植物AGAMOUS基因調控機制研究進展
2011-07-30 09:52:26徐雷王利琳
浙江農業科學 2011年6期
關鍵詞:功能
徐雷,王利琳
(1.江蘇新天地生物肥料工程中心有限公司,江蘇無錫214200;2.杭州師范大學生命與環境科學學院,浙江杭州310036)
20世紀90年代,Coen和Meyerowitz對擬南芥和金魚草花器官突變體(主要是MADS-box轉錄因子的代表性突變體)進行遺傳分析,提出了花器官形態建成的粗糙的“ABC模型”,后來經過眾多學者的修改和補充,該模型發展為“ABCDE”模型。其中,C功能基因AGAMOUS(AG)在花器官發育中扮演著重要的角色,對果實和種子的形成也至關重要。花發育調控基因的上下游基因及其功能是目前植物發育遺傳學的研究熱點之一,AG基因是最早被克隆的花發育調控基因,其上下游基因及其功能研究更是其中的熱點,明確其作用分子機制對于農藝、園藝和基因控制技術具有重要的價值。
1990年,Yanofsky等在擬南芥中首次克隆了花同源異型基因AG。在擬南芥中,AG與APETALA1(AP1)基因、APETALA2(AP2)基因相互拮抗,同時AG還與WUSCHEL(WUS)相拮抗,終止WUS的花分生組織指定,它還是第4輪花器官發育的指定基因。在胚珠的發育中,AG也發揮著重要的調控作用。在其他一些物種中,AG同源基因相對保守,但是部分發生了復制,而且彼此功能冗余。雖然目前對AG基因已進行了較多的研究,但是對于在花發育調控網絡中,AG的上下游基因及其作用的分子機制,目前還沒有文章進行綜述。
1 AG序列分析及特征
AG早在裸子植物和被子植物分化之前就已經出現,并且在隨后3億年的進化過程中功能保守[1]。目前已有20多種植物(包括裸子植物和被子植物)的AG同源基因被克隆。序列分析和基因表達研究也表明,盡管這些基因在序列上有差異,但功能相近[2]。
AG基因序列分為5個區域,從5’端開始依次命名為N末端、M區(編碼約57個氨基酸的MADS-box區)、I區(Intervening region)、K區和C末端。M區高度保守,I區和K區相對保守,C末端特異性較強。其編碼285個氨基酸的轉錄因子,含有9個外顯子,8個內含子。
AG亞家族的C類基因(通過進化樹分析,所有AGAMOUS-LIKE基因分為3類:C類、D類(基于該進化枝含有矮牽牛FBP7和FBP11)、裸子植物類),在C末端具有2個相對保守的區域,被稱為AGⅠ區和AGⅡ區[3]。本研究通過對最近發布的豆目植物AG序列豌豆AG同源基因PsAG[4]、百脈根LjAGa和LjAGb[1]進行軟件預測氨基酸序列比對,也發現這2個相對保守區的存在(圖1)。結合高志紅等[2]的判斷,可以驗證此為AG同源基因的特征之一。
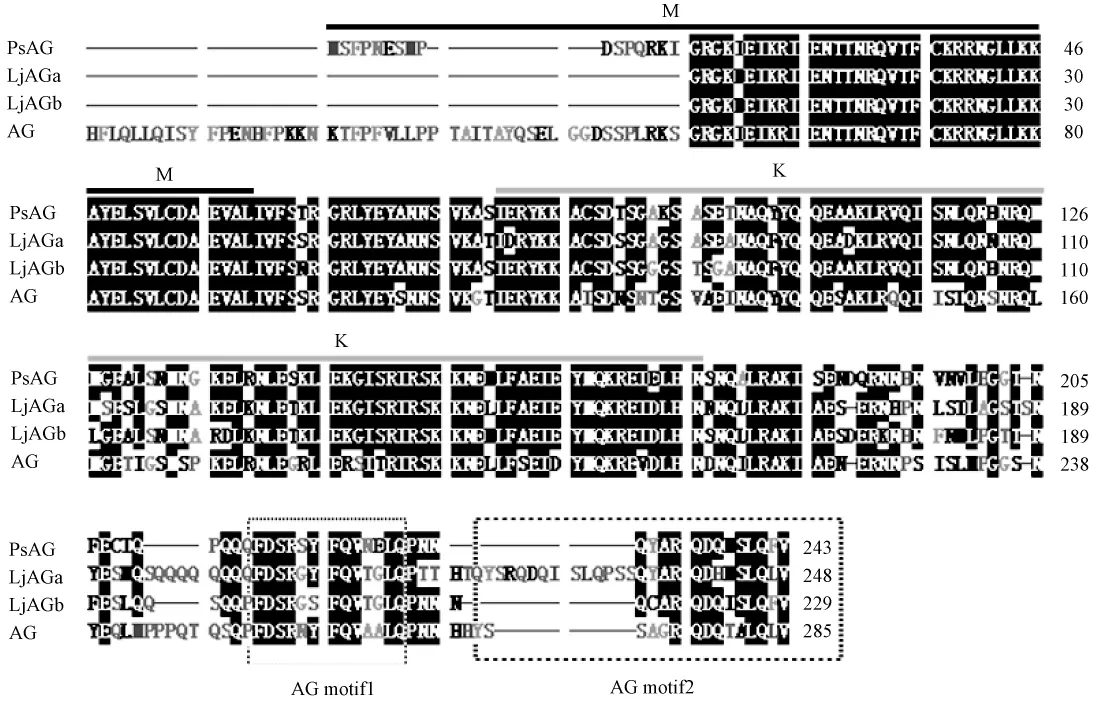
圖1 PsAG、LjAGa、LjAGb、AG蛋白預測氨基酸序列比對
2 擬南芥AG分子作用機制
自從1990年在擬南芥中克隆到AG之后,至今,對花發育及控制花形態建成的分子遺傳機制的研究取得了很多的進展,主要都是建立在對擬南芥研究的基礎之上。擬南芥花自外向內,第1輪為萼片,第2輪為花瓣,第3輪為雄蕊,第4輪為心皮。
2.1 擬南芥AG時空表達模式
在擬南芥野生型花中,AG mRNA和蛋白在花發育第3階段,花的第3、4輪檢測到[5]。AG表達的時空性保證了花器官屬性,花器官數量和模式的正確[6]。
