培養(yǎng)液添加物與去核方法對豬體細胞核移植效率的影響
2011-07-12 05:11:20張林波田見暉
黑龍江動物繁殖 2011年4期
關(guān)鍵詞:效率
張 歆 ,張林波,田見暉*
(1.吉林農(nóng)業(yè)大學 生命科學學院生物制藥創(chuàng)新實驗室,吉林 長春 130118;2.中國農(nóng)業(yè)大學 動物科學技術(shù)學院/農(nóng)業(yè)部動物遺傳育種與繁殖重點實驗室,北京 100193)
體細胞核移植(SCNT),又稱體細胞克隆。作為動物細胞工程技術(shù)的常用手段,即把體細胞移入去核卵母細胞中,使其重組并能發(fā)育成新的胚胎,最終發(fā)育成個體。其過程主要包括:卵母細胞體外成熟即發(fā)育至第2次減數(shù)分裂中期(MⅡ期),脫去顆粒細胞后將供體細胞注入去核的卵母細胞內(nèi),經(jīng)過電融合、化學激活和后期培養(yǎng)發(fā)育到相應(yīng)的時期后移植到受體內(nèi)直至妊娠。進行豬的克隆研究最重要的意義在于進行異種器官移植,生產(chǎn)無免疫排斥反應(yīng)的轉(zhuǎn)基因豬,將其器官應(yīng)用于人類臨床醫(yī)學的相關(guān)研究,因而具有巨大的社會和經(jīng)濟價值。生產(chǎn)轉(zhuǎn)基因豬首先在體外培養(yǎng)的體細胞中進行基因?qū)耄Y選穩(wěn)定整合外源基因的陽性轉(zhuǎn)基因細胞,然后將轉(zhuǎn)基因體細胞核移植到去掉細胞核的卵細胞中生產(chǎn)重構(gòu)胚胎,經(jīng)移植到母體中發(fā)育產(chǎn)生仔畜[1]。盡管有很多體細胞核移植的克隆豬出生,也做了許多技術(shù)上的改進,但是總的效率還是很低。影響轉(zhuǎn)基因克隆效率的因素很多,包括卵母細胞的成熟液成分及培養(yǎng)時間,去核和融合方法,供體細胞的類型、周期,及供體核進入卵母細胞后的重編程過程等。近些年來許多研究者都致力于對提高克隆效率的研究。本文將從三方面對克隆效率的影響及近年來在這幾方面的成果進行綜述,即在卵母細胞的成熟、去核和后期發(fā)育過程中添加化學成分對體細胞核移植效率的影響。
1 在成熟液中添加化學成分對SCNT的作用
卵母細胞的成熟是胚胎體外生產(chǎn)體系的一個非常關(guān)鍵的步驟,它影響著卵母細胞的發(fā)育能力和質(zhì)量,進而直接影響胚胎的發(fā)育,甚至是后代幼仔的健康[2]。和體內(nèi)的成熟相比,體外發(fā)育的能力還是很低的,因此,提高SCNT的效率必須對成熟培養(yǎng)液進行優(yōu)化。
1.1 成熟液體系在各國家的應(yīng)用情況
關(guān)于卵母細胞的成熟很多人用的是兩步成熟過程,第1步是卵丘卵母細胞在含有激素的成熟液中培養(yǎng)20~22 h,比如添加eCG和hCG來促進核成熟;第2步使卵丘卵母細胞在不含有激素的成熟液中培養(yǎng)18~24 h,移除激素是為了減慢核成熟提高細胞質(zhì)的成熟。比較了卵母細胞的成熟體系之后,發(fā)現(xiàn)有8個實驗室都在用北卡羅來納州大學的培養(yǎng)液(NCSU-23),添加10%的豬卵泡液作為基本的成熟培養(yǎng)液,有4個實驗室用NCSU37+10%豬卵泡液,5個利用TCM199+10%PFF,有1個用TCM199添加0.1%PVA。這些基礎(chǔ)培養(yǎng)液還另外添加了許多不同成分,如半胱氨酸、EGF、dbcAMP、胰島素和氨基酸等[3]。
