A phylogeny of the Tylototriton asperrimus group (Caudata: Salamandridae) based on a mitochondrial study: suggestions for a taxonomic revision
2011-12-25 06:40:50YUANZhiYongJIANGKeShunQingYANGJunXiaoNGUYENQuangTruongNGUYENThienTaoJINJieQiongCHEJing
Zoological Research 2011年6期
關鍵詞:物種
YUAN Zhi-Yong, JIANG Ke, Lü Shun-Qing, YANG Jun-Xiao, NGUYEN Quang Truong, NGUYEN Thien Tao, JIN Jie-Qiong, CHE Jing,
(1. State Key Laboratory of Genetic Resources and Evolution, Kunming Institute of Zoology, the Chinese Academy of Sciences, Kunming Yunnan 650223, China; 2. College of Life and Environment Sciences, Huangshan University, Huangshan 245021, China; 3. Institute of Ecology and Biological Resources, 18 Hoang Quoc Viet St., Hanoi, Vietnam; 4. Vietnam National Museum of Nature, 18 Hoang Quoc Viet St., Hanoi, Vietnam)
A phylogeny of theTylototriton asperrimusgroup (Caudata: Salamandridae) based on a mitochondrial study: suggestions for a taxonomic revision
YUAN Zhi-Yong1, JIANG Ke1, Lü Shun-Qing2, YANG Jun-Xiao1, NGUYEN Quang Truong3, NGUYEN Thien Tao4, JIN Jie-Qiong1, CHE Jing1,*
(1. State Key Laboratory of Genetic Resources and Evolution, Kunming Institute of Zoology, the Chinese Academy of Sciences, Kunming Yunnan 650223, China; 2. College of Life and Environment Sciences, Huangshan University, Huangshan 245021, China; 3. Institute of Ecology and Biological Resources, 18 Hoang Quoc Viet St., Hanoi, Vietnam; 4. Vietnam National Museum of Nature, 18 Hoang Quoc Viet St., Hanoi, Vietnam)
A phylogenetic hypothesis for the Asian newts of theTylototriton asperrimusgroup was generated using data from two mitochondrial fragments including COI and the ND1-ND2 regions. Four distinct clades (A, B, C, D) were resolved with high nodal support within this monophyletic group. Clade A includedT. asperrimus,T.hainanensis,T. notialis, “T. vietnamensis”, and two unnamed salamander populations from Vietnam. Clade A, constituted the sister group of clades B + C. Newly identified clade C likely represents a new cryptic species. Clade C was the sister group ofT. wenxianensis. The trueT. vietnamensisexclusively constituted clade D. Our results bring into question some previous taxonomic decisions, and a revision is required. This study illustrates the necessity to include samples from type localities in taxonomic studies, and highlights the importance of fine-grained geographical sampling.
Tylototriton notialis;Tylototriton hainanensis; Salamander; Southeast Asia; Tonkin; Cryptic diversity
The Asian newt genusTylototritonAnderson, 1871 occurs from Nepal, through north-central India, Sikkim, Myanmar, northern Thailand, Laos, and Vietnam into southern China. Presently it contains nine species (AmphibiaWeb, 2011). The monophyly of the genus and two groups (or subgenera: Dubois & Raffa?lli, 2009), theT. verrucosusgroup andT. asperrimusgroup, have been consistently supported in recent studies (Stuart et al, 2010; Weisrock et al, 2006).
With the addition of two newly described species,T. vietnamensisB?hme et al, 2005 andT. notialisStuart et al, 2010, theT. asperrimusgroup includes five species in total, along withT. asperrimusUnterstein, 1930,T. hainanensisFei et al, 1984, andT. wenxianensisFei et al, 1984. However, despite recent progress, uncovering the dimensions of amphibian diversity and inferring evolutionary history remain difficult, in part because of extreme morphological conservation and associated homoplasy (Bossuyt & Milinkovitch, 2000; Wake, 1991). Identifying species is challenging for theT. asperrimusgroup, and determining the exact distributions of some species has been confusing (Fei et al, 2005, 2009; Nguyen et al, 2009), for example,T. asperrimus,T. wenxianensis, andT. vietnamensis. Moreover, previous molecular phylogenetic studies on this group are chiefly based on limited samples and at the same time, unfortunately, materials from the type localities are often absent (Stuart et al, 2010; Weisrock et al, 2006).
Here we report results of our use of mitochondrial data to investigate the phylogenetic relationships ofTylototriton,with an emphasis on understanding the evolutionary history of theT. asperrimusgroup.