2.2 擬南芥AG功能
目前公認AG至少具有4個功能:抑制A功能基因;終止花分生組織;指定雄蕊和心皮屬性;調控胚珠的發育。AG的正確表達需要第2個內含子,它有3 kb大小,含有2個CArG-box[7],是調控蛋白的目標序列[7-8],如正調控蛋白LFY、WUS,負調控蛋白LUG、SEU、BLR、AP2等。
2.3 擬南芥AG與上下游基因分子作用機制
在花發育調控網絡中,AG與眾多基因相互作用。在不同的途徑中,其位置處于相對的上游或下游,這是復雜的調控網絡所導致的。在此,每個作用途徑中,我們分別加以闡述(圖2)。
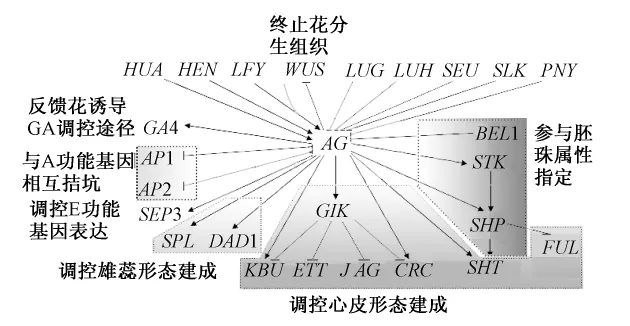
圖2 AG上下游作用基因及其功能(箭頭為正調控,短線為負調控)
AP1和AP2抑制AG在第1、2輪的表達,而AG也抑制它們在第3、4輪的表達。對于AP1,是由SEUSS(SEU)或SEUSS-like
(SLK)蛋白作為銜接蛋白把LEUNIG(LUG)蛋白與AP1蛋白和SEP3蛋白結合,構成復合體,直接結合AG第2個內含子,抑制AG在第1、2輪的表達。LEUNIG-HOMOGOG(LUH)是LUG的旁系同源基因,LUH蛋白也是與SEU蛋白組成復合體,抑制AG的表達。在負調控AG時,LUG比LUH更重要[7,9-10]。在環境脅迫條件下,LUG和LUH功能有所不同[10]。
LFY蛋白是AG表達的直接激活子[11],但其本身還不足以誘導AG,需要其他的因子輔助,可能是WUS[11]。酵母實驗顯示,LFY和WUS分別獨立地結合AG第2個內含子的不同區域[12],但是AG的轉錄只有在WUS和LFY同時存在時,才被激活[13]。WUS和LFY之間作用部分冗余,因為WUS的過表達能夠減少AG激活時對LFY的需求,而且wus和lfy突變體顯示出累加的表型[12]。至于WUS和LFY兩者之間如何相互作用,還不清楚。WUS是終止花分生組織的關鍵且明確的調節開關[11]。WUS激活AG,而AG表達后以非直接的方式抑制WUS[13],這樣產生一個負反饋環,以調節細胞增生與花分生組織的分化[12]。AG雖然沒有影響WUS表達區域,但是在約第7階段,心皮原基形成時,AG達到足夠的水平,關閉了WUS的表達[11,14],以終止花分生組織。除了AG,WUS也受到CLV途徑的負調控。AG和CLV途徑主要是獨立發揮功能,因為ag和clv1突變體在分生組織決定上的效果是累加的[12-13]。
BELLRINGER(BLR即PENNYW ISE(PNY))編碼一個BELL同源結構域蛋白,BLR蛋白直接結合到AG的第2內含子,抑制AG表達。blr突變體老的莖終端花中萼片轉變為心皮[8]。
HUA和HEN基因可能在AG RNA加工中發揮功能,在花發育后期維持AG的表達。HUA1蛋白類似CCCH鋅指蛋白,能結合單鏈RNA和DNA;HUA2蛋白含有核定位信號,有轉錄激活活性。HUA ENHANCER 1(HEN1)蛋白是個核蛋白,是microRNA積聚所需要的;HEN2蛋白類似DexH-box RNA解旋酶,有核定位信號,可能是核酸外切酶的一個成分,它還參與了AG mRNA前體的加工[15]。HEN4蛋白含有KH結構域,是RNA結合蛋白,與HUA1蛋白定位相同并且相互作用,在營養器官發育中也發揮作用[15]。HEN4,HUA1和HUA2可能阻止了發生在AG第2個內含子內的轉錄提前終止,通過利于AG mRNA前體的加工來發揮在花生殖器官屬性和花分生組織終止指定中的作用[15]。另外,HUA1,HUA2,HEN1功能冗余地維持B類功能基因的表達,它們也在營養器官發育中發揮作用[16]。