1.2 成熟液中添加leptin等物質(zhì)對SCNT的影響
體外成熟的卵母細胞受諸多因素的影響,包括成熟時間、溫度、相關(guān)的細胞因子等[4]。在這些因素中,補充細胞因子在卵母細胞核質(zhì)成熟中起著重要作用[5]。研究者發(fā)現(xiàn),在小鼠的卵母細胞卵泡發(fā)育的各個階段和卵母細胞成熟過程中都發(fā)現(xiàn)有l(wèi)eptin的高表達,推測可能與卵母細胞的成熟有關(guān)。向豬卵母細胞的成熟液中添加leptin,最終發(fā)現(xiàn)可以提高孤雌發(fā)育和SCNT胚胎的發(fā)育能力,而且具有劑量依賴性。研究還發(fā)現(xiàn),leptin是通過MAPK通路來促進核與質(zhì)的成熟[6]。實驗結(jié)果表明,核移植的顯微操作過程中,leptin處理組和對照組相比,在融合和激活之后卵母細胞的死亡率都會降低[7]。在最近一項實驗中提到,銨可以影響核和質(zhì)的成熟,尤其是谷氨酰胺,能夠產(chǎn)生銨離子,對細胞產(chǎn)生毒害作用。若將IVM液中的谷氨酰胺換成丙酰胺基谷酰胺或者甘氨酰基谷酰胺就可以減少銨的累積,保護卵母細胞不受銨的破壞,順利完成減數(shù)分裂和后期的發(fā)育[8]。據(jù)Guo-Shi Liu報道,褪黑素在卵泡液中的濃度大約是10-11mol/L,這就說明褪黑素在卵母細胞成熟過程中起著重要作用。作者在成熟培養(yǎng)液中添加不同濃度褪黑素,結(jié)果囊胚率和囊胚細胞數(shù)均高于對照組[9]。而在另外一項報道中也有類似的結(jié)果,只是細胞數(shù)一項沒有提高[10]。諸多實驗結(jié)果表明,褪黑素是一種直接作用的抗氧化劑,具有強烈清除活性氧的作用[11],有助于提高SCNT效率。
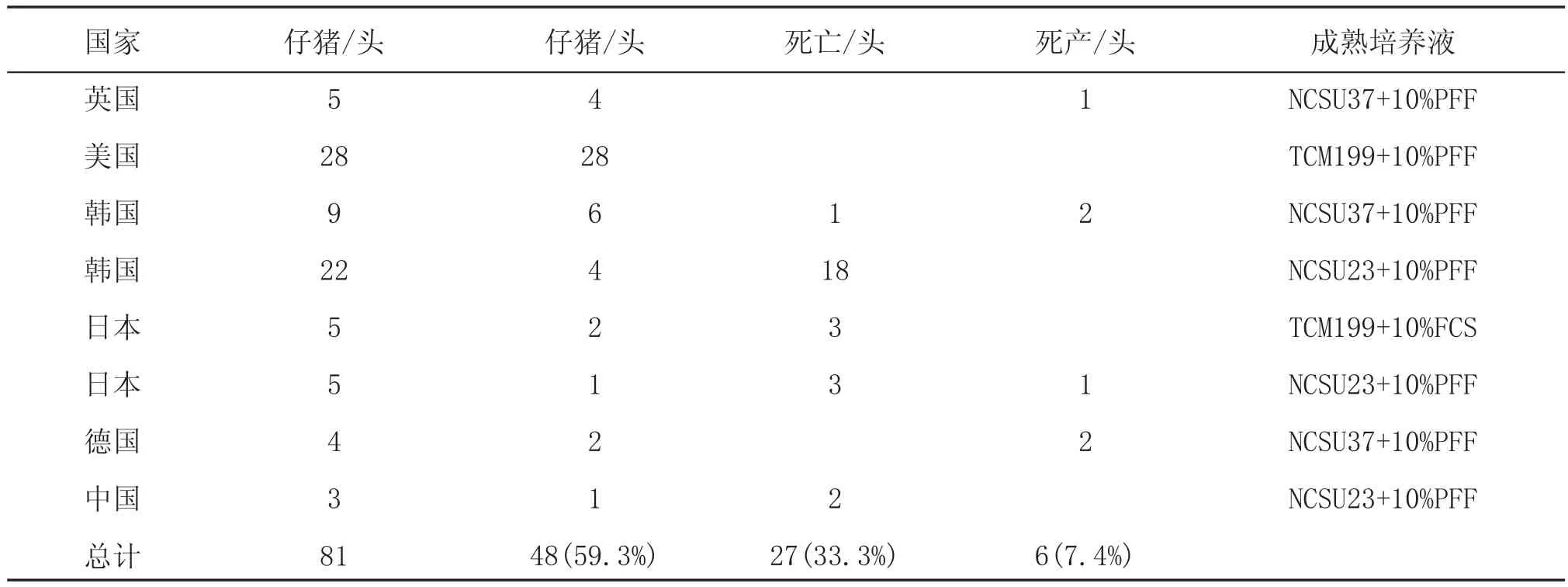
表1 各國家之間生產(chǎn)非轉(zhuǎn)基因豬其成熟培養(yǎng)液使用情況比較
2 去核方法對SCNT效率的影響
2.1 物理去核
利用胞質(zhì)體作為核移植的受體首先就要清除卵母細胞內(nèi)的遺傳物質(zhì),這樣就必須先確定卵母細胞核的位置。通常所用的方法是盲吸法,就是以第1極體作為標記來確定MⅡ期卵母細胞紡錘體的位置,吸掉整個極體和極體下面很少的一部分胞質(zhì)。但是許多情況下赤道板和第1極體不在相鄰的位置上[12],這種現(xiàn)象在牛和兔上也有報道。實驗指出只有50%的紡錘體存在于第1極體附近,甚至更少[13]。為了提高去核率,第1極體下面有時有1/3的胞質(zhì)被吸走,這樣大量地去除胞質(zhì)可能會導致轉(zhuǎn)基因核重排能力的下降甚至發(fā)育不完全。而且,若在卵母細胞成熟之前即脫掉顆粒細胞,這樣會破壞第1極體和MⅡ期紡錘體的相互作用,導致在去核之后仍有遺傳物質(zhì)的殘留[14]。還有許多物理去核方法,諸如在卵母細胞成熟的不同時期進行去核等[15-17],對后期發(fā)育都存在不同程度的影響。
2.2 化學去核
