大豆疫霉拮抗細(xì)菌B048液體發(fā)酵培養(yǎng)基的篩選與發(fā)酵條件的優(yōu)化
2011-11-14 07:56:08趙平生王子迎
合肥師范學(xué)院學(xué)報(bào) 2011年6期
關(guān)鍵詞:大豆
汪 莞, 趙平生, 周 濤, 王子迎
(合肥師范學(xué)院生命科學(xué)系,安徽合肥 230601)
大豆疫霉拮抗細(xì)菌B048液體發(fā)酵培養(yǎng)基的篩選與發(fā)酵條件的優(yōu)化
汪 莞, 趙平生, 周 濤, 王子迎
(合肥師范學(xué)院生命科學(xué)系,安徽合肥 230601)
通過單因子實(shí)驗(yàn),篩選得到了適合大豆疫霉拮抗細(xì)菌B048液體發(fā)酵的最佳碳源為玉米粉,最佳氮源為豆粕,最佳無機(jī)鹽離子為氯化鈉。通過正交實(shí)驗(yàn)確定了該培養(yǎng)基的最佳配比為:玉米粉5 g/L、豆粕15 g/L、氯化鈉10 g/L。在確定適合該菌株的培養(yǎng)基組分基礎(chǔ)上,對(duì)其培養(yǎng)條件進(jìn)行優(yōu)化,結(jié)果表明:初始p H值為6.0,培養(yǎng)時(shí)間為48 h,轉(zhuǎn)速為180 r/min時(shí)為該菌株的最佳培養(yǎng)條件。
大豆疫霉;B048;培養(yǎng)基;發(fā)酵
大豆疫霉(Phytophthorasojae)侵染大豆引起的疫霉根腐病是大豆生產(chǎn)上的毀滅性病害[1],最早于1948年發(fā)生在美國(guó),此后不斷蔓延,現(xiàn)已廣泛分布亞、非、歐、南北美洲的近二十個(gè)國(guó)家的大豆主要產(chǎn)區(qū),每年給大豆生產(chǎn)帶來的直接經(jīng)濟(jì)損失高達(dá)幾十億美元,成為大豆生產(chǎn)上最嚴(yán)重的病害[2,3]。該病菌于80年代后期首次在我國(guó)的東北地區(qū)發(fā)現(xiàn),隨后該病在我國(guó)北方的大豆主產(chǎn)區(qū)逐步擴(kuò)展,目前已經(jīng)成為黑龍江省大豆生產(chǎn)上的頭號(hào)病害,每年造成巨大的經(jīng)濟(jì)損失[4]。90年代后期,在我國(guó)的局部大豆產(chǎn)區(qū),也發(fā)現(xiàn)了大豆疫霉的傳入,在條件適宜的年份,其造成的大豆產(chǎn)量損失高達(dá)50%以上[5-8]。
目前,大豆疫霉根腐病的防治方法主要是化學(xué)防治,其中使用最多的藥劑是甲霜靈(Metalaxy,商品名Ridomil)。甲霜靈為內(nèi)吸性殺菌劑,生物活性強(qiáng),且效期持久[9]。但由于甲霜靈對(duì)病菌的作用位點(diǎn)單一,病原菌容易對(duì)其產(chǎn)生抗性突變,因此,開發(fā)新的替代防治方法在控制大豆疫霉根腐病上尤顯重要[10]。在大豆疫霉根腐病的防治中,未見從土壤中篩選拮抗菌作為病害生防因子的報(bào)道。為可持續(xù)地控制大豆疫病,避免大豆疫病因單一的化學(xué)防治而過快地產(chǎn)生抗藥性,生物防治已成為大豆疫病防治的重要研究領(lǐng)域。