1 Material and Methods
1.1 Sampling
Tissue samples from adults, including toe tips, muscle, and liver, and whole juvenile specimen were collected in the field following Animal Use Protocols approved by the Kunming Institute of Zoology Animal Care and Ethics Committee. The tissues were preserved in 95% ethanol. A total of 36 individuals ofTylototritonfrom 17 localities, representing all 9 currently recognized species in this genus, were examined in this study. Nineteen sequences were downloaded from GenBank for certain comparisons and validations. Outgroup taxa wereEchinotriton andersoniandE. chinhaiensis,chosen mainly based on the study of Weisrock et al (2006). Fig. 1 provides an overview on the distribution of our samples. Details of taxonomic sampling and GenBank accession numbers are provided in Tab. 1. Species identity was based on morphological characters determined in the field. Some specimens could not be reliably identified to species and we refer to them as“sp.”.
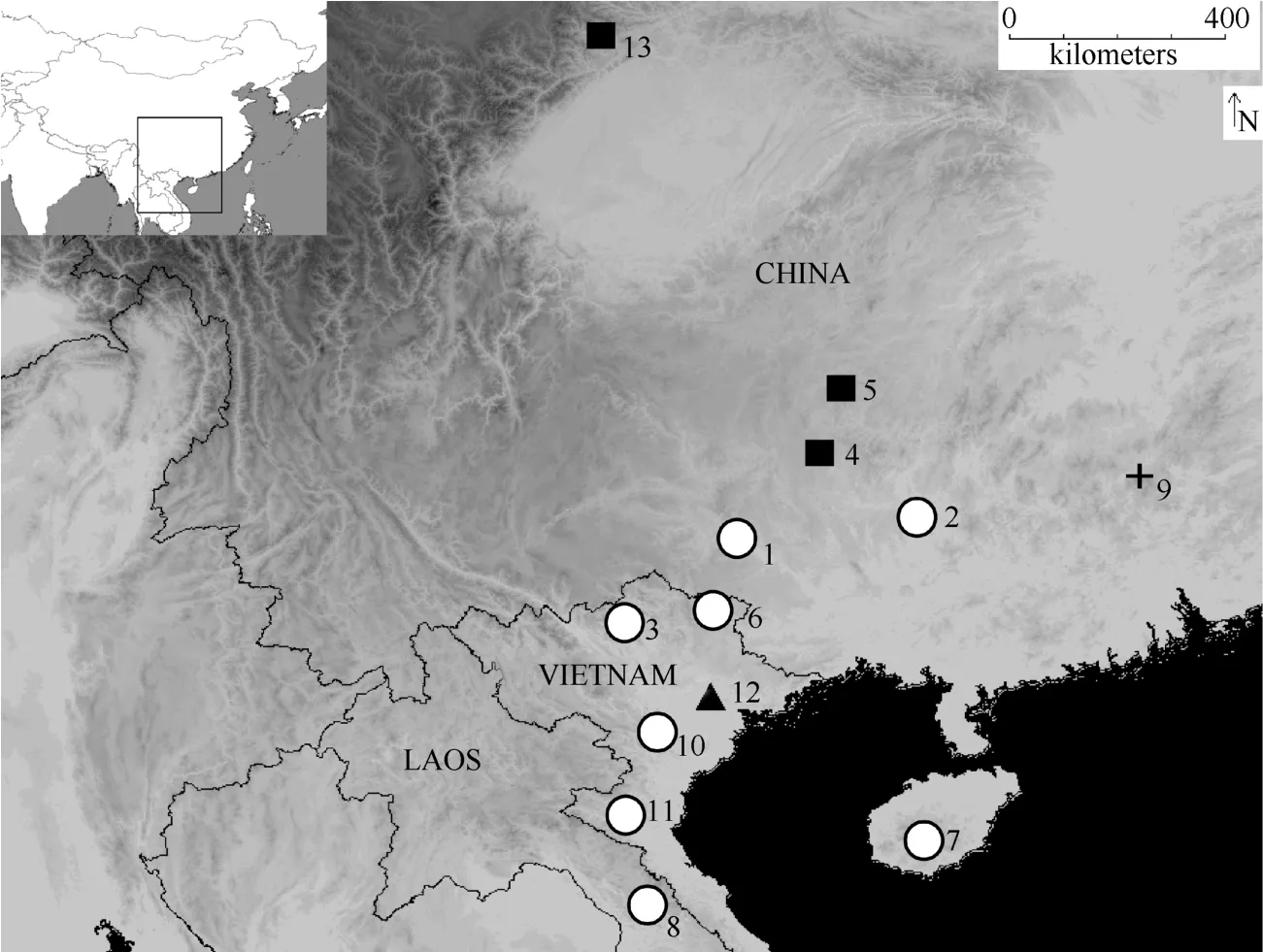
Fig. 1 Map showing the geographic locations of samples representing the Tytolotriton asperrimus group used in present analyses
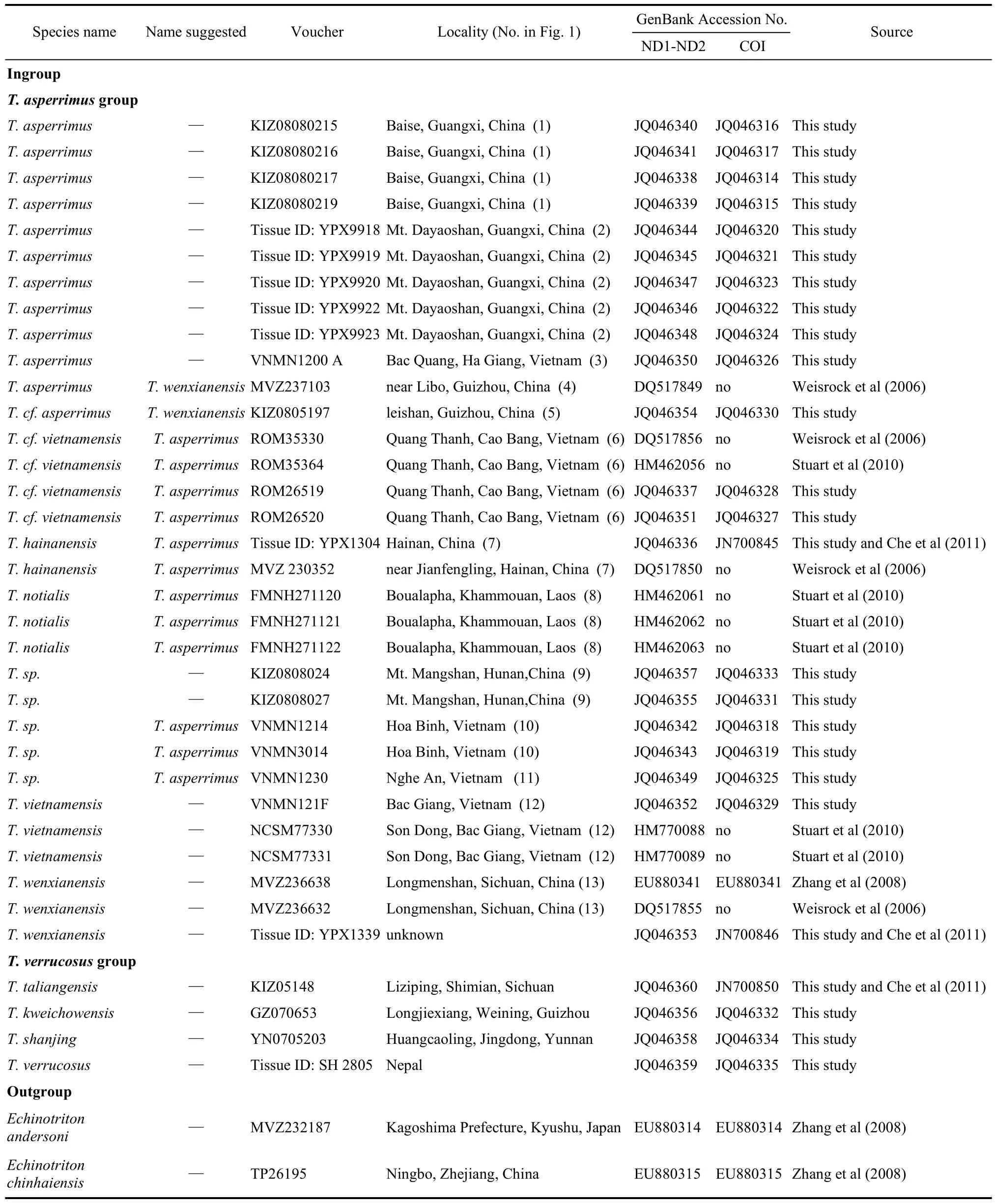