上述基因主要作為相對的AG上游基因發揮作用,調控AG的時空表達模式或調控其表達量,以此為基礎,AG再發揮相應的功能。LFY和WUS促進AG表達,AP1和AP2調控其限于心皮和雄蕊中表達,HUA和HEN則負責調控AG的正確轉錄和加工。在花發育后期,BLR調控莖終端花中AG的表達部位;而HUA和HEN維持其表達。
AG調控著下游約1 000個基因的表達[17]。其中已確定AG蛋白直接與AG、AP3、CRC、SEP3、DAD1和GA4啟動子CArG-box結合,促進其表達[18-19]。其中,AG通過調控GA4的表達,促進GA的生物合成[18],以對GA開花誘導途徑提供反饋。
AG主要通過調控SPL和DAD1,調控雄蕊的形態建成。
AG調控SPL的表達,誘導小孢子產生,形成花粉。在花發育第5~6階段,AG足以激活SPL發揮功能[17],但是并不參與維持其表達[20]。另外,還有其他的輪特異因子參與小孢子發生的誘導[20]。
在花發育后期階段,AG是花藥形成和分裂以及花絲的建成和延長所必需的[17]。DEFECTIVE IN ANTHER DEHISCENCE1(DAD1)編碼茉莉酸生物合成起始步驟的催化酶,在花發育后期,AG蛋白結合到DAD15’編碼區中10 bp的CArG區,上調DAD1在雄蕊中的表達[17],而茉莉酸控制著雄蕊的成熟,這樣,AG通過控制茉莉酸的合成起始,從而協調雄蕊的成熟。DAD1還受到AUXIN RESPONSE FACTOR 6(ARF6)和AUXIN RESPONSE FACTOR 8(ARF8)的正調控,通過茉莉酸途徑,控制著內三輪花器官的長度,影響花的開放[21]。
AG通過調控SPT、CRC、GIK、KNU等基因,調控心皮的形態建成。
SPATULA(SPT)編碼螺旋—環—螺旋轉錄因子,控制著心皮邊緣和傳導管(the transmitting tract)的細胞增殖[18]。它的表達可能同時受到AG和SHP的冗余調控[22]。
CRABSCLAW(CRC)表達于心皮的遠軸部位,參與到心皮原基遠軸組織(如外部表皮組織)的發育[18],也涉及雌蕊和蜜腺的指定[19]。CRC可以在AG不存在的情況下,指導心皮發育,但是AG可以強化其表達。CRC也可以激活AG[18]。
AG直接結合到GIK下游732bp的CArG box,上調它的表達。GIANT KILLER(GIK)編碼一個AT-hook類型的DNA結合蛋白,AT-hook基序結合目標DNA序列核基質附著區(Matrix attachment regions,MARs)的DNA雙螺旋的小溝,不同于通常的轉錄因子主要結合大溝。植物中,對AT-hook基序蛋白在發育中的作用了解還很少[19]。GIK蛋白持續結合到ETT的MARs,負調控ETT的表達,而且這種抑制與ETT啟動子中H3K9的二甲基化的動態變化密切相連。在體外,GIK直接結合到CRC、JAG和KNU的MARs,抑制它們的轉錄,在體內,gik突變體中,JAG和KNU的mRNA水平相對較高,CRC輕微增加[19]。