為此,作者從大豆疫霉生存的土壤中篩選具有高拮抗活性和易定殖的生物防治菌株,分離到一株拮抗效果好的細(xì)菌,命名為B048。為實(shí)現(xiàn)該生防菌株的產(chǎn)業(yè)化,本文對(duì)該菌株的液體發(fā)酵培養(yǎng)基的篩選和發(fā)酵條件進(jìn)行了優(yōu)化研究,以期為該菌株的工業(yè)生產(chǎn)并獲得高產(chǎn)抗菌物質(zhì)提供參考依據(jù)。
1 材料與方法
1.1 材料
大豆疫霉生防菌B048由本實(shí)驗(yàn)室在土壤中分離得到,保存于本實(shí)驗(yàn)室。基礎(chǔ)培養(yǎng)基和種子培養(yǎng)基為肉湯培養(yǎng)基(蛋白胨10.0 g/L、氯化鈉5.0 g/L、葡萄糖5.0 g/L、牛肉浸出粉3.0 g/L)。
1.2 方法
1.2.1 種子菌液制備
無菌條件下,用接種環(huán)挑取少量大豆疫霉生防菌B048菌株,在肉湯培養(yǎng)基上劃線,用封口膜封口后,放入37℃恒溫箱中培養(yǎng),48h后,得到活化的大豆疫霉生防菌B048菌株單菌落,然后挑取少量活化的大豆疫霉生防菌B048單菌落于肉湯培養(yǎng)基中,28℃、110 r/min條件下培養(yǎng)12 h,得到種子菌液。
1.2.2 活菌數(shù)計(jì)數(shù)
按平板菌落計(jì)數(shù)法計(jì)算活菌數(shù):將種子菌液稀釋至10-4-10-7,然后取100μL均勻涂布到肉湯培養(yǎng)基平板上,12 h后計(jì)算活菌數(shù)。
1.2.3 培養(yǎng)基組分的篩選
碳源篩選時(shí),分別以含量為5 g/L的玉米粉、可溶性淀粉、蔗糖、乳糖、麥芽糖、D-果糖共6種碳源替代肉湯培養(yǎng)基中的葡萄糖,其他成分不變;氮源篩選時(shí),分別以含量為15g/L的黃豆粉、豆粕、硝酸鉀、硝酸鈉、硝酸銨、硫酸銨共6種氮源替換肉湯培養(yǎng)基中的牛肉膏和蛋白胨;無機(jī)鹽離子的篩選時(shí),以優(yōu)化的碳源與氮源制成液體培養(yǎng)基,然后在該培養(yǎng)基中分別添加5 g/L的MgSO4、KH2PO4、CaCO3,以不添加金屬離子的培養(yǎng)基作為對(duì)照。將不同的培養(yǎng)基均用1 mol/L的HCl和0.1 mol/L的NaOH調(diào)至p H為7.5后滅菌20 min。無菌操作下每種液體培養(yǎng)基各倒三個(gè)250 mL錐形瓶,每個(gè)錐形瓶的裝液量為150 mL,用接種環(huán)挑取少量的大豆疫霉生防菌B048,接種在搖瓶中,放入28℃的搖床上培養(yǎng)24 h。
