肉雞和大鼠磷代謝調(diào)控的差異比較研究
2011-06-12 08:00:28方熱軍李美君項(xiàng)智峰
飼料博覽 2011年12期
關(guān)鍵詞:水平
陳 娟,方熱軍,李美君,項(xiàng)智峰
(湖南農(nóng)業(yè)大學(xué)動(dòng)物科技學(xué)院,長(zhǎng)沙 410128)
磷在細(xì)胞生物學(xué)中起關(guān)鍵作用,許多細(xì)胞過程包括核酸合成、代謝、能量代謝、細(xì)胞信號(hào)、膜的完整性、肌肉的功能、酶的活性、脂質(zhì)的代謝和骨的礦化都需要磷或者磷的其他形式。由于磷在動(dòng)物體內(nèi)的重要作用,飼料和飼料添加劑中磷的添加量越來(lái)越多,而單胃動(dòng)物對(duì)植酸磷的利用率很低,大部分植酸磷隨排泄物排出。規(guī)模化畜禽養(yǎng)殖業(yè)中過量磷的排泄所導(dǎo)致的環(huán)境污染,已成為全球關(guān)注的熱點(diǎn)問題。
有學(xué)者對(duì)豬禽糞尿氮磷排放量的分析結(jié)果表明,我國(guó)目前豬禽糞尿氮年排放總量為213.1萬(wàn)t,養(yǎng)殖業(yè)及農(nóng)村畜禽糞尿污染己成為農(nóng)業(yè)面源污染的主要來(lái)源。由此可見,合理添加飼料中磷,減少過量磷排泄導(dǎo)致的生態(tài)環(huán)境污染問題已成為磷在畜牧生產(chǎn)應(yīng)用中丞待解決的難題,提倡低磷耗養(yǎng)殖已成為畜牧業(yè)可持續(xù)發(fā)展的重要方向之一。因此,進(jìn)一步掌握動(dòng)物對(duì)磷吸收的機(jī)制,研究比較不同動(dòng)物磷代謝調(diào)控的差異比較,是進(jìn)行低磷養(yǎng)殖的重要理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)設(shè)計(jì)
試驗(yàn)一,選擇體重約1.5 kg 60日齡的三黃肉雞40只,隨機(jī)分為A、B、C、D 4個(gè)組,4組日糧磷水平依次為0.4%、0.6%、0.8%、1.0%,其中B組為對(duì)照組(所有試驗(yàn)組日糧中鈣∶有效磷均為2∶1),每組5個(gè)重復(fù),每個(gè)重復(fù)2只雞(1個(gè)代謝籠)。試驗(yàn)二,選用120只平均體重為18~22 g,21日齡的雄性昆明大白鼠,隨機(jī)分成A、B、C、D 4個(gè)組,4組日糧總磷水平依次為0.2%、0.4%、0.6%、0.8%,其中C組為對(duì)照組,每組5個(gè)重復(fù),每重復(fù)6只大白鼠。
1.2 試驗(yàn)日糧
三黃肉雞和大鼠試驗(yàn)日糧分別按照“中國(guó)肉雞飼養(yǎng)標(biāo)準(zhǔn)”和“大白鼠的營(yíng)養(yǎng)需要”配制。日糧組成及營(yíng)養(yǎng)水平見表1、2。
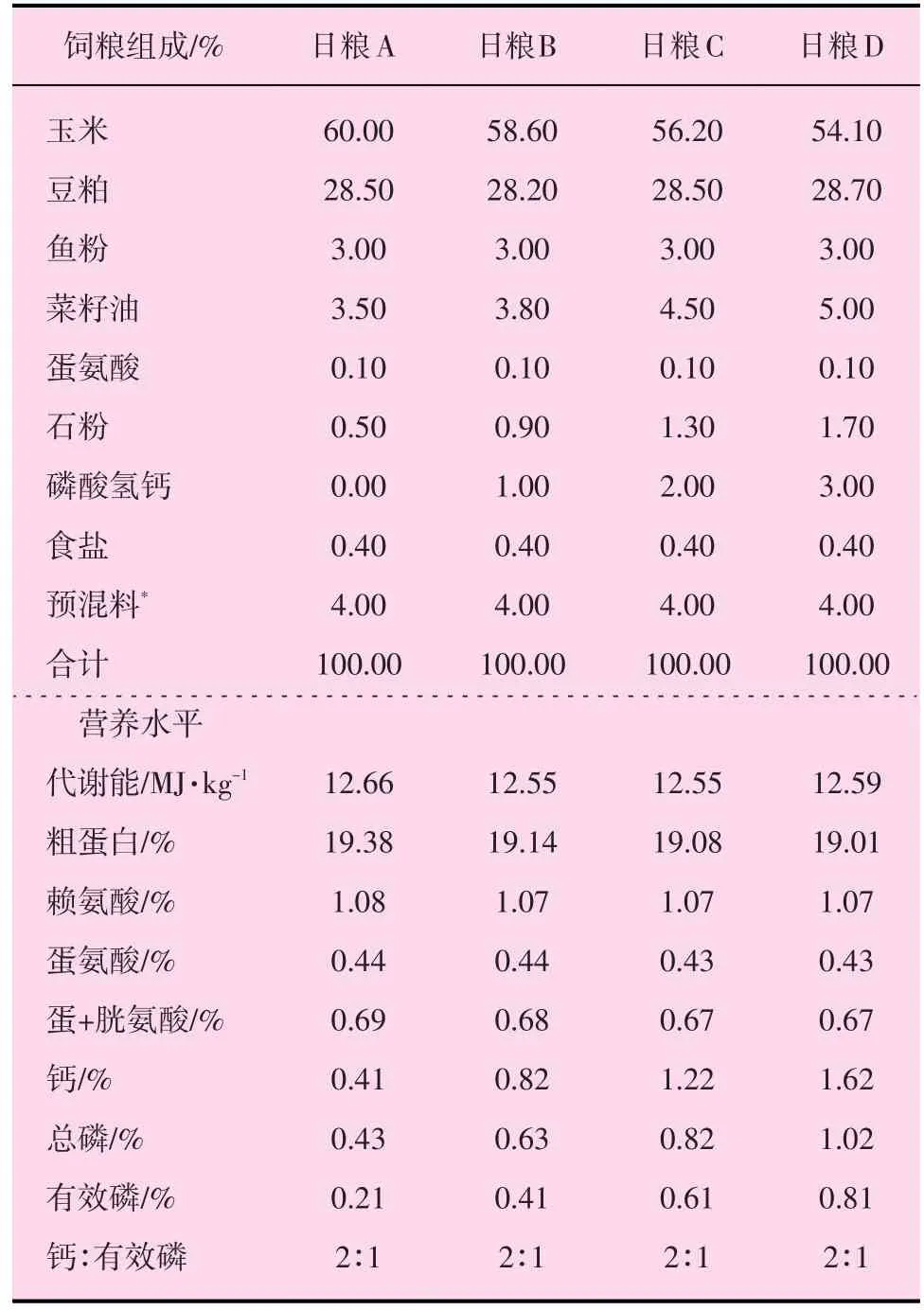
表1 肉雞日糧組成及營(yíng)養(yǎng)水平
1.3 樣品收集與制備
試驗(yàn)設(shè)預(yù)試期3 d和正試期7 d,試驗(yàn)期末屠宰所有試驗(yàn)動(dòng)物,取動(dòng)脈血3000 rpm離心后置血清于-20℃冰箱保存,備測(cè)血鈣和血磷;迅速截取十二指腸、空腸和回腸,將腸道內(nèi)食靡排出后用生理鹽水清洗,經(jīng)液氮處理后分裝包好迅速置于-80℃冰箱,待用。

表2 大白鼠基礎(chǔ)日糧組成及營(yíng)養(yǎng)水平
1.4 血磷和血鈣的測(cè)定