Tab. 1 Voucher specimens, localities, and GenBank accession numbers for sample used for the Tylototriton
1.2 Extraction, amplification and sequencing
Genomic DNA was extracted using the phenol-chloroform method (Sambrook et al, 1989). Two mitochondrial sequences were amplified. One fragment covers a 1 014 bp region that encodes part of the NADH dehydrogenase subunit 1, tRNA Ile, tRNA Gln, tRNA Met, and part of the NADH dehydrogenase subunit 2 genes (hereafter, ‘ND1 to ND2’). PCR primers KIZ1S (5'-TGACCAATAGCARCAATA-3') and KIZ1A (5'-GTGGGCRATAGATGARTA-3') were designed specifically for successful amplification. Internal primers KIZL4437 (5'-AAGCTTTCGGGCCCATACC-3') and KIZH4419 (5'-AAGCTTTTGGGCCCATACC-3') were applied to sequencing. Another gene is a 561 bp region from cytochrome oxidase subunit I (COI) which was amplified following the standard protocol (Che et al, in press). Amplification was performed in a 25 μL volume reaction with the following procedures: initial denaturation step for 5 min at 94 °C, 35 cycles of denaturation for 1 min at 94 °C, annealing for 1 min at 59 °C for ND1 to ND2 and 45 °C for COI, extension for 1 min at 72 °C. Final extension at 72 °C was conducted for 10 min.