化學去核就是使用化學藥物進行處理來輔助移除卵母細胞的染色體。利用細胞骨架修飾因子,如細胞松弛素B、秋水仙胺和nocodazole可以改變重構(gòu)胚胎中DNA內(nèi)含物的結(jié)構(gòu)和功能。秋水仙素是一種微管解聚劑,可以有效地誘導卵母細胞的去核過程。而且,在激活之后用秋水仙素處理可以使豬的轉(zhuǎn)基因胚胎完整地發(fā)育,誘導單個的前核形成并提高DNA的倍性[18]。和秋水仙素相似,nocodazole可以成功地進行輔助去核過程,它能夠穩(wěn)定中期赤道板,產(chǎn)生1個小的突起,使染色質(zhì)更為明顯,容易去除[19]。nocodazole的去核作用使人們開始嘗試利用化學輔助去核方法生產(chǎn)轉(zhuǎn)基因克隆豬。在研究的早期有人應(yīng)用etoposide,或者etoposide和放線菌酮的復合體,或者乙醇和秋水仙胺。但是此種化學處理方法和常規(guī)的去核方法相比,卵裂率和后期發(fā)育率均很低[20]。
2.3 其他的去核方法
用3%的蔗糖處理小鼠的卵母細胞會使其在光學顯微鏡下出現(xiàn)更多的半透明部分,更容易觀察減數(shù)分裂的紡錘體形態(tài)[21]。還有人報道說,POL-scope系統(tǒng)可以使紡錘體的定位觀察更加直觀,但是這種儀器非常昂貴,在不同的品種之間還需要多重復合[22]。如果把牛的卵母細胞離心然后結(jié)合CB的處理,去核后也有發(fā)育能力,但是這種方法必須移除透明帶,這可能會阻礙到后期的進一步發(fā)育[23]。
3 在胚胎發(fā)育液中添加物質(zhì)對SCNT效率的影響
3.1 CSF2
有一系列的論據(jù)可以證明,CSF2在早期胚胎發(fā)育過程中作為一個生理學重要的調(diào)節(jié)器而發(fā)揮作用。在腔上皮組織和輸卵管以及子宮內(nèi)膜組織中有表達[24,25]。CSF2在胚胎發(fā)育過程中作為一個重要的調(diào)節(jié)因子可以提高克隆胚胎的耐受性,在移植之后可以提高囊胚率和妊娠率。其原因一方面可能是CSF2可以促進有絲分裂,增加胚胎的細胞數(shù),充分地促進囊胚腔的形成。另一方面,CSF2通過阻斷細胞凋亡來增加細胞數(shù)。通過觀察,CSF2處理的卵母細胞有相當數(shù)量的與外滋養(yǎng)層有關(guān)的內(nèi)細胞團,表明CSF2影響著囊胚的分化[26]。此結(jié)論在人和小鼠上都有過驗證。
3.2 TSA(組蛋白脫乙酰基酶抑制劑)
目前認為:供體核的不完全重編程是導致克隆效率低的主要原因。關(guān)于供體核的重編程,其研究主要集中在DNA甲基化和組蛋白乙酰化。組蛋白的乙酰化發(fā)生在核心組蛋白的賴氨酸殘基上[27]。核心組蛋白在MⅠ期高度乙酰化,然后立刻去乙酰化,在MⅡ期大多數(shù)的核心組蛋白賴氨酸位點都去乙酰化,除了H4K8位點[28]。體細胞基因組中核心組蛋白的乙酰化有兩種,第1種發(fā)生在組蛋白H3K4的3個賴氨酸位點上,在體細胞核移植之后立刻發(fā)生去乙酰化,3 h后完全去乙酰化。卵母細胞激活之后在這些位點上發(fā)生再次乙酰化。在SCNT過程中這種組蛋白賴氨酸位點的去乙酰化和重新乙酰化過程對克隆胚胎的基因表達和全能性的建立是非常關(guān)鍵的,也直接影響著克隆效率。
最近的一項研究表明,提高供體細胞或者克隆胚胎中組蛋白的乙酰化水平可以促進發(fā)育從而提高效率[29]。用一種組蛋白脫乙酰基酶抑制劑處理牛的胎兒成纖維細胞可以提高克隆胚胎的發(fā)育[30]。另一種組蛋白脫乙酰基酶抑制劑TSA,可以提高克隆效率,在牛和小鼠上都已經(jīng)有過驗證[31,32]。在2009年,也有研究表示,TSA處理可以提高克隆胚胎基因組的乙酰化水平,在很多物種中都提高了后期發(fā)育能力[33]。