ETTIN(ETT,AUXIN RESPONSE FACTOR3)與AUXIN RESPONSE FACTOR 4(ARF4)作用冗余,參與花分生組織和生殖器官的近遠軸模式,以及雌蕊的頂基軸模式建成[19]。JAGGED(JAD)、KNUCKLES(KNU)和NUBBIN(NUB)編碼C2H2鋅指轉錄因子,作用冗余地促進雄蕊和心皮原基的增生[19]。KNU涉及花分生組織的決定和配子體的指定,它在分生組織的表達直接受到AG的調控[19]。AG參與胚珠的發育,此結論最初來自于ap2單突變與ap2 ag雙突變的比較。在ap2單突變體中,花瓣大部分缺失,萼片轉變成心皮結構,在上面有胚珠形成,其中一些轉變成心皮化的結構。而ap2 ag雙突變體中,萼片也表現心皮屬性,但是胚珠轉變成心皮結構,數目顯著增加,這暗示著AG也參與胚珠的指定[23]。其作用的分子機制可能是由于SHP1,SHP2,STK同時受到AG的調控,而它們控制胚珠的發育。在心皮中,AG蛋白和SEP蛋白可能與STK或SHP蛋白之一組成穩定的復合體促進胚珠發育,同時這些復合體中任何一個可能都足夠促進心皮發育[24]。
在擬南芥中,胚珠作為新的分生組織從心皮組織中形成。WUS在珠心中表達,類似在莖頂端和花分生組織中的作用方式,以非細胞自主的形式,控制胚珠原基中細胞增生[23]。隨后,增生細胞命運由胚珠屬性因子所指定,如同源異形因子BELL1(BEL1)和MADS-box家族成員SEEDSTICK(STK)、SHATTERPROOF1(SHP1)、SHP2及AG[23],而WUS的表達再次受到AG和SEP3蛋白的抑制[5],BEL1可能在蛋白質水平負調控AG并且維持WUS的抑制狀態[23]。對于胚珠中,WUS短暫的表達起始,其調節機制現在還不了解。
通過酵母三雜交和遺傳分析,Brambilla等[23]發現SHP1、SHP2、STK和AG分別與SEP蛋白形成二聚體后和BEL1相互作用,控制珠被的屬性和形成。
FRUITFULL(FUL)也與心皮發育有關,參與調控受精后心皮瓣(valve)細胞分化和增大,可能與SHP相互拮抗[25]。
SEP蛋白在AG發揮功能時,也具有重要的作用。SEP蛋白能夠建立促進心皮和胚珠發育的蛋白間的相互作用,沒有足夠的SEP活性,胚珠發育中的主要基因無法發揮功能[24]。其中,在促進心皮發育時,SEP3比SEP1、SEP2更有效,而AG直接促進SEP3的表達[24],這也有利于自身功能的發揮。
AG在不同的時期發揮不同的功能(圖3)。目前對A調控花藥和花絲形態建成時的目標基因還不明確,對AG調控胚珠發育的作用時期也不明確,這些還有待進一步的深入研究。
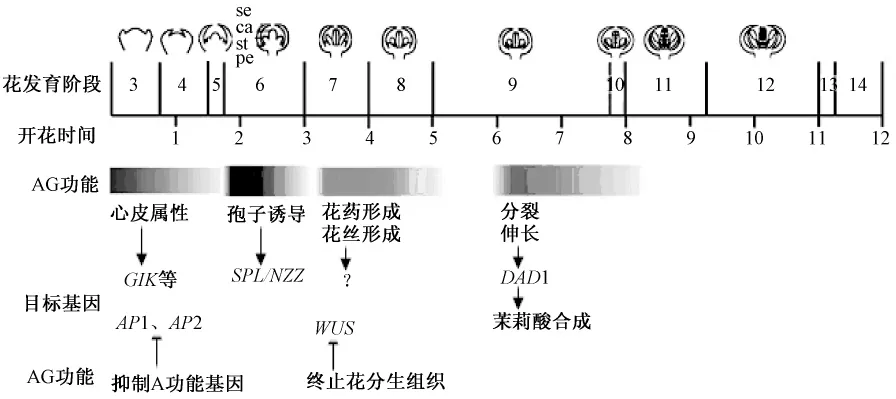
圖3 AG主要作用時間
3 其他植物中AG基因
目前已經克隆了20多種植物的AG同源基因,其中部分出現了復制,如百脈根中,它們的表達模式也發生了變化,其功能還有待進一步的研究。而同樣的豆目植物豌豆中,我們通過豌豆早褐病毒對PsAG的保守序列進行沉默,發現在豌豆中也出現了復制,但是功能相對保守[26]。