血鈣的測(cè)定采用偶氮胂Ⅲ法;血磷的測(cè)定采用還原鉬藍(lán)法;試驗(yàn)數(shù)據(jù)采用EXCEL初步整理后使用統(tǒng)計(jì)分析軟件SPSS 17.0進(jìn)行處理和方差分析,結(jié)果以“平均值±標(biāo)準(zhǔn)誤”表示。
1.5 小腸各段NaPi-Ⅱb的測(cè)定
RNA的提取按照美國(guó)Invitrogen公司提供的Trizol試劑的使用說明書操作;cDNA第一鏈的合成參照上海生工公司提供的Taq酶使用說明書操作。分別根據(jù)雞GenBank收錄的雞NaPi-Ⅱb基因序列(NM_204474),β-actin基因序列(NM_205518)和已知的大鼠(AF157026)CDS保守序列設(shè)計(jì),使用Primer Premier 5.0進(jìn)行引物設(shè)計(jì),引物合成由上海生物工程公司完成。引物序列為:


反應(yīng)體系為:上下游引物(10μmol·L-1)各 0.3 μL,1×qPCR mix 5 μL,50×ROX 0.2 μL, DNA 溶液0.5 μL,加滅菌蒸餾水至10 μL。循環(huán)反應(yīng)條件為:95℃預(yù)變性60 s,95℃PCR變性15 s,60℃下延伸30 s,退火溫度60℃,40個(gè)循環(huán)。
2 試驗(yàn)結(jié)果
2.1 日糧磷水平對(duì)血磷和血鈣的影響
日糧磷水平對(duì)血磷和血鈣的影響見表3。

表3 日糧磷水平對(duì)血磷和血鈣的影響mg·L-1
由表3可知,隨著日糧總磷水平的提高,雞和大鼠血清磷的含量都呈上升趨勢(shì),在0.4%時(shí)血清磷最低,1.0%時(shí)最高,且兩者0.4%與0.6%、0.8%和1.0%磷水平之間皆差異極顯著(P<0.01)。試驗(yàn)雞血清鈣含量隨日糧總磷水平的提高而下降,在4個(gè)磷水平間差異極顯著(P<0.01)。試驗(yàn)大鼠血鈣隨日糧總磷水平的提高而升高,但各組間差異均不顯著(P>0.05)。
2.2 日糧磷水平對(duì)腸段NaPi-Ⅱb mRNA表達(dá)的影響
日糧磷水平對(duì)不同腸段NaPi-Ⅱb mRNA表達(dá)量的影響見附圖。


附圖 磷水平對(duì)不同腸段NaPi-Ⅱb mRNA表達(dá)量的影響
本試驗(yàn)通過Real-time PCR研究顯示,不同磷水平顯著影響大鼠和肉雞各腸段中NaPi-Ⅱb mRNA相對(duì)表達(dá)量(P<0.05)。大鼠日糧磷含量0.2%水平組與正常磷水平組比較,回腸、空腸、十二指腸NaPi-Ⅱb mRNA表達(dá)量最高,隨著日糧磷水平的提高,空腸和十二指腸NaPi-Ⅱb mRNA表達(dá)量明顯降低,回腸NaPi-Ⅱb mRNA表達(dá)量當(dāng)日糧磷含量從0.2%升至0.4%時(shí)顯著降低,但是當(dāng)日糧磷含量從0.4%升至0.8%時(shí),NaPi-Ⅱb mRNA表達(dá)量無(wú)顯著變化。此結(jié)果與對(duì)大鼠、小鼠和虹鱒研究結(jié)果相一致,即高磷飼糧降低腸道NaPi-Ⅱb mRNA表達(dá)量[1-3]。在肉雞試驗(yàn)中,空腸中NaPi-Ⅱb mRNA,在磷含量0.4%水平日糧時(shí)組中表達(dá)最高達(dá)1.88,是磷含量1.0%水平日糧組的427%,但是在回腸和十二指腸內(nèi)未出現(xiàn)顯著升高趨勢(shì),只是在1.0%水平日糧組中,回腸和十二指腸內(nèi)NaPi-Ⅱb mRNA的表達(dá)量最低。可見低磷提高了小腸中NaPi-Ⅱb mRNA的表達(dá),高磷抑制小腸中NaPi-Ⅱb mRNA的表達(dá);不同磷水平對(duì)不同動(dòng)物不同的組織部位中NaPi-Ⅱb mRNA影響不同,大鼠和肉雞空腸均隨著日糧磷水平的提高NaPi-Ⅱb mRNA表達(dá)量降低,回腸和十二指腸內(nèi)未出現(xiàn)顯著降低趨勢(shì)。