1.2.4 最佳培養(yǎng)條件的篩選
確定最佳培養(yǎng)時(shí)間時(shí),將菌株培養(yǎng)時(shí)間調(diào)節(jié)為12、24、36、48、60、72h;確定最佳培養(yǎng)基初始p H值時(shí),用0.1 mol/L檸檬酸和0.1 mol/LNaOH將優(yōu)化培養(yǎng)基p H值調(diào)節(jié)為4.0、6.0、7.5、9.0和11.0;確定最佳培養(yǎng)轉(zhuǎn)速時(shí),將搖床轉(zhuǎn)速分別調(diào)整為60、120、180、240 r/min。每種液體培養(yǎng)基各倒三個(gè)250 m L錐形瓶,裝液量為150 m L,接種后放入28℃的搖床上培養(yǎng)24 h。
2 結(jié)果與分析
2.1 培養(yǎng)基的優(yōu)化
2.1.1 碳源
由圖1可以看出,麥芽糖作為碳源時(shí),B048的生長(zhǎng)量最大,達(dá)到2.25×1010cfu/m L,其他碳源的菌株生長(zhǎng)量大小依次為玉米粉、可溶性淀粉、D-果糖、葡萄糖、乳糖和蔗糖,但由于麥芽糖的市場(chǎng)價(jià)格高于玉米粉,同時(shí)以玉米粉作為碳源的液體培養(yǎng)基的活菌數(shù)也不低,達(dá)到了1.77×1010cfu/m L,所以,綜合菌株生長(zhǎng)量和生產(chǎn)成本的便于獲取和價(jià)格低廉,最終選擇玉米粉作為大豆疫霉拮抗細(xì)菌B048的液體發(fā)酵培養(yǎng)基中的最佳碳源。
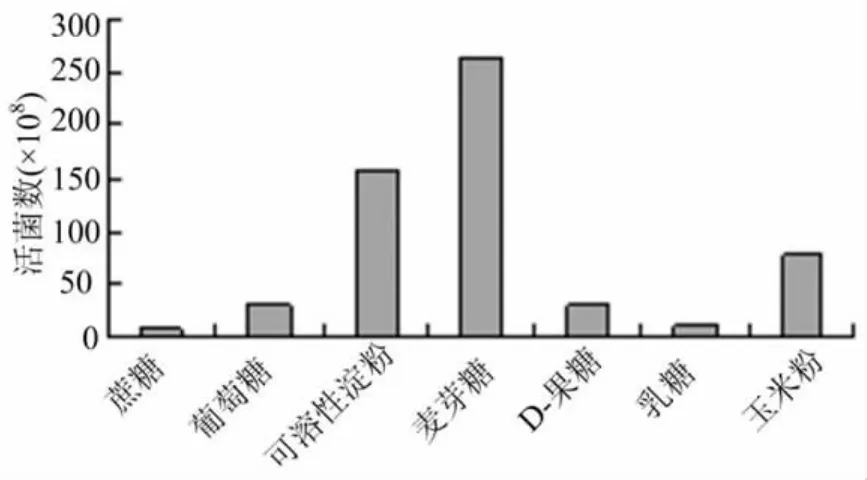
圖1 不同碳源對(duì)大豆疫病生防菌B048生長(zhǎng)的影響
2.1.2 氮源
由圖2可知,黃豆粉作為氮源時(shí),大豆疫霉拮抗細(xì)菌B048的液體發(fā)酵培養(yǎng)基中,活菌數(shù)最多,達(dá)到1.54×1010cfu/m L,其他氮源的菌株生長(zhǎng)量大小依次為牛肉浸膏和蛋白胨、硝酸鈉、豆粕、硫酸銨、硝酸鉀和硝酸銨。另一方面,豆粕作為一種工業(yè)下腳料,便于獲取且價(jià)格低廉,同時(shí)以豆粕作為氮源的液體培養(yǎng)基的活菌數(shù)也達(dá)到了7.7×109cfu·m L-1,所以,綜合成本因素和培養(yǎng)效果的考慮,選擇豆粕作為大豆疫霉拮抗細(xì)菌B048的液體發(fā)酵培養(yǎng)基中的最佳氮源。

圖2 不同氮源對(duì)大豆疫病生防菌B048生長(zhǎng)的影響
2.1.3 無機(jī)鹽離子
