星形膠質(zhì)細胞和神經(jīng)元之間谷氨酰胺的轉(zhuǎn)運研究現(xiàn)狀
2011-11-17 08:10:16王紅王栓科汪靜
中國科技信息 2011年13期
關(guān)鍵詞:系統(tǒng)
王紅 王栓科 汪靜
蘭州大學第二醫(yī)院骨科研究所,蘭州 730000
星形膠質(zhì)細胞和神經(jīng)元之間谷氨酰胺的轉(zhuǎn)運研究現(xiàn)狀
王紅 王栓科 汪靜
蘭州大學第二醫(yī)院骨科研究所,蘭州 730000
從谷氨酰胺的星形膠質(zhì)細胞排除到神經(jīng)元對其進行攝取,谷氨酸-谷氨酰胺循環(huán)是必不可少的一步,這也是神經(jīng)元補充谷氨酸的主要途徑。本文主要是對參與這一過程的谷氨酰胺轉(zhuǎn)運體的類別和功能做一綜述。體內(nèi)大多數(shù)參與谷氨酰胺轉(zhuǎn)運的轉(zhuǎn)運體是相互關(guān)聯(lián)的,許多屬性都是相同的,如都對微小p H值變換很敏感等。介導成人大腦中谷氨酰胺轉(zhuǎn)運的轉(zhuǎn)運系統(tǒng)主要是N轉(zhuǎn)運系統(tǒng)(S N1)和A轉(zhuǎn)運系統(tǒng)(S A T/A T A),它們都是AAAP轉(zhuǎn)運家族的成員。它們之間主要的區(qū)別就是生理條件下S N1對谷氨酰胺轉(zhuǎn)運是可逆的,這使得S N1既可以維持星形膠質(zhì)細胞內(nèi)谷氨酰胺的濃度梯度,也可介導谷氨酰胺的凈外流。但在增值的星形膠質(zhì)細胞內(nèi),S N1的功能被A S C T2取代。
神經(jīng)膠質(zhì)細胞;神經(jīng)元;谷氨酸-谷氨酰胺循環(huán);A轉(zhuǎn)運系統(tǒng);N轉(zhuǎn)運系統(tǒng)
astrocyte; neuron; glutamate-glutamine cycle; system A transport; system N transport
谷氨酸是大腦興奮性突觸傳遞的重要神經(jīng)遞質(zhì),釋放到突觸間隙的谷氨酸被星形膠質(zhì)細胞重攝取后被酰胺化為一種非興奮性神經(jīng)遞質(zhì)谷氨酰胺,谷氨酰胺被釋放到細胞外間隙,被神經(jīng)元重攝取后轉(zhuǎn)化為谷氨酸[1,2]。這種將谷氨酰胺從星形膠質(zhì)細胞轉(zhuǎn)移到神經(jīng)元的過程就是大家熟知的谷氨酸-谷氨酰胺循環(huán)。但在此過程中谷氨酰胺的轉(zhuǎn)移到底是如何被介導和調(diào)控的直到現(xiàn)在還未達成共識,且谷氨酰胺還和其他許多中性氨基酸共用轉(zhuǎn)運通道。
目前,對它們的研究有了很大的進展。首先,在星形膠質(zhì)細胞和神經(jīng)元內(nèi)的谷氨酰胺轉(zhuǎn)運的功能性研究已經(jīng)定位到特定的轉(zhuǎn)運體上。其次,對氨基酸轉(zhuǎn)運蛋白新家族鑒定的突飛猛進,使我們可以單獨的對這些谷氨酰胺轉(zhuǎn)運體進行研究。本綜述主要是對參與谷氨酰胺在細胞間轉(zhuǎn)運的轉(zhuǎn)運體的最新進展進行闡述。
1. 谷氨酸-谷氨酰胺循環(huán)
星形膠質(zhì)細胞和神經(jīng)元是代謝偶聯(lián)的,其中最重要的途徑之一是谷氨酸-谷氨酰胺循環(huán)。研究表明,谷氨酸-谷氨酰胺循環(huán)并不是完全封閉的[3]。因為谷氨酸不能穿過血腦屏障,所以中樞神經(jīng)系統(tǒng)(C N S)中谷氨酸是葡萄糖經(jīng)糖酵解和三羧酸循環(huán),由三羧酸循環(huán)的中間物合成的。在此過程中需丙酮酸羧化酶的作用,但該酶在神經(jīng)元中不存在,而大量存在于星形膠質(zhì)細胞中并有功能性活性。谷氨酰胺的合成是谷氨酸-谷氨酰胺循環(huán)的重要部分。合成反應由谷氨酰胺合成酶催化,該酶只存在于星形膠質(zhì)細胞,在少突膠質(zhì)細胞中很少,神經(jīng)元中不存在,因此它是星形膠質(zhì)細胞的標志性酶。