3 討論
3.1 不同磷水平日糧對(duì)動(dòng)物血磷的影響
隨著日糧磷水平的減少,會(huì)引起機(jī)體內(nèi)磷離子的一系列變化,機(jī)體組織內(nèi)磷離子的吸收和保留要加強(qiáng),相反磷的排出就會(huì)減少;當(dāng)日糧中磷水平升高時(shí),機(jī)體內(nèi)磷離子的反應(yīng)相反。細(xì)胞或者多細(xì)胞器官可以通過特殊的磷感受體感受到細(xì)胞外磷離子的變化[4]。反過來(lái)這種感受體通過改變細(xì)胞內(nèi)蛋白的磷酰基位置來(lái)改變?cè)摰鞍追磻?yīng)機(jī)制,受基因控制的蛋白質(zhì)的合成加速了細(xì)胞和細(xì)胞磷感受體組成。
血磷的濃度影響FGF-23、sFRP-4、IGF-Ⅰ等影響磷吸收因子的水平,而這些因子的水平又調(diào)節(jié)腎臟的重吸收。不過關(guān)于這種說法,目前還存在爭(zhēng)議[5]。飼喂低磷日糧動(dòng)物體內(nèi)血磷的降低會(huì)引起血鈣濃度的升高,而血鈣濃度的升高會(huì)限制PTH的釋放,從而減少腎臟內(nèi)磷離子的排出。磷離子的濃度可能會(huì)直接影響到PTH的釋放。低磷日糧或血磷濃度的降低刺激1,25(OH)2D3的合成,1,25(OH)2D3的合成與PTH的無(wú)關(guān)。相反當(dāng)飼喂高磷日糧時(shí)血鈣濃度降低,PTH的濃度升高。由于磷離子對(duì)細(xì)胞功能的重要性,一系列機(jī)制參與調(diào)節(jié)磷離子的平衡。飼喂低磷日糧動(dòng)物排泄物中磷的含量會(huì)減少,這是對(duì)磷缺少以及PTH少量的一種調(diào)節(jié)[6]。有試驗(yàn)研究發(fā)現(xiàn),日糧磷水平0.4%與0.6%、0.8%相比對(duì)野豬血清鈣水平影響差異極顯著(P<0.01);日糧磷水平0.6%與0.8%對(duì)血清鈣水平影響差異不顯著(P>0.05)。日糧磷水平在0.4%時(shí)血清磷最低,0.8%時(shí)最高,并且磷水平0.8%與磷水平0.6%日糧血清磷都極顯著地高于0.4%磷水平日糧(P<0.01)。Cromwell等和Coalson等的研究報(bào)道也同樣證實(shí)了這一試驗(yàn)結(jié)果[7-8]。
Huber等試驗(yàn)發(fā)現(xiàn),從低磷日糧到高齡日糧肉雞血磷濃度隨著日糧磷的濃度升高而上升,最高達(dá)到1 mmol·L-1,低磷日糧組肉雞血鈣濃度顯著高于對(duì)照組和高磷日糧組,另外,小腸食靡內(nèi)磷離子濃度隨日糧磷濃度增加而上升,鈣離子與此相反,在空腸內(nèi)鈉磷轉(zhuǎn)運(yùn)體容積,NaPi-Ⅱb mRNA表達(dá),血磷濃度和磷的排出彼此不相關(guān)[9]。因此,肉雞空腸內(nèi)磷的吸收和NaPi-Ⅱb的表達(dá)是不受日糧磷濃度調(diào)控的。目前關(guān)于空腸內(nèi)NaPi-Ⅱb mRNA表達(dá)量隨日糧磷含量增加上升的機(jī)制還不清楚。
3.2 不同日糧磷水平對(duì)NaPi-Ⅱb mRNA表達(dá)的影響