在其他植物中,AG花器官屬性指定和花分生組織決定功能,通常不是由單個的AG同源基因來發揮的。在玉米中,這些功能由不同的AG旁系同源基因來執行。在擬南芥中,這2個功能也是可分的,如在擬南芥反義RNA或弱突變(如ag-4,它產生一個內部缺失的AG蛋白)造成的部分AG缺失的植株中就會出現分生組織不終止,但是器官屬性正常,即花內生花的現象。如此表明器官屬性功能并不明顯地與分生組織終止功能重疊。我們通過豌豆早褐病毒,在豌豆中,對PsAG保守序列進行沉默時,也發現了這樣的現象。
在裸子植物中,AG同源物與孢子葉球的發育的聯系,表明AG在生殖器官屬性指定中的角色是古老的,但是同時它也提示這些孢子葉球是決定的,即其分生組織是終止的。
ag突變體中外圍花器官數目增加,但是在玫瑰中,花瓣和雄蕊的增加和最終的花分生組織的終止也可以由花分生組織的定點增大而引起,就像在sup和ult突變體中所見[13]。所以現在對這一結果,還不能肯定的歸為AG的一個功能。
4 展望
AG基因在植物花器官的發育中占有重要位置,與植物的繁殖息息相關,在植物的進化過程中,其功能相對保守,沒有發生較大的變化。
目前確定的4個AG功能,彼此關聯,共同起到調控花正常發育的作用。但是對這些功能發揮的途徑,了解還不多,其下游基因開發較少,還有許多基因有待確定,比如雄蕊形態建成的啟動基因還不清楚。隨著更多下游基因的確定,才能更清晰地認識植物花器官的發育調控網絡,對于植物花器官改造具有重要意義,具有重要的園藝價值。
利用AG的功能來控制外來物種入侵具有重要意義。目前對AG功能的研究已經成功拓展應用到基因控制技術中,Liu等[27]把AG第2個內含子與白喉毒素A片段(Diphtheria toxin A,DT-A)的基因相融合,發現超過89%轉基因擬南芥雄蕊和心皮消除,完全不育。
由于AG基因位于調控植物果實和種子發育的基因上游,明確其不同的調控途徑,無疑對于作物的育種或果樹的掛果研究具有參考價值。
[1]Dong Z C,Zhao Z,Liu C W,et al.Floral Patterning in Lotus japonicus[J].Plant Physiol,2005,137:1272-282.
[2]高志紅,張玉明,王珊,等.植物花發育調控基因AGAMOUS的研究進展[J].西北植物學報,2008,28:638-644.
[3]Kramer E M,Jaramillo M A,Di Stilio V S.Patterns of gene duplication and functional evolution during the diversification of the agamous subfamily of MADS box genes in angiosperms[J].Genetics,2004,166:1011-1023.
[4]Hecht V,Foucher F,Ferrandiz C,et al.Conservation of Arabidopsis flowering genes in model legumes[J].Plant Physiol,2005,137:1420-1434.
[5]Urbanus S L,de Folter S,Shchennikova A V,et al.In planta localization patterns of MADS domain proteins during floral development in Arabidopsis thaliana[J].BMC Plant Biol,2009,9:1-16.