星形膠質(zhì)細胞迅速攝取細胞外的谷氨酸[4],谷氨酸載體和谷氨酰胺合成酶在星形膠質(zhì)細胞中的重疊分布表明,在突觸位置攝取釋放的谷氨酸和在星形膠質(zhì)細胞中轉(zhuǎn)變?yōu)楣劝滨0肥蔷o密聯(lián)系的[5]。谷氨酰胺從星形膠質(zhì)細胞釋放作為谷氨酸的前體在神經(jīng)元中積累,在突觸前末端轉(zhuǎn)變成谷氨酸,進入小泡準備新一輪的興奮反應。谷氨酸前體的返回保持了谷氨酸能神經(jīng)元的谷氨酸水平。由于谷氨酸不能穿過血腦屏障,所以多余的谷氨酸必須通過T A C氧化降解后再從C N S 中排出,多余的谷氨酰胺經(jīng)水解變成谷氨酸后也以相同的途徑氧化降解。
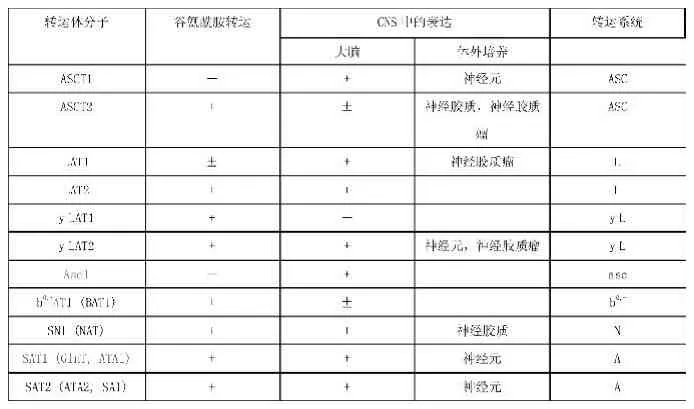
表1
2. 中性氨基酸轉(zhuǎn)運體
谷氨酰胺轉(zhuǎn)運體家族主要分為兩類:鈉離子依賴性和非鈉離子依賴性。目前在中樞神經(jīng)系統(tǒng)中已發(fā)現(xiàn)7個谷氨酰胺轉(zhuǎn)運體家族,鈉離子依賴性者包括s y s t e m A、s y s t e m N、s y s t e m y(+) L及A S C,s y s t e m B,非鈉離子依賴性者包括s y s t e m L、s y s t e m b。前者主要依靠N a+-K+-A T P酶提供能量,進行膜內(nèi)外谷氨酰胺的轉(zhuǎn)運;后者轉(zhuǎn)運谷氨酰胺不需要N a+提供能量[6]。近幾年來,通過基因研究,獲得幾種編碼轉(zhuǎn)運蛋白的c D N A,并證實谷氨酰胺載體蛋白系統(tǒng)分屬于4個不同的基因家族。如N a+依賴谷氨酰胺載體蛋白基因有:系統(tǒng)N(S N l)、系統(tǒng)A(A T A l,A T A2),系統(tǒng)A S C/B0 (A S C T2或A T B0),系統(tǒng)B0,+(A T B0,+)和系統(tǒng)y+L(y+L A T1,y+L A T2)。非N a+依賴谷氨酰胺載體蛋白基因編碼系統(tǒng)L(L A T1,L A T2)和系統(tǒng)b0,+(b0,+A T),其中轉(zhuǎn)運二性離子氨基酸用“0”上標,轉(zhuǎn)運陽離子氨基酸用“+”上標[7]。具體見表[8]。
3.N和A轉(zhuǎn)運系統(tǒng)的谷氨酰胺轉(zhuǎn)運
在星形膠質(zhì)細胞和神經(jīng)元中,轉(zhuǎn)運系統(tǒng)N和A對于谷氨酰胺的轉(zhuǎn)運作用最重要。培養(yǎng)的小鼠星形膠質(zhì)細胞中的谷氨酰胺轉(zhuǎn)運分析結(jié)果[9]顯示,N系統(tǒng)是谷氨酰胺攝取的主要通路。在腦組織,S N1 m R N A和載體蛋白僅限于膠質(zhì)細胞,S N1位于神經(jīng)元細胞。通過研究肝腺泡發(fā)現(xiàn),末梢靜脈周圍的肝細胞質(zhì)膜富含S N1載體蛋白及谷氨酰胺合成酶,并且很活躍。S N1介導谷氨酰胺攝取的 K m值大約是1.1±1.6 m m,說明S N1對谷氨酰胺攝取的過程不受膜去極化的影響。這種H+交換必將導致p H值的變化,這與谷氨酰胺攝取的p H依賴是一致的[10]。 更重要的是在生理水平下谷氨酰胺/H+交換是可逆的。神經(jīng)元活動增加將降低星形膠質(zhì)細胞內(nèi)N a+和H+濃度梯度,最終通過S N1轉(zhuǎn)運導致谷氨酰胺外流,使得星形膠質(zhì)細胞內(nèi)谷氨酰胺的重新分布[9]。