PCR products were purified with a Gel Extraction Mini Kit (Watson BioTechnologies, Shanghai), and sequenced in both directions using a BigDye Terminator Cycle Sequencing Kit (version 2.0, Applied Biosystems) and a ABI PRISM 3730 automated DNA sequencer (Applied Biosystems). The sequence data were translated into amino acids to confirm the absence of premature stop codons, i.e. to preclude the sequenced of nuclear DNA pseudogenes, and also submitted to a BLAST search in GenBank to confirm that the required sequences had been amplified. Nucleotide sequences were aligned using Clustal X 1.81 (Thompson et al, 1997) with default parameters, and then optimized by eye in MEGA 4.0. (Tamura et al, 2007)
1.3 Molecular analyses
Considering that all mtDNA gene sequences are virtually inherited as one linkage group, the two mtDNA gene segments were concatenated into a single partition. The ten samples for which COI sequences are lacking were treated as missing data when combined. Mixedmodel Bayesian phylogenetic inference was performed using Bayesian inference (BI) with MrBayes version 3.1.2 (Ronquist & Huelsenbeck, 2003). Data were partitioned by ND1 codon position, tRNA, ND2 codon position, and COI codon position. The model of sequence evolution that best described each of the data partitions was inferred using the Akaike Information Criterion as implemented in MrModeltest 2.3 (Posada & Crandall, 1998). These were TIM+G for ND1 first codon position; HKY+I for ND1 second codon position; GTR+I for ND1 third codon position and tRNA; TRN+G for ND2 first codon position; HKY+G for ND2 second codon position; GTR+G for ND2 third codon position and COI third codon position; and F81 for COI first and second codon positions. Four independent Bayesian analyses were performed on each dataset. In each analysis, four chains were run for 5 000 000 generations using the default priors, trees were sampled every 100 generations, log-likelihood scores were tracked for stabilization and the first 25% of the trees were discarded as burn-in.
2 Results and Discussion
2.1 Molecular analyses
The aligned sequences yielded 1 014 base pairs of ND1 to ND2 (305 variable, 233 parsimony-informative sites) and 561 bp base pairs of COI (169 variable, 133 parsimony-informative sites). The expected bias against guanine nucleotides was observed (G=12.0%, A= 35.1%, T=25.6 %, and C=27.3 % for ND1 to ND2; and A=25.9%, T=27.4%, C=29.4%, G=17.3% for COI). These data indicated the mitochondrial gene was sequenced, and not a nuclear copy. Forty-seven new sequences were determined and deposited in GenBank (Tab. 1).
The BI tree is shown in Fig. 2. Consistent with Weisrock et al (2006) and Stuart et al (2010), the monophly of the genus ofTylototritonis strongly supported. A major difference is that our data did not recover the two major clades withinTylototritonreported in previous studies. This is not surprising given our sequencing of a shorter fragment and our far denser sampling forT. asperrimusgroup. The monophyly of theT. asperimmusgroup as shown by Stuart et al (2010) collapsed into two subclades (A+B+C and D) in the present analyses. Clade A including seven strongly supported subclades (A1?A7) was recovered as the sister group of B+C. The position of clade D was not resolved in the present study.