以優(yōu)化的碳源和氮源制成的液體培養(yǎng)基,分別加入等量的不同的無機(jī)鹽離子,結(jié)果(圖3)表明,Na+能促進(jìn)菌株生長(zhǎng),使菌株的生產(chǎn)量顯著增加,K+對(duì)菌株的生長(zhǎng)影響不明顯,Ca2+、Mg2+對(duì)菌株的生長(zhǎng)有輕微的抑制作用,加入氯化鈉作為無機(jī)鹽離子的液體培養(yǎng)基,獲得的大豆疫霉拮抗細(xì)菌B048的活菌數(shù)遠(yuǎn)高于加硫酸鎂、磷酸二氫鉀和碳酸鈣作為金屬離子的液體培養(yǎng)基,菌株生長(zhǎng)量達(dá)到了2.85 ×109,所以選擇氯化鈉作為大豆疫霉拮抗細(xì)菌B048的液體發(fā)酵培養(yǎng)基中最佳的無機(jī)鹽離子。

圖3 不同無機(jī)鹽離子對(duì)大豆疫病生防菌B048生長(zhǎng)的影響
2.1.4 正交試驗(yàn)法優(yōu)化篩選培養(yǎng)基各組分的配比
由表1和表2可知,不同水平的玉米粉對(duì)發(fā)酵液中活菌數(shù)有一定的影響,培養(yǎng)基中玉米粉的含量越低,發(fā)酵液中活菌數(shù)越多,反之越少,極差為111.8,中、高含量的玉米粉的發(fā)酵液中,活菌數(shù)的含量相差不大,豆粕含量適中時(shí),活菌數(shù)含量最多,低或高的豆粕含量都會(huì)降低發(fā)酵液中的活菌數(shù)量,極差為154.7,氯化鈉含量對(duì)發(fā)酵液中活菌數(shù)量的影響最大,極差達(dá)到了167.9,高含量的氯化鈉會(huì)抑制菌株的活性,適中的氯化鈉含量有利于活菌數(shù)的增加。
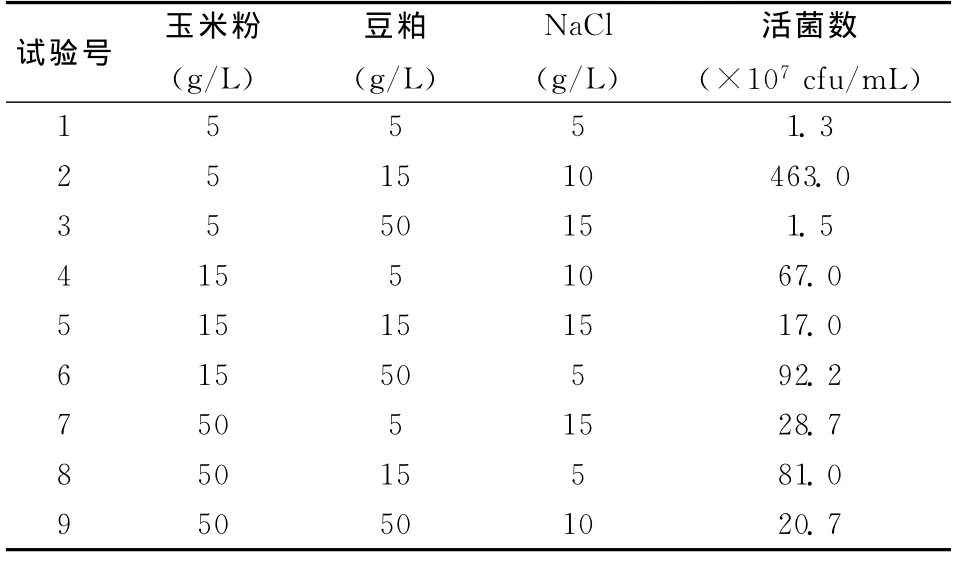
表1 3因素3水平的正交試驗(yàn)統(tǒng)計(jì)結(jié)果
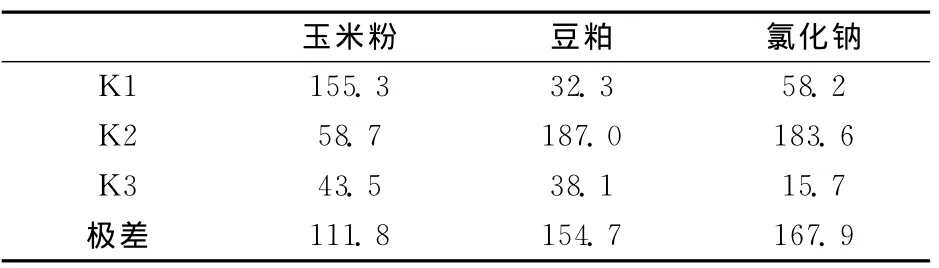
表2 培養(yǎng)基各組分配比優(yōu)化的正交試驗(yàn)活菌數(shù)評(píng)價(jià)結(jié)果(×107 cuf/m L)
2.2 培養(yǎng)條件的確定
2.2.1 培養(yǎng)時(shí)間
結(jié)果(圖4)表明,菌株在培養(yǎng)36-48 h內(nèi),生長(zhǎng)量呈明顯上升趨勢(shì),48h時(shí)達(dá)最大值,活菌數(shù)最多,達(dá)到9.5×108,然后隨著培養(yǎng)時(shí)間的延長(zhǎng)(48-72 h),生長(zhǎng)量呈明顯下降趨勢(shì),因此,菌株以培養(yǎng)48h較為適宜。

圖4 最佳培養(yǎng)時(shí)間的確定
2.2.2 轉(zhuǎn)速