上述星形膠質(zhì)細胞對谷氨酰胺的轉(zhuǎn)運主要是通過S N1,但是在神經(jīng)元主要是通過A系統(tǒng)。在過量的M e A I B不能抑制原代培養(yǎng)小鼠星形膠質(zhì)細胞谷氨酰胺的吸收,說明其缺乏A系統(tǒng)介導的攝取。鼠S A T1/A T A1主要分布于神經(jīng)元,在大腦和脊髓廣泛表達[11],在突觸小泡很少。原代培養(yǎng)大鼠小腦顆粒細胞中有S A T1的/ A T A1表達,但星形膠質(zhì)細胞中卻沒有。人A T A1 m R N A(9 k b)僅在胎盤和心臟有表達。在腦組織,無論鼠A T A1及A T A2均限于神經(jīng)元,膠質(zhì)細胞中不存在,與S N1、A S C T2/A T B0分布不同,后者在腦組織中僅表達在膠質(zhì)細胞中。
4. p H值對N和A轉(zhuǎn)運系統(tǒng)的影響
神經(jīng)元的電活動使得星形膠質(zhì)細胞H+、N a+和跨膜電位梯度下降,促使谷氨酰胺通過S N1[9]排除。活躍神經(jīng)元釋放K+使星形膠質(zhì)細胞發(fā)生去極化,從而加速了電依賴的N a+- H CO3-同向轉(zhuǎn)運體對H CO3
-的轉(zhuǎn)運。這將使得星形膠質(zhì)細胞內(nèi)部發(fā)生堿化,并促使谷氨酸-谷氨酰胺循環(huán)大量激活,從而加速了谷氨酸的攝取以及谷氨酰胺的合成[12]。堿化過程必然伴隨著對應的細胞外酸化,導致跨膜H+梯度下降或反轉(zhuǎn),促進S N1介導的谷氨酰胺外流交換H+。接下來我們還需進一步對S N1轉(zhuǎn)運體的能學進行研究,以便于我們更好模擬細胞活動變化對電化學梯度的影響,以及對谷氨酰胺外流的作用。相比于S N1,沒有跡象表明p H值變化可通過S A T1/A T A1或S A T2/A T A2扭轉(zhuǎn)谷氨酰胺轉(zhuǎn)。S A T/A T A轉(zhuǎn)運體主要的功能是在細胞質(zhì)內(nèi)儲積谷氨酰胺,尤其是在神經(jīng)元。然而,當p H值從7.4下調(diào)到7.0時,S A T1/A T A1或S A T2/A T A2轉(zhuǎn)運速率下降一半,但當上調(diào)到7.8時,速率明顯加快,將近翻了一倍[11]。 這種酸堿度敏感性的機制尚不太清楚。對系統(tǒng)A的功能研究發(fā)現(xiàn)[13],細胞內(nèi)和細胞外p H改變也可導致類似情況發(fā)生,表明是依賴p H感受器調(diào)節(jié)的,而不是H+的濃度梯度。到目前為止,表達性研究還是證明不了是通過S A T/A T A轉(zhuǎn)運體導致的H+轉(zhuǎn)移。
5. 小結(jié)
在成人腦組織中,星形膠質(zhì)細胞和神經(jīng)元之間谷氨酰胺轉(zhuǎn)移主要是由N轉(zhuǎn)運系統(tǒng)(S N1)和A轉(zhuǎn)運系統(tǒng)(S A T/ A T A)介導的,它們都是A A A P轉(zhuǎn)運家族的成員,在形態(tài)和功能上是相聯(lián)系的,如在生理范圍內(nèi),都對微小的p H變化敏感等。兩者主要的區(qū)別在于在生理條件下,S N1對谷氨酰胺的內(nèi)流轉(zhuǎn)運表現(xiàn)為穩(wěn)定的可逆性,這可維持星形膠質(zhì)細胞內(nèi)的谷氨酰胺濃度梯度,且可調(diào)節(jié)谷氨酰胺的凈外流。雖然在過去的十幾年中研究者對于星形膠質(zhì)細胞和神經(jīng)元之間谷氨酰胺轉(zhuǎn)運的研究取得了一定的成就,但要進一步了解其機制,仍有很長的一段路要走。
[1] Hertz L., Dringen R., Schousboe A. andRobinson S. R. (1999) Astrocytes: Glutamateproducers for neurons. J. Neurosci. Res. 57,417-428
[2] Magistretti P. J., Pellerin L., Rothman D.L. and Shulman R. G. (1999)Energy ondemand. Science 283, 496-497.