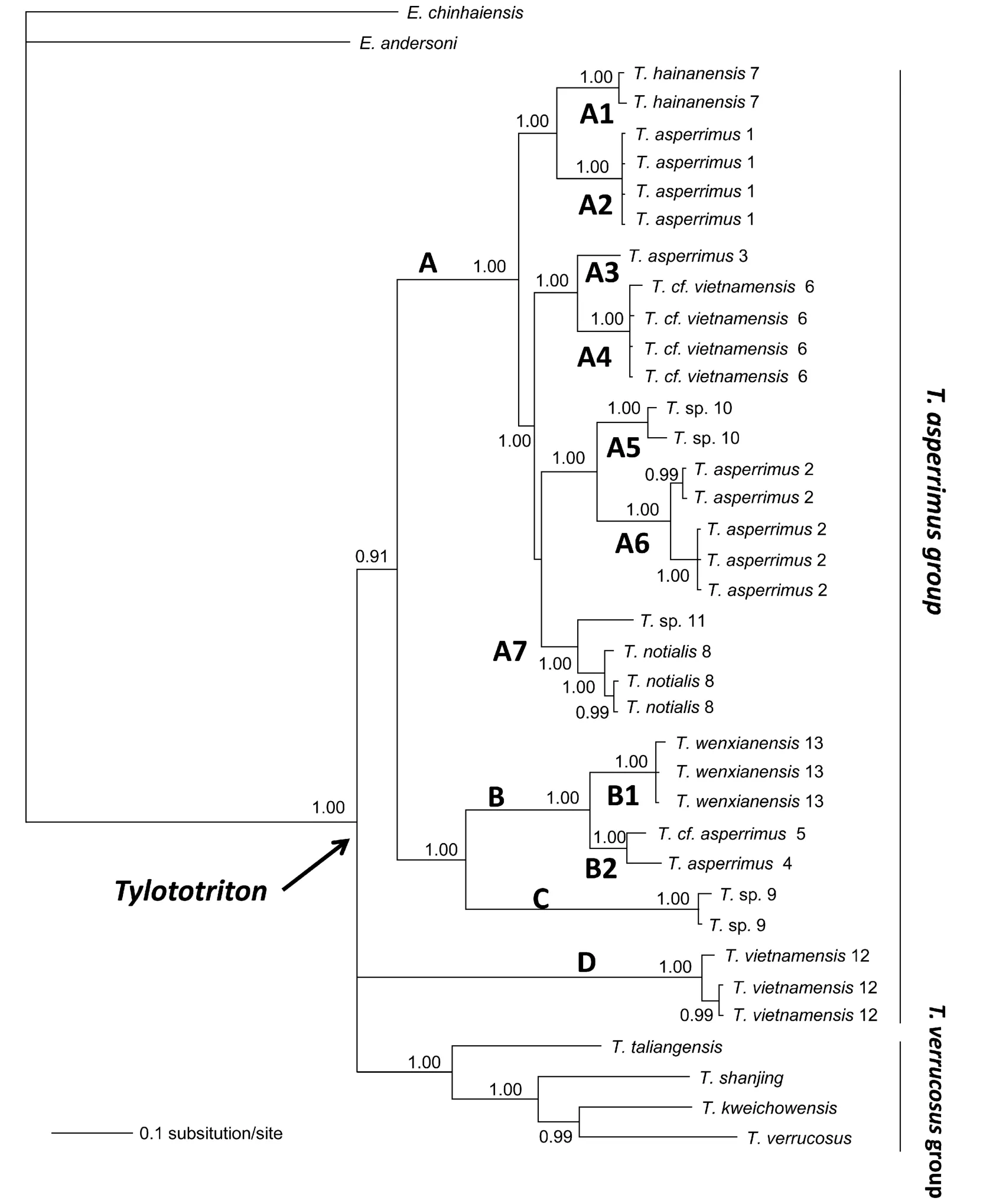
Fig. 2 A Bayesian tree resulting from mixed-model analysis of mitochondrial DNA
Our increased sampling reveals previously unrecorded phylogenetic structure. On the one hand,T. asperrimusdoes not contain a single maternal lineage (Fig. 2). The nominal form ofT. asperrimus(type locality: Dayao shan, Guangxi; locality 2) is in lineage A6, which is the sister taxon to the two apparently unnamed samplesT.sp. from Hoa Binh, Vietnam (A5). Lineages A2, A3, and B2 have also been assigned toT. asperrimus. Salamanders from Baise, Guangxi (locality 1) form lineage A2, which is closely related toT. hainanensis(lineage A1). Lineage A3 is from Ha Giang, Vietnam (locality 3), and it is the sister group of “T. vietnamensis” from Cao Bang, Vietnam (locality 6). Lineage B2 is from Libo and Leigongshan, Guizhou, China (localities 4 and 5), is the sister group ofT. wenxianensis(lineage B1). Additionally, the trueT.vietnamensisfrom Bac Giang, Vietnam, exclusively constitutes clade D, which is distantly related to the population from Cao Bang (locality 6).
On the other hand, compared with previous studies, our data unexpectedly identify a distinct evolutionary clade C from Mangshan, Hunan (locality 9), which is the sister group of clade B.
2.2 Taxonomic implications
Species identification within theT. asperrimusgroup is challenging because all members are superficially very similar and conservative in morphology (Stuart et al, 2010). Except forT. hainanensisandT. notialis, which occur in limited geographic regions, Hainan Island and Laos, respectively,T. wenxianensis,T. asperrimus, andT. vietnamensishave more dispersed distributions (Chen, 1991; Dai et al, 2011; Fei et al, 2005, 2009; Nguyen et al, 2009) and species borders are not clear.