而且TSA處理的兔SCNT胚胎的組蛋白乙酰化程度比未處理組要高。這種現(xiàn)象在TSA處理的小鼠SCNT胚胎中也得到了驗證[32]。在豬的手工克隆中也有TSA的應(yīng)用,而且可以很顯著地提高發(fā)育能力,并順利完成整個發(fā)育過程[34]。但是在同年,據(jù)Meng報道,用TSA處理兔子的SCNT胚胎,限制了其后期發(fā)育的能力,沒有提高囊胚率,處理和未處理組均有后代產(chǎn)生,但是TSA處理組沒有發(fā)育到成年[29]。可能的原因就是,在不同的物種中體細胞核移植操作的差異導致了不同的結(jié)果。在現(xiàn)在的研究中,人們試圖篩選TSA處理合適的時間及濃度,這就需要正確的SCNT操作方法,TSA適當?shù)奶幚硪约疤禺惖奈锓N之間成功的結(jié)合才能夠最終達到提高轉(zhuǎn)基因效率的目的。
3.3 褪黑素
機體在有氧代謝過程中產(chǎn)生的活性氧族(ROS)對大多數(shù)細胞都具有毒性作用。當ROS產(chǎn)生增多或者機體清除能力下降時機體就會出現(xiàn)氧化應(yīng)激。為了阻止機體產(chǎn)生氧化應(yīng)激,在培養(yǎng)介質(zhì)中添加抗氧化劑或者自由基清除劑,結(jié)果發(fā)現(xiàn)可以有效地提高胚胎的發(fā)育能力。褪黑素,又名N-乙酰-5-氧基色胺,是有效的自由基清除劑和抗氧化劑[35]。和其他已知的自由基清除劑相比,褪黑素具有多功能性而且比較普遍。它作為一種既疏水又親水的復合溶液,可以溶解在水中和類脂中,說明褪黑素和它的代謝產(chǎn)物可以直接清除羥基自由基、烴氧基、過氧化氫根、亞硝酸鹽陰離子、氧化亞氮和單獨的氧,而且,褪黑素還能夠修復被氧化的分子,抑制脂質(zhì)過氧化反應(yīng)等[36]。有報道說,褪黑素還可以提高小鼠的囊胚發(fā)育率[37]。通過毒性試驗驗證,在小鼠和兔子的胚胎發(fā)育上無論在體內(nèi)還是體外組均無毒害作用。據(jù)Yuichi ro Kitagawa報道,在低氧條件下,豬卵母細胞體外發(fā)育過程中可以減少過氧化氫含量,從而減少了DNA斷裂的發(fā)生,提高了胚胎的發(fā)育能力[38]。Krzysztof Papis等人還做了進一步的研究,發(fā)現(xiàn)在20%高氧條件下添加褪黑素可以促進發(fā)育,在7%低氧條件下反而會抑制發(fā)育,這是否說明褪黑素在低氧條件下產(chǎn)生了副作用,或者影響了乙酰化過程還需要進一步驗證[39]。
3.4 nocodazole
nocodazole是一種細胞骨架修飾因子。若將細胞支架的修飾因子注入到SCNT的卵母細胞中,可以影響核的重塑,抑制由于阻斷微管的解聚而導致的同源染色體的丟失,提高DNA的倍性和SCNT的發(fā)育。核的重排和保持核的倍性是轉(zhuǎn)基因胚胎發(fā)育的必要條件。在目前國內(nèi)的研究中,已經(jīng)驗證了nocodazole的作用,它是一種細胞骨架修飾因子,可改變供體核的形態(tài),引入已經(jīng)去核的卵母細胞中從而影響豬轉(zhuǎn)基因胚胎的后期發(fā)育。結(jié)果顯示,卵母細胞激活之后用nocodazole處理可以提高單核的形成率,通過排出極體來抑制DNA的丟失,從而保證了核的倍性。nocodazole處理可以使轉(zhuǎn)基因胚胎完成完整的發(fā)育過程,但是幼仔的形成率一項和對照組相比沒有明顯的差異[40]。在先前許多研究中都發(fā)現(xiàn),無論是孤雌胚胎還是轉(zhuǎn)基因胚胎中,細胞骨架修飾因子如秋水仙胺和細胞松弛素B都可以保留核的遺傳信息,這很可能影響了微管的組裝[41]。