[6]Liu Z.Transcription co-repressors in flower development[J].Chin Bull Bot,2003,20:385-394
[7]Gregis V,Sessa A,Colombo L,et al.AGL24,SHORT VEGETATIVE PHASE,and APETALA 1 redundantly control AGAMOUS during early stages of flower development in Arabidopsis[J].Plant Cell,2006,18:1373-1382.
[8]Bao X,Franks R G,Levin J Z,et al.Repression of AGAMOUS by BELLRINGER in floral and inflorescence meristems[J].Plant Cell,2004,16:1478-1489.
[9]Sridhar V V,Surendrarao A,Liu Z.APETALA1 and SEPALLATA 3 interact with SEUSS to mediate transcription repression during flower development[J].Development,2006,133:3159-3166
[10]Sitaraman J,Bui M,Liu Z.LEUNIG-HOMOLOG and LEUNIG perform partially redundant functions during Arabidopsis embryo and floral development[J].Plant Physiol,2008,147:672-681.
[11]Lenhard M,Bohnert A,Jürgens G,et al.Termination of stem cellmaintenance in Arabidopsis floral meristems by interactions between WUSCHEL and AGAMOUS[J].Cell,2001,105:805-814.
[12]Lohmann JU,Hong R L,Hobe M,et al.A molecular link between stem cell regulation and floral patterning in Arabidopsis[J].Cell,2001,105:793-803.
[13]Sablowski R.Flowering and determinacy in Arabidopsis[J].J Exp Bot,2007,58:899-907.
[14]Zhao L,Kim Y J,Dinh T T,et al.miR172 regulates stem cell fate and defines the inner boundary of APETALA 3 and PISTILATA expression domain in Arabidopsis floral meristems[J].Plant J,2007,51:840-849.
[15]Cheng Y,Kato N,Wang W,et al.Two RNA binding proteins,HEN4 and HUA1,act in the processing of AGAMOUS pre-mRNA in Arabidopsis thaliana[J].Dev Cell,2003,4:453-466.
[16]Jack T.New members of the floral organ identity AGAMOUS pathway[J].Trends Plant Sci,2002,7:286-287
[17]Ito T,Ng K H,Lim T S,et al.The homeotic protein agamous controls late stamen development by regulating a jasmonate biosynthetic gene in Arabidopsis[J].Plant Cell,2007,19:3516-3529.
[18]Gómez-Mena C,Folter S,Rosta M M,et al.Transcriptional program controlled by the floral homeotic gene AGAMOUS during early organogenesis[J].Development,2004,132:429-438.
[19]Ng K H,Yu H,Ito T.AGAMOUS controls GIANT KILLER,a multifunctional chromatin modifier in reproductive organ patterning and differentiation[J].PLoS Biol,2009,7(11):1000251.
[20]Ito T,Wellmer F,Yu H,et al.The homeotic protein AGAMOUS controls microsporogenesis by regulation of SPOROCYTELESS[J].Nature,2004,430:356-360.
[21]Tabata R,Ikezaki M,Fujibe T,et al.Arabidopsis auxin response facror 6 and 8 regulate jasmonic acid biosynthesis and floral organ development via repression of Class 1 KNOX genes[J].Plant Cell Physiol,2010,51:164-175.
[22]Pinyopich A,Ditta G S,Savidge B,et al.Assessing the redundancy of MADS-box genes during carpel and ovule development[J].Nature,2003,424:85-88.
[23]Brambilla V,Battaglia R,Colombo M,et al.Genetic and molecular interactions between BELL1 and MADS Box factors support ovule development in Arabidopsis[J].Plant Cell,2007,19:2544-2556.
[24]Favaro R,Pinyopich A,Battaglia R,et al.MADS-box protein complexes control carpel and ovule development in Arabidopsis[J].Plant Cell,2003,15:2603-2611.
[25]Liljegren S J,Ditta G S,Eshed Y,et al.SHATTERPROOF MADS-box genes control seed dispersal in Arabidopsis[J].Nature,2000,404:766-770.
[26]徐雷,宋偉杰,王利琳.豌豆AGAMOUS同源基因功能的初步研究[J].科學通報,2009,54:3207-3212.
[27]Liu Z R,Liu Z C.The second intron of agamous drives carpeland stamen-specific expression sufficient to induce comp lete sterility in Arabidopsis[J].Plant Cell Rep,2008,27:855-863.
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