由圖5可以看出,轉(zhuǎn)速?gòu)?20 r/min增加至180 r/min時(shí),菌株的生長(zhǎng)量顯著增加,當(dāng)轉(zhuǎn)速繼續(xù)增加時(shí),菌株生長(zhǎng)量明顯下降,這說明轉(zhuǎn)速影響著菌株B048的生長(zhǎng)周期,當(dāng)轉(zhuǎn)速120 r/min時(shí),培養(yǎng)48 h的B048細(xì)菌還處在對(duì)數(shù)期,當(dāng)轉(zhuǎn)速為180 r/min時(shí),B048培養(yǎng)48 h就進(jìn)入了生長(zhǎng)衰退期,導(dǎo)致活菌數(shù)下降。因此其最適宜的轉(zhuǎn)速為180r/min。

圖5 最佳轉(zhuǎn)速的確定
2.2.3 pH值
圖6表明,在液體發(fā)酵培養(yǎng)基p H值為4.0—6.0時(shí),菌株生長(zhǎng)量呈明顯上升趨勢(shì),在p H值為6.0時(shí),最有利于菌株生長(zhǎng),活菌數(shù)達(dá)到3.15× 107,此后隨著p H值增大,菌株生長(zhǎng)量則明顯下降,當(dāng)p H為9.0—11.0時(shí),幾乎會(huì)抑制活菌數(shù)的含量,因此,可以判定,該菌株適合在中性的環(huán)境下生存。
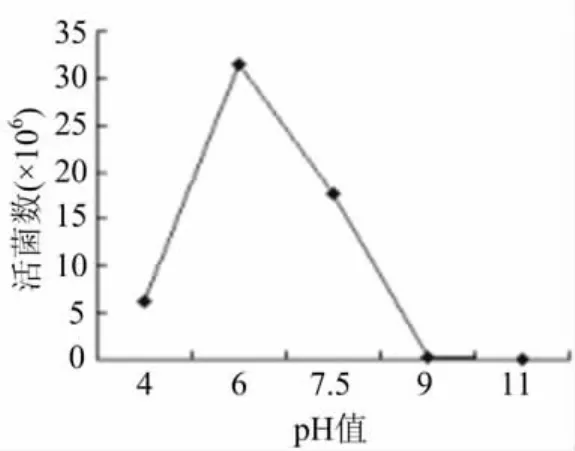
圖6 最佳p H值的確定
3 討論
在用生物防治控制大豆疫霉根腐病方面,從土壤中篩選拮抗菌作為病害生防因子的方法,是非常有潛力的,自然環(huán)境是個(gè)巨大的微生物菌種庫(kù),而土壤則是微生物生活的大本營(yíng),是尋找和發(fā)現(xiàn)有重要應(yīng)用潛力的拮抗菌種的主要來源。況福元等[11]從土壤中分離篩選獲得2株解淀粉芽抱桿菌(Bacillusamyloliquefaciens)對(duì)菜心炭疽病菌希金斯炭疽菌(Colletotrichumhigginsianum)具有強(qiáng)烈抑制作用;麥明曉等[12]從土壤中篩選出一株對(duì)香蕉的尖孢鐮刀菌具有較強(qiáng)抑制作用的師崗鏈霉菌(Streptomycesmorookaense);本實(shí)驗(yàn)的作者從大豆疫霉生存的土壤中篩選具有高拮抗活性和易定殖的生物防治菌株,分離到的一株拮抗效果相對(duì)最好細(xì)菌,命名為B048菌株。
通過單因素試驗(yàn)和正交試驗(yàn)相結(jié)合,初步得到了適合搖瓶條件下生防菌菌株發(fā)酵的液體培養(yǎng)基配方為:玉米粉5 g/L、豆粕15 g/L、氯化鈉10 g/L;在最適培養(yǎng)條件為:初始p H值為6.0,培養(yǎng)時(shí)間為48 h,轉(zhuǎn)速為180 r/min的條件下,生防菌B048的活10 g/L菌數(shù)高達(dá)4.63×109cuf/mL。多種拮抗菌己被證實(shí)對(duì)病原菌具有防治作用,且拮抗菌應(yīng)用于病害防治有很多優(yōu)點(diǎn),例如,大豆疫霉拮抗細(xì)菌B048液體發(fā)酵培養(yǎng)基中使用的玉米粉和豆粕都是工業(yè)下腳料,便于獲取且價(jià)格低廉,為該菌株的工業(yè)化生產(chǎn)節(jié)約了生產(chǎn)成本,另一方面,生防菌B048的活菌數(shù)為4.63× 109cuf/m L,遠(yuǎn)高于目前市場(chǎng)上同類的微生物菌劑的活菌數(shù)含量2.0×108cuf/mL[13]。