小腸是磷吸收的主要部位。早期研究表明,小腸內(nèi)上皮細(xì)胞對(duì)磷的吸收是依賴于鈉離子。小腸內(nèi)磷吸收載體主要是NaPi-Ⅱb,其已經(jīng)在大鼠和人體內(nèi)克隆。有研究發(fā)現(xiàn)大鼠十二指腸和空腸內(nèi)磷轉(zhuǎn)運(yùn)體的活性要高于回腸,但是回腸內(nèi)NaPi-Ⅱb蛋白的表達(dá)顯著高于十二指腸,而十二指腸BBMV上鈉離子依賴式磷轉(zhuǎn)運(yùn)的活性卻比回腸高。低磷日糧飼喂的大鼠空腸BBMV上鈉磷轉(zhuǎn)運(yùn)蛋白的活性比標(biāo)準(zhǔn)日糧或者高磷日糧高67%,但是在十二指腸內(nèi)沒有顯著變化。Huber等研究發(fā)現(xiàn),未斷奶前的小山羊空腸內(nèi)鈉離子依賴式磷轉(zhuǎn)運(yùn)體可以根據(jù)體內(nèi)磷離子的平衡調(diào)節(jié)磷的排出[9]。雖然高磷日糧可以抑制鈉離子依賴式磷轉(zhuǎn)運(yùn)體的數(shù)量和活性,但是低磷日糧并不能促進(jìn)鈉離子依賴式磷轉(zhuǎn)運(yùn)體的數(shù)量和活性,十二指腸內(nèi)磷的轉(zhuǎn)運(yùn)體是不屬于鈉離子依賴式磷轉(zhuǎn)運(yùn)體,其結(jié)構(gòu)及性質(zhì)還有待研究。空腸內(nèi)磷的吸收機(jī)制是骨化三醇結(jié)合細(xì)胞核受體后引起的基因機(jī)制[10]。和單胃動(dòng)物一樣,山羊空腸內(nèi)磷的吸收主要是由小腸刷狀緣膜上的鈉離子依賴式磷轉(zhuǎn)運(yùn)體的數(shù)量決定的低磷日糧同樣會(huì)促進(jìn)山羊空腸內(nèi)Ⅱ型鈉離子依賴式磷轉(zhuǎn)運(yùn)體數(shù)量[9]。但是與單胃動(dòng)物不同的是,低磷日糧促進(jìn)空腸內(nèi)鈉離子依賴式磷轉(zhuǎn)運(yùn)體的活性的同時(shí)不改變血液骨化三醇的濃度。另外,鈉離子依賴式磷轉(zhuǎn)運(yùn)體的最大結(jié)合常數(shù)和解離常數(shù)等性質(zhì)不發(fā)生改變[11]。同樣,高磷日磷雖然會(huì)導(dǎo)致PTH目標(biāo)分子鈉離子依賴式磷轉(zhuǎn)運(yùn)體的減少,但是并不會(huì)引起反芻動(dòng)物PTH濃度的變化[12]。
Murer等研究發(fā)現(xiàn),對(duì)空腸和腎臟鈉磷轉(zhuǎn)運(yùn)體的生理調(diào)控主要是通過控制轉(zhuǎn)運(yùn)體的數(shù)量[13]。由NaPi-Ⅱb控制的鈉離子依賴式磷轉(zhuǎn)運(yùn)體具有較低的Km值,表現(xiàn)出較好的親水性。在雞空腸內(nèi)鈉磷轉(zhuǎn)運(yùn)體的Km值大概是40 mol·L-1,與其他哺乳動(dòng)物相似。在雞腎臟內(nèi)由NaPi-Ⅱa調(diào)節(jié)的鈉離子依賴式磷轉(zhuǎn)運(yùn)體的Km值為100 mol·L-1。同哺乳動(dòng)物一樣,雞體內(nèi)磷的平衡取決于NaPi-Ⅱ的表達(dá)以及鈉離子依賴式磷轉(zhuǎn)運(yùn)體的活性,但是Vmax和NaPi-ⅡmRNA的表達(dá)并未受日糧磷水平的限制而明顯變化。食靡和血漿磷濃度都沒有影響到小腸內(nèi)磷的吸收。高磷日糧會(huì)引起血磷濃度降低而血鈣濃度升高。血漿磷離子濃度的升高必然會(huì)引起磷排出的升高,這也是降低小腸或者腎臟對(duì)磷吸收的主要原因。隨著血漿磷離子濃度的升高,腎臟內(nèi)鈉離子依賴式磷轉(zhuǎn)運(yùn)體Vmax以及NaPi-Ⅱa mRNA的表達(dá)顯著下降,這也就降低了腎臟對(duì)磷的重吸收功能,最后導(dǎo)致排泄物中磷含量增加。