[3] Hertz L;Yu ACH;Kala G Neuronal-astrocyticand cytosolic-mitochondrial metabolite traffickingduring brain activation, hyperammonemia andenergy deprivation
[4] Suchak SK;Baloyianni NV;Perkinton MS The'glial' glutamate transporter, EAAT2 (Glt-1)accounts for high affinity glutamate uptake intoadult rodent nerve endings
[5] Ventura R.Harris KM Three-dimensionalrelationships between hippocampal synapses andastrocytes 1999
[6] Bode BP.Recent moIecuIar advances inmammalian gIutamine transport.J Nutr,2001,131(9 Suppl):2475S-2485S
[7] 周濟宏.谷氨酰胺轉(zhuǎn)運載體的種類與分布及功能特點.醫(yī)學研究生學報.第20 卷第4期,2007 年4 月
[8] Stefan Bro è er and Neville Brookes,Transfer of glutamine between astrocytes and Neurons,Journal of Neurochemistry,2001,77,705-719
[9] Chaudhry F. A., Reimer R. J., Krizaj D.,Barber D., Storm-Mathisen J.,Copenhagen D. R.and Edwards R. H. (1999) Molecular analysis of System N suggests novel physiological roles in nitrogen metabolism and synaptic transmission.Cell99,769-780
[10] Nagaraja T. N. and Brookes N. (1996)Glutamine transport in mouse cerebral astrocytes.J. Neurochem.66,166-1674
[11] Varoqui H., Zhu H., Yao D., Ming H. and Erickson J. D. (2000) Cloning and functional identication of a neuronal glutamine transporter.J. Biol. Chem.275,4049-4054
[12] Brookes N. (2000) Functional integration of the transport of ammonium, glutamate and glutamine in astrocytes. Neurochem. Int.37,121-129
[13] Yao D., Mackenzie B., Ming H., Varoqui H., Zhu H., Hediger M. A. and Erickson J. D.(2000) A novel system A isoform mediating Na1/neutral amino acid cotransport. J. Biol.Chem.275,22790-22797
Research Status of the Transfer of glutamine between astrocytes and neurons
Wang hong Wang shuan-ke Wang jing
Institute of Orthopedics,Lanzhou University Second Hospital,Lanzhou 730000,China
The export of glutamine from astrocytes, and the uptake of glutamine by neurons, are integral steps in the glutamate-glutamine cycle, a major pathway for the replenishment of neuronal glutamate. We review here the functional and classification of the transporters that mediate this transfer. The participating glutamine transporters are functionally and structurally related, sharing the following properties,such as they are sensitive to small pH changes in the physiological range. The emerging picture of glutamine transfer in adult brain is of a dominant pathway mediated by system N transport (SN1) in astrocytes and system A transport (SAT/ATA) in neurons, they are members of the AAAP family of transporters. A key difference between SN1 and the SAT/ATA transporters is the reversibility of glutamine fluxes via SN1 under physiological conditions, which allows SN1 both to sustain a glutamine concentration gradient in astrocytes and to mediate the net outward flux of glutamine. It is likely that the ASCT2 transporter displaces the SN1 transporter as the main carrier of glutamine export in proliferating astrocytes.
10.3969/j.issn.1001-8972.2011.13.078
猜你喜歡
工業(yè)設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術(shù)與產(chǎn)品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(shù)(2019年12期)2019-12-25 03:06:46
制造技術(shù)與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(yè)(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(shù)(2017年9期)2017-09-26 03:41:45
知識經(jīng)濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(shù)(2016年6期)2016-04-20 06:21:32