本試驗(yàn)只是在搖床條件下對(duì)發(fā)酵培養(yǎng)基的組成及其培養(yǎng)條件進(jìn)行了優(yōu)化,而拮抗菌制劑的生防作用的發(fā)揮與制備工藝、使用方法和菌劑使用時(shí)溫度、濕度、p H值等環(huán)境因素密切相關(guān),所以,在拮抗菌制劑商業(yè)化過程中,一方面要繼續(xù)加大對(duì)拮抗菌活性物質(zhì)發(fā)酵工藝、提取與純化方法等的研究,另一方面加大拮抗菌制劑發(fā)揮最佳作用條件的研究,今后尚需在發(fā)酵罐中和田間實(shí)驗(yàn)里進(jìn)行驗(yàn)證,在實(shí)際生產(chǎn)中對(duì)發(fā)酵條件的控制方面也尚需進(jìn)一步研究;其次,本實(shí)驗(yàn)中對(duì)最佳碳源、氮源、無機(jī)鹽離子和正交實(shí)驗(yàn)法優(yōu)化培養(yǎng)基配比研究,僅僅以液體發(fā)酵液中的活菌數(shù)含量作為指標(biāo),并沒有其他的與之相對(duì)應(yīng)的篩選模型作為依據(jù),例如,對(duì)峙法、牛津杯法、離體植物組織平皿法、活體植株篩選法等[14],因此,可能會(huì)對(duì)實(shí)驗(yàn)結(jié)果造成一定的誤差,在以后的實(shí)驗(yàn)中,可以設(shè)計(jì)類似的實(shí)驗(yàn)方案,如測(cè)定大豆疫霉拮抗細(xì)菌B048對(duì)大豆疫霉的抑菌圈大小,來進(jìn)一步驗(yàn)證實(shí)驗(yàn)的準(zhǔn)確性和完整性;最后,在今后的研究中,可以對(duì)該拮抗菌株B048的活性物質(zhì)進(jìn)行研究,利用活性物質(zhì)分離純化技術(shù),確定其活性物質(zhì)的組成、性質(zhì)和合成途徑,也可以進(jìn)一步研究基因遺傳工程等手段,提高拮抗菌的活性及抑菌譜,這些可能為提高發(fā)酵濾液中活性成分含量的關(guān)鍵條件。
[1] Tyler BM.Phytophthorasojae:root rot pathogen of soybean and model oomycete[J].Molecular Plant Pathology,2007,8(1):1-8.
[2] Schmitthenner AF.Problem and processing in control ofPhytophthoraroot rot of soybean[J].Plant Disease,1985,(69):362-368.
