者。
Salt Stress on Four Species of Climbing Plants
DU Yue-qing,LIU Jian-Qiang,CHEN Li-Chao,JIN Shui-hu*
(Zhejiang Tiantai Landscaping Co.,Ltd, Shaoxing 312000, China)
Abstract:Experiments were conducted on salt stress on morphology, physiology and biochemistry changes ofAnodendron affine,Piper hancei,P. kadsuraandIpomoea pes-caprae. The results showed that cell membrane permeability of four tested species increased with stress duration and salt concentration. The POD ofP. hancei,P. kadsuraandI. pes-capraeincreased with salt concentration. POD ofA. affinedecreased with stress duration and salt concentration. Free proline content ofP. kadsura,A. affineandI. pes-capraeincreased with stress duration, that ofP. hanceiincreased first and then decreased with stress duration. The soluble protein content of four tested species increased first and then decreased. The MDA content of tested four species increased with stress duration, MDA content topped the highest after twenty days of salt stress.
Key words:salt stress; physiological and biochemical indexs;Anodendron affine;Piper hancei;P. kadsura;Ipomoea pes-caprae