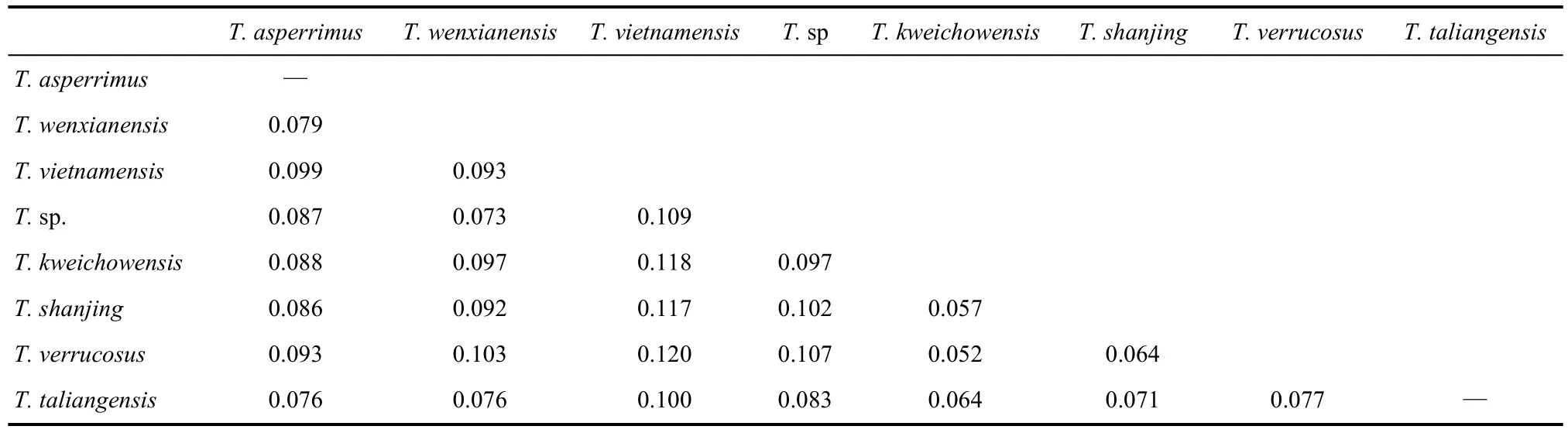
Tylototriton wenxianensiswas reported to occur in Gansu, Sichuan, Chongqing, southeastern Guizhou, Anhui, and nothern and central Hunan provinces in China (Fei et al, 2005, 2009). Our limited samples supported the monophyly ofT. wenxianensisas a separate species, which can be interpreted as support for the taxonomic revision by Fei et al (1984). Moreover, we consider two samples from southeastern and extremely southern Guizhou, originally identified as “T. asperimmus” by Weisrock et al (2006) and our team,to be conspecific withT. wenxianensis,based on the phylogeny and genetic distance estimation. Among these, one sample from Guizhou (locality 4) was unfortunately used to represent the nominalT. asperimmusin comparisons of genetic distances with other species, for example,T. notialis(Stuart et al, 2010). Based on our data, we refer clade B to beT. wenxianensis(Tab. 1, Fig. 3). The level of genetic divergence betweenT. wenxianensis(B) and other clades is relatively higher (0.073?0.103) than other comparisons withinTylototriton(Tab. 2). Denser sampling within this clade is required, including investigation of the recent report of a population ofT. wenxianensisrelatively far to the north in Henan province, China (Chen et al, 2010).
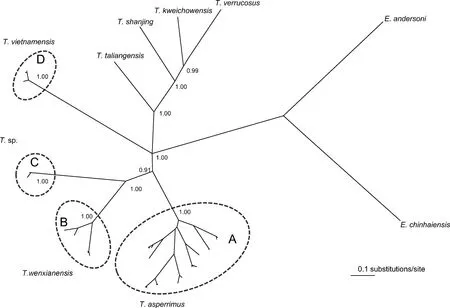
Fig. 3 Unrooted phylogenetic tree of mitochondrial gene sequences reconstructed using Bayesian methods for the TylototritonPosterior probabilities are displayed only at the major nodes. Taxonomic change suggestions were indicated on this tree.
Tab. 2 K2P pairwise distances of the ND1-ND2 sequences used in this study for the Tylototriton
Currently,T. asperrimusis reported to be distributed in Guangxi, Guangdong, northern, western, and southern Guizhou, China (Fei et al, 2005, 2009) and northern Vietnam, for example, Lao Cai and Ha Giang (Nguyen et al, 2009). The distributions ofT. asperrimusandT. vietnamensisin Vietnam require further assessments. Presently,T. vietnamensisis only reported to occur in Cao Bang, Lang Song, and Bac Giang (Nguyen et al, 2009). Based on our new molecular analyses and calculated genetic distances (Tab. 2),T. vietnamensisfrom the type locality of Bac Giang was distinctly separated as our clade D, as hypothesized by Stuart et al (2010). However, the assumed “T. vietnamensis” (orT. cf. vietnamensis: Stuart et al, 2010; Weisrock et al, 2006) from Cao Bang (A4) was distantly separated from the trueT. vietnamensis. Together,T. hainanensis,T. notialis, “T. vietnamensis”, and three unnamed salamanders from Vietnam clustered with the trueT. asperrimusfrom Daoyaoshan, China, which constituted clade A. Based on the evolutionary relationships and genetic distances, we assign all members of our clade A to one species,T. asperrimus(Tab. 1, Fig. 3). Accordingly, we assignT. notialisStuart et al, 2010 andT. hainanensisFei et al, 1984 as junior objective synonyms ofT. asperrimusUnterstein 1930.