[3] Wrather JA,Stienstra WC,Koenning SR.Soybean disease loss estimates for the United States from 1996 to 1998[J].Canadian Journal of Plant Pathology,2001,3:122-131.
[4] 沈崇堯,蘇顏純.中國(guó)大豆疫霉根腐病的發(fā)現(xiàn)及初步研究[J].植物病理學(xué)報(bào),1991,21(4):298.
[5] 周肇慧,蘇顏純.大豆疫病的檢疫研究一病原菌的分離鑒定[J].植物檢疫,1995,(5):257-261.
[6] 李寶英,馬淑梅.大豆疫霉病研究初報(bào)[J].大豆科學(xué),1996,15(2):164-165.
[7] 馬書君.黑龍江省大豆疫霉根腐病發(fā)生發(fā)生情況調(diào)查[J].大豆科學(xué),1997,16(1):88-89.
[8] 王曉鳴,Schmitthenner AF,馬書君.黑龍江省大豆疫霉根腐病調(diào)查與病原分離[J].植物保護(hù),1998,(3):9-11.
[9] Xiaoli Liu,Mingsheng Dong,Xiaohong Chen.Antimicrobial activity of an endophyticXylariasp.YX-28 and identification of its antimicrobial compound 7-amino-4-methylcoumarin[J].Applied Microbiology &Biotechnology,2008,78:241-247.
[10] Latijnhouwers M,de Wit PJ,Govers F.Oomycetes and fungi:similar weaponry to attack plants[J].Trends in Microbiology,2003,11(10):462-469.
[11] 況福元,吳小麗,呂風(fēng)青.菜心炭疽病菌拮抗細(xì)菌的篩選及鑒定.微生物學(xué)通報(bào)[J].2009,(9):1350-1355.
[12] 麥明曉,黃惠琴,葉建軍.香蕉枯萎病拮抗放線菌Da08006的篩選與鑒定[J].生物技術(shù)通報(bào),2009,(S1):169-171.
[13] 章四平,劉圣明,王建新.枯草芽孢桿菌生防菌株NJ 18的發(fā)酵條件優(yōu)化[J].南京農(nóng)業(yè)大學(xué)學(xué)報(bào),2010,33(2):58-62.
[14] 孫力軍,陸兆新.培養(yǎng)基對(duì)解淀粉芽孢桿菌ES-2菌株產(chǎn)抗
菌脂肽的影響[J].中國(guó)農(nóng)業(yè)科學(xué)2008,41(10):3389-3398.
Screening of Liquid Medium and Optimization of Fermentation Conditions for a Bacterium B048 against Phytophthora Sojae
WANG Wan, ZHAO Ping-sheng, ZHOU Tao, WANG Zi-ying
(Departmentoflifescience,HefeiNormalUniversity,Hefei230061,China)
Single factor test and orthogonal chart were used to optimize the fermentation medium of a bacterium B048.The results indicated that the medium was corn flour 5 g/L,soybean meal 15 g/L,sodium chloride 10 g/L.On the basis of the medium components,the culture conditions were optimized.The results show that the initial pH value is 6,the incubation time is 48 h,and the rotational speed is 180 r/min.
Phytophthorasojae;B048;culture medium;fermentation
Q939.96
A
1674-2273(2011)06-0079-04
2011-06-20
國(guó)家自然科學(xué)基金項(xiàng)目(30800040);安徽省優(yōu)秀青年科技基金項(xiàng)目(10040606Y04);教育部留學(xué)回國(guó)人員科研啟動(dòng)基金項(xiàng)目
汪莞(1990-),女,安徽合肥人,合肥師范學(xué)院生命科學(xué)系08生物科學(xué)專業(yè)學(xué)生;通訊聯(lián)系人:王子迎(1976-),安徽靈璧縣人,教授,博士,研究方向:真菌病害控制。
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國(guó)化肥信息(2022年7期)2022-08-31 01:29:28
中國(guó)化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場(chǎng)周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報(bào)·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