4 結(jié)語
在卵母細胞成熟、去核以及后期發(fā)育過程中添加化學物質(zhì),均不同程度地提高了核移植的效率,但傳統(tǒng)的添加實驗基于經(jīng)驗式的摸索,具有盲目性和不可重復性,而且效率低下。近些年研究表明,體細胞核重編程的不完全是克隆胚胎發(fā)育效率低下的重要原因,雖然已有個別實驗室通過添加組蛋白去乙酰化酶抑制劑等藥物來提高克隆胚胎的發(fā)育能力,但進展緩慢。因此,采用表觀遺傳學研究的相關(guān)手段,解析克隆胚胎發(fā)育過程中體細胞核的重編程過程將是未來克隆技術(shù)研究的一個重要方面。此類研究對于改進體細胞核移植技術(shù),保證克隆胚胎的安全性具有重大的意義。
[1] 張運海,潘登科,孫秀柱,等.利用體細胞核移植技術(shù)生產(chǎn)表達綠色熒光蛋白的豬轉(zhuǎn)基因克隆胚胎.中國科學C輯生命科學,2005,35(5): 439-445.
[2] Eppig JJ,O’Brien MJ.Comparison and preimplantation developmental competence after mouse oocyte growth and development in vitro and in vivo.Theriogenology,1998,49(2), 415–422.
[3] P Fisher,W C Chen ,I ChoiR,et al.Somatic cellnuc leartrans fer: Past, present and future perspectives.Theriogenology,2007,68S (S214–S231).
[4] Nagai T.The improvement of in vitro maturation systems for bovine and por c ine oocy tes.Theriogenology 2001,55,1291–1301.
[5] Coy P, Romar R, Ruiz S, et al.Birth of piglets after transferring of in vitro-produced embryospre-matured with R-roscovitine.Reproduction,2005,129(6): 747–755.
[6] Craig JA,Zhu H,Dyce PW, et al.Leptin enhances porcine preimplantation embryo development in vitro.Mol Cell Endoc rinol,2005,229(1-2):141–147.
[7] Zhang Kun,Li Ning.Ef fects of leptin supplementa-tion in in vitro maturation medium on meiotic maturation of oocytes and preimp lantation development of par thenogenetic and cloned embryos in pigs .Animal Reproduction Science,2007,101(1-2): 85–96.