The level of genetic divergence among A1-A7 ranged from 0.021 to 0.047 (not shown here), which is much lower than the hypothesized inter-specific divergence (0.052?0.120, Tab. 2). Samples from clade A are distributed around the Gulf of Tonkin (Fig. 1). The closure of the Tonkin Strait in the past probably allowed gene flow exchanges between populations of clade A.
Samples from clade C could be treated as a cryptic species, and the level of genetic divergence between lineages C and other clades withinTylototritonranged from 0.073?0.109; however, it also is similar toT. asperrimusin morphology and we could not easily identify it when we are in the field. Further detailed studies including a combination of various datasets should be done to ascertain whether it represents a distinct species (Hou et al, unpublished data), or, like clade B2, it should be assigned toT. wenxianensis. A denser sampling covering the entire distribution ranges regarding clade B and C is desirable.
Altogether, the description of amphibian diversity and evolutionary history remains a difficult task in some cases because of extremely conservative morphology and in other cases homoplasy (Bossuyt & Milinkovitch, 2000; Wake, 1991). In this case, molecular tools do help, however samples from type localities should be fully considered when formal taxonomic revisions and descriptions of new species are undertaken. Inclusion of additional genes, especially nuclear genes, may prove to be essential. MtDNA data alone cannot assess introgression and gene flow.
Acknowledgements:We thank HOU M, SHI HT, ORLOV NL, MURPHY RW, YAN F, ZHOU WW, DING L, and LI JT for specimen collection, or providing tissues. We thank WAKE DB and PAPENFUSS TJ for reviewing and offering helpful comments on this manuscript. Technical support from the laboratory of Prof. ZHANG YP is gratefully acknowledged.
AmphibiaWeb Information on amphibian biology and conservation. [web application]. 2011. Berkeley, California: AmphibiaWeb [DB/OL]. Available: http://amphibiaweb.org/. (Accessed: Nov 16, 2011).
Anderson J. 1871. Description of a new genus of newts from western Yunan [J].Proc Zool SocLond, 1871: 423-425.
Bossuyt F, Milinkovitch MC. 2000. Convergent adaptive radiations in Madagascan and Asian ranid frogs reveal covariation between larval and adult traits [J].Proc Natl Acad Sci USA, 97: 6585-6590. B?hme W, Schottler T, Nguyen TQ, K?hler J. 2005. A new species of salamander, genusTylototriton(Urodela: Salamandridae), from northern Vietnam [J].Salamandra, 41: 215-220.
Che J, Cheng HM, Yang JX, Jing JQ, Jiang K, Yuan ZY, Murphy RW, Zhang YP. Universal COI primers for DNA barcoding amphibians [J].Mol Ecol Resour(in press).
Chen BH. 1991. The Amphibians and Reptiles Fauna of Anhui [M]. Hefei: Publishing House of Science and Technology. (in Chinese)
Chen XH, Wang XW, Tao J. 2010. A new subspecies of genusTylototritonfrom China (Caudata: Salamandridae) [J].Acta Zootax Sin, 35: 666-670. (in Chinese).
Dai ZX, Zhang XM, Zhou Q, Liu ZS. 2011. Amphibian resources in Hubei province [J].Sichuan J Zool, 30: 144-147. (in Chinese).
Dubois A, Raffa?lli J. 2009. A new ergotaxonomy of the family Salamandridae Goldfuss, 1820 (Amphibia, Urodela) [J].Alytes, 26: 1-85.
Fei L, Ye CY, Yang R. 1984. A new species and a new subspecies of the genusTylototriton(Caudata: Salamandridae) [J].Acta Zool Sin,30: 85-91. (in Chinese).
Fei L, Ye CY, Jiang JP, Xie F. 2005. An Illustrated Key to Chinese Amphibians [M]. Chengdu: Sichuan Publishing House of Science and Technology. (in Chinese).
Fei L, Ye CY, Jiang JP. 2009. Colored Atlas of Chinese Amphibians [M]. Chengdu: Sichuan Publishing House of Science and Technology (in Chinese).
Nguyen SV, Ho CT, Nguyen TQ. 2009. Herpetofauna of Vietnam [M]. Germany: Edition Chimaira, Frankfurtam Main.