但是小雞空腸內(nèi)Vmax和NaPi-Ⅱb mRNA的表達(dá)量并未隨血漿磷離子的濃度有所變化。日糧中總磷的增加會(huì)導(dǎo)致磷的排出增加。相關(guān)數(shù)據(jù)表明腎臟內(nèi)鈉磷轉(zhuǎn)運(yùn)體以及NaPi-Ⅱa mRNA會(huì)隨血磷濃度降低從而導(dǎo)致磷排出量增加,但是空腸內(nèi)鈉磷轉(zhuǎn)運(yùn)體以及NaPi-Ⅱb mRNA表達(dá)量并未隨血磷濃度有所變化。
4 結(jié)論
日糧中磷的水平影響NaPi-Ⅱb載體蛋白mRNA表達(dá)和磷的攝入量,當(dāng)飼糧磷水平較低時(shí),NaPi-Ⅱb mRNA表達(dá)量隨飼糧磷水平升高而有所增加;但是當(dāng)飼糧磷水平升高到一定程度時(shí),會(huì)抑制NaPi-Ⅱb mRNA表達(dá);NaPi-Ⅱb載體蛋白mRNA表達(dá)量在不同動(dòng)物不同部位有所不同,在大鼠小腸不同腸段中磷吸收高低依次為回腸、空腸、十二指腸,在肉雞不同腸段中磷吸收高低依次為十二指腸、空腸、回腸,日糧添加總磷水平0.4%有利于提高磷利用率。
[1]Katai K,Miyamoto K,Kishida S,et al.Regulation of intestinal Na+-dependen phosphate co-transporters by a low-phosphate di?et and 1,25-dihydroxyvitamin D3[J].Biochem,1999,343:705-712.
[2]Segawa H,Kaneko I,Yamanaka S,et al.Intestinal Na-P(i)co?transporter adaptation to dietary P(i)content in vitamin D receptor null mice[J].American Journal Physiol Renal Physiol,2004,287:39-47.
[3]Sugiura S H,Ferraris N K,Ferraris R P.In vivo fractional Pi ab?sorption and Na/Pi-ⅡmRNA expression in rainbow trout are up?regulated by dietary P restriction[J].Ameircan Journal of Physiol?ogy Regulatory,Interative and Comparative Physiology,2003,285:770-781.
[4]Lamarche M G,Wanner B L,Crepin S.et al.The phosphate regu?lon and bacterial virulence:a regulatory network connecting phos?phate homeostasis and pathogenesis[J].FEMS Microbiol Reviews,2008,32(3):461-473.
[5]Burnett S A M,Gunawardene S C,Bringhurst F R,et al.Regula?tion of cterminaland intact FGF-23 by dietary phosphatein men and women[J].Journal of Bone and mineral Research,2006,21(8):1187-1196.