[8] Yuan Y, Krisher RL .Effect of ammonium during in vitro maturation on oocyte nuclear maturation and subsequent embryonic development in pigs.Animal Reproduction Science,2010,117(3-4):302–307.
[9] Jian-Min Shi, Xiu-Zhi Tian, Liu GS, et al.Melatonin exists in porcine fol licular fuid and improves in vitro maturation and parthenogenetic development of porcine oocytes.J Pineal Res,2009,47(4):318–323.
[10] Kang JT, Koo OJ, Kwon DK, et al.Eects of melatonin on in vitro maturation of porcine oocyte and expression of melatonin receptor RNA in cumulus and granulose cells.J Pineal Res, 2009, 46:22–28.
[11] Youngs CR.Factorsn fluencing the success of embryoinpig.Ther iogenology,2001,56(8):1311-1320.
[12] Li GP, White KL, Bunch TD, et al.Review of enuc leation methods and procedures used in animal cloning: state of the art.Cloning Stem Cells,2004,6:5–13.
[13] Nour MS, Takahashi Y.Preparation of young preact ivated oocytes with high enuc leation effciency for bovine nucleartrans fer.Theriogenology ,1999,51(3):661–666.
[14] Mitalipov SM, White KL, Far rar VR,et al.Deve lopment of nucleartrans fer and par thenogenet ic rabbi t embryos ac t ivated with inositol 1,4,5-trisphosphate.BiolReprod,1999,60:821–827.
[15] Liu JL,WangMK, Sun QY,et al.Ef fect of telophase enucleation on bovine somatic nuclear transfer.Theriogenology ,2000,54:989–898.
[16] Lee JH, Campbell KH.Effects of enucleation and caffeine on maturation-promoting factor(MPF)and mi togen-activated protein kinase(MAPK)activities in ovine oocytes used as recipient cytoplasts for nuclear transfer.Biol Reprod,2006,74:691–698.
[17] Lee J-H, Panarce M, Medina M,et al.Establishment of pregnancy by ovine nuclear transfer embryos reconst ructed using caf feine-treated in vitro matured oocytes as cytoplastrecipients.Reprod Fertil Dev ,2006,;18:135–136.
[18] Tani T, Shimada H, Kato Y,et al.Demecolcineassisted enucleation for bovine cloning.Cloning Stem Cells, 2006,8:61–66.
[19] Zernicka-Goetz M, Kubiak J, Antony C,et al.Cy toske letal organization of rat oocy tes dur ing metaphase IIarrest and following abortiveactivation: a study by confocal laser scanning microscopy.Mol Reprod Dev,1993, 35:165–175.
[20] Ibanez E, Albertini DF, Overstrom EW.Demecolcine-induced oocy teenucleation for somaticcell cloning: coordination between cell-cycle egress, kinetics of cortical cytoskeletal interactions,and second polar body ext rusion.Biol Reprod,2003,68(4):1249–1258.
[21] Wang MK, Liu JL, Li GP, et al.Sucrose pretreatment for enucleation: an effcient and non-damage method for removing the spindle of the mouse MII oocyte.Mol Reprod Dev,2001,58:432–436.
[22] Liu L,Oldenbourg R,Trimarchi JR,et al.A reliable,noninvasive technique for spindle imaging and enuc l ea tion of mamma lianoocytes.Nat Biotechnol ,2000,18:223–225.
[23] Tatham BG, Dowsing AT, Trounson AO.Enucleation by cent ri fugation of in vitro-matured bovine oocytes for use in nuclear transfer.Biol Reprod,1995,53:1088–1094.
[24] Robertson SA, Jasper MJ, Norman RJ,et al.Granulocyte-macrophage colony-s timulating factor promotes glucose transport and blastomere viability in murine preimplantation embryos.Biol Reprod,2001,64(4):1206–1215.
[25] Emond V,Mac Laren LA, Kimmins S,etal.Expression of cyclooxygenase-2 and granulocytemacrophage colony-st imulat ing factor in the endometrial epitheliumof the cow is up-regulated dur ing early pregnancy and in response to int rauterine infusions of inter feron-tau.Biol Reprod,2004,70(1):54–64.