Posada D, Crandall KA. 1998. MODELTEST: testing the model of DNA substitution [J].Bioinformatics, 14: 817-818.
Ronquist F, Huelsenbeck JP. 2003. MRBAYES 3: Bayesian phylogenetic inference under mixed models [J].Bioinformatics, 19: 1572-1574.
Sambrook J, Fritsch E, Maniatis T. 1989. Molecular Cloning: A Laboratory Manual 2nd ed. [M] New York: Cold Spring Harbor Laboratory Pres.
Stuart BL, Phimmachak S, Sivongxay N, Robichaud WG. 2010. A new species in theTylototriton asperrimusgroup (Caudata: Salamandridae) from central Laos [J].Zootaxa, 2060: 59-68.
Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0 [J].Mol Biol Evol, 24: 1596-1599.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. 1997. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools [J].Nucleic Acids Res, 25: 4876-4882.
Wake DB. 1991. Homoplasy: the result of natural selection, or evidence of design limitations? [J].Am Nat, 138: 543-567.
Weisrock DW, Papenfuss TJ, Macey JR, Litvinchuk SN, Polymeni R, Ugurtas IH, Zhao EM, Jowkar H, Larson A. 2006. A molecular assessment of phylogenetic relationships and lineage accumulation rates within the family Salamandridae (Amphibia, Caudata) [J].Mol Phylogenet Evol, 41: 368-383.
Zhang P, Papenfuss TJ, Wake MH, Qu LH, Wake DB. 2008. Phylogeny and biogeography of the family Salamandridae (Amphibia: Caudata) inferred from complete mitochondrial genomes [J].Mol Phylogenet Evol, 49: 586-597.
基于線粒體基因對細痣疣螈種組系統發育關系的探討及物種修訂建議
袁智勇1, 蔣 柯1, 呂順清2, 楊軍校1, NGUYEN Quang Truong3, NGUYEN Thien Tao4, 金潔瓊1, 車 靜1,*
(1.中國科學院昆明動物研究所 遺傳資源與進化國家重點實驗室,云南 昆明650223; 2.黃山學院 生命與環境科學學院,安徽 黃山245021; 3. Institute of Ecology and Biological Resources, 18 Hoang Quoc Viet St., Hanoi, Vietnam; 4. Vietnam National Museum of Nature, 18 Hoang Quoc Viet St., Hanoi, Vietnam)
該文基于兩段線粒體基因COI 和ND1-ND2部分序列片段構建了細痣疣螈種組的系統發育關系, 結果顯示該單系群內包含了四個明顯的進化支系(A、B、C、D)。A支系與B、C支系構成姐妹群關系; A支系包含了細痣疣螈、海南疣螈、老撾疣螈和“越南疣螈”以及另外兩個來自越南未被命名的種群; C支系代表了一個新的獨立進化支, 可能是一個隱存種;C支系與B支系構成姐妹群關系。此外,來自模式產地的越南疣螈單獨構成支系D。該文結果不支持以往細痣疣螈種組部分物種的劃分, 建議對該種組進行新的分類修訂。該文強調了在分類研究中應該重視使用來自模式產地的樣品并進行廣泛的地理居群采樣。
老撾疣螈; 海南疣螈; 蠑螈; 東南亞; 東京灣; 隱存多樣性
Q959.520.9; Q349.1; Q951.3
A
0254-5853-(2011)06-0577-08
2011-11-18;接受日期:2011-11-28
袁智勇,男,碩士研究生,主要從事東亞和東南亞蠑螈科物種遺傳多樣性及演化研究
10.3724/SP.J.1141.2011.06577
date: 2011-11-18; Accepted date: 2011-11-28
s: This work was supported by grants from the Ministry of Science and Technology of China (MOST Grant 2011FY120200), the National Natural Science Foundation of China (31090100, 30870281), the Chinese Academy of Sciences (KSCX2-YW-Z-0807, KSCX2-EW-Z-2), and the Bureau of Science and Technology of Yunnan province.
*通訊作者(
),女,副研究員。Tel: +86-871-5199030; Fax: +86-871-5133185; E-mail: chej@mail.kiz.ac.cn
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
學生天地(2019年36期)2019-08-25 08:59:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30

- Zoological Research的其它文章
- New details indicated by different stainings during conjugation of ciliated protozoa Paramecium
- Histological and immunocytochemical study of deferens ducts in the Chinese rat snake (Zaocys dhumnades)
- PTEN在小鼠卵細胞和早期胚胎發育中的功能研究
- 云南新平哀牢山西黑冠長臂猿分布與群體數量
- 杭州灣及錢塘江河口水鳥群落組成、季節動態及種間相關性分析
- 上海市南匯東灘圍墾后海岸帶濕地冬春季水鳥生境選擇