[6]Steele T H,Underwood J L.Renal response to phosphorus depriva?tion in the isolated rat kidney[J].Laboratory Investigation Kidney International,1978,13:124-128.
[7]Cromwell G L,Hays V W,Chaney C H,et al.Effects of dietary phosphorus and calcium level on perfor-mance,bone mineraliza?tion and carcass characteristics of swine[J].Journal of Animal Sci?ence,1970,30:519-525.
[8]Coalson J A,Maxwell C V,Hillier J C,et al.Calcium and phospho?rus requirements of young pigs reared under controlled environ?mental conditions[J].Journal of Aminal Science,1972,35:1194-1200.
[9]Huber K C,Walter B,Schroder J.et al.Epithelial phosphate trans?porters in small ruminants[J].Annals of the New York Academy of Science,2000,915:95-102.
[10]Hattenhauer O,Traebert M,Murer H,et al.Regulation of small in?testinal Na-P(i)typeⅡb cotransporter by dietary phosphate in?take[J].American Journal of Physidogy Gastrointestional and Liv?er Physiology,1999,277:756-762.
[11]Schroder B,Kappner H,Failing K,et al.Mechanisms of intestinal phosphate transport in small ruminants[J].British Journal of Nutri?tion,1995,74:635-648.
[12]Wan Z M,Scott D,Loveridge N,et al.The effect of high phospho?rus intake on calcium and phosphorus retention and bone turnover in growing lambs[J].Expenimental Physiology,1994,79:175-181.
[13]Murer H,Biber J.A molecular view of proximal tubular inorganic phosphate(Pi)reabsorption and of its regu1ation[J].Pflugers Ar?chiv European Journal of Physiology,1997,433:379-389.
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國(guó)火炬(2010年12期)2010-07-25 13:26:22
中國(guó)火炬(2010年8期)2010-07-25 11:34:30