[26] Barbara Loureiro, Luciano Bonil la, Jeremy Block,et al.Colony-Stimulating Factor 2(CSF-2) Improves Development and Post transfer Survival of Bovine Embryos Produced in Vitro.Endocrinology,2009,150(11):5046-5054.
[27] Akiyama T, Nagata M, Aoki F.Inadequate histone deacety lation dur ing oocyte meiosis causes aneuploidy and embryo death in mice.Proc Nat l Acad Sci,2006,103:7339–7344.
[28] Kim JM, Liu H, Tazaki M,et al.Changes in histone acetylation during mouse oocyte meiosis.J Cell Biol,2003,162:37–46.
[29] Qinggang Meng,Zsuzsanna Polgar,Jun Liu,et al.Live Birth of Somatic Cell-Cloned Rabbi ts following Trichostatin A Treatment and Cotransfer of Par thenogenetic Embryos.Cloning Stem Cells,2009,11(1):203-208.
[30] Shi W,Hoeflich A,Flaswinkel, et al.Induction of a senescent-l ike phenotype does not confer the ability of bovine immor talcells to support the development of nucleart ransfer embryos.Biol Reprod,2003,69(1):301–309.
[31] Enright BP, Kubota C, Yang X, et al.Epigenetic character istics and development of embryos cloned from donor cells treated by trichostatin A or 5-aza-2 -deoxycytidine.Biol Reprod,2003,69(3):896–901.
[32] Wang F, Kou Z,Zhang Y,et al.Dynamic reprogramming of histone acetylation and methylation in the first cell cycle of cloned mouse embryos.Biol Reprod,2007,77(6),1007–1016.
[33] Zhao J, Ross JW,Prather RS,et al.Signi ficant Improvement in Cloning Efficiency of an Inbred Miniature Pig by Histone Deacetylase Inhibitor Treatment after Somatic Cell Nuclear Transfer.Biology of reproduction,2009, 81(3), 525–530.
[34] Li J, Svarcova O, Vaj ta G., et al.High in vitro development after somaticcell nuclear transfer and trichostatin A treatment of reconst ructed porcine embryos..Ther iogenology,2008, 70(5)800–808.
[35] Har de land R.Ant ioxidat ive protec tion by melatonin: multiplicity of mechanisms from radical detoxi fcation to radical avoidance.Endocrine,2005,27(2):119–130.
[36] Rosen J, Than NN, Koch D, et al.Interactions of melatonin and its metabol ites with the ABTS cation radical: ex tension of the radi cal scavenger cascade and formation of novel class of oxidation produc ts, C2-subst i tuted 3-indolinones.J Pineal Res, 2006, 41(4):374–381.
[37] Ishizuka B, Kuribayashi Y, Murai K,et al.The ef fect of melatonin on in vitro fer ti l ization and embryo development in mice.J Pineal Res,2000, 28:48–51.
[38] Yuichiro Kitagawa,Keita Suzuki,Akihiro Yonedaa,et al.Effects of oxygen concent ration and ant ioxidants on the invitro developmental ability, production of reactive oxygen species(ROS),and DNA fragmentation in porcine embryos.Theriogenology ,2004,62(7):1186–1197.
[39] Papis K, Poleszczuk O, Wenta-Muchalska E,Modlinski JA.Melatonin effecton bovine embryo development in vitro in relation to oxygen concen tration.J Pinea l Res,2007,43(4):321–326.
[40] Lee J, You J, Kim J, et al.Postactivation treatment with nocodazole maintains normal nuclear ploidy of c loned pig embryos by increasing nuclear retention and formation of single pronucleus.Theriogenology, 2010,73(4):429–436.
[41] Song K, Hyun SH, Shin T, et al.Post-activation treatment with demecolcine improves development of somatic cellnuclear transfer embryos in pigs by modi fying the remodel ing of donor nuclei.Mol Reprod Dev, 2009,76(7):611–619.
猜你喜歡
瘋狂英語·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數(shù)理化(高中版.高考數(shù)學)(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經(jīng)濟(2017年6期)2017-07-12 09:27:16
中國衛(wèi)生(2016年9期)2016-11-12 13:27:54
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國洗滌用品工業(yè)(2015年7期)2015-02-28 19:02:38
電子設(shè)計工程(2015年12期)2015-02-27 12:06:10
中國衛(wèi)生(2014年11期)2014-11-12 13:11:32
