杭州灣及錢塘江河口水鳥群落組成、季節動態及種間相關性分析
2011-12-25 06:39:58蔣科毅邵學新
Zoological Research 2011年6期
關鍵詞:物種
蔣科毅, 吳 明, 邵學新
(中國林科院亞熱帶林業研究所 國家林業局杭州灣濕地生態系統定位研究站, 浙江 富陽 311400)
杭州灣及錢塘江河口水鳥群落組成、季節動態及種間相關性分析
蔣科毅, 吳 明, 邵學新
(中國林科院亞熱帶林業研究所 國家林業局杭州灣濕地生態系統定位研究站,浙江 富陽311400)
2007年7月—2010年11月, 對杭州灣曹娥江口至甬江口間潮間帶及其附近海涂水庫的水鳥群落進行定期監測, 共記錄9目18科128種。其中, 候鳥119種, 占總數的93%; 而國家重點保護鳥類11種。對13種鸻鷸類和9種鴨類進行種群數量隨時間變化的種間相關性分析, 兩個類群各有21對和23對顯著相關。通過對水鳥季節動態及其遷徙模式分析, 將其遷徙過程分為6個階段, 即:4月上旬—5月中旬、7月下旬—9月下旬(旅鳥遷徙高峰期)、2月下旬—3月下旬、10月上旬—12月中旬(冬候鳥遷徙高峰期)、12月下旬—次年2月中旬(越冬期)和5月下旬—7月中旬(繁殖期)。種間吸引作用可能是影響該區域遷徙季節鸻鷸類和越冬地鴨類群落組成的重要原因, 而該區域特殊地理位置和水鳥群落構成可能是造成鸻鷸類春秋季遷徙高峰數量巨大差異的主要因素。
遷徙水鳥; 種間相關性; 遷徙模式
杭州灣為我國東部沿海錢塘江口延伸的河口灣, 其正處于東亞—澳大利亞候鳥遷徙路線上的中間位置(Wilson & Barter, 1998), 是重要的遷徙鳥類中轉停歇地和越冬地(Barter, 2002)。過去在杭州灣及錢塘江河口開展的水鳥研究多僅涉及北岸(Lu et al, 1988; Qian et al, 1985; Wang & Qian, 1988; Ge et al, 2006;而涉及杭州灣南岸的相關研究較少(Zhuge et al, 1986; Zhu & Yang, 1988; Zhu, 1989a; Zhu et al, 1989b)。
鳥類的遷徙活動受各種因素制約(D?nhardt & Lindstr?m, 2001), 種間吸引作用對鳥類遷徙活動的影響已獲得較多學者的關注(Thomson et al, 2003; Forsman & Thomson, 2008; Hromada et al, 2008; Forsman et al, 2009)。目前, 國內對水鳥遷徙動態的研究已有較多報道(Ma et al, 2002a, b; Zhang et al, 2004; Ge et al, 2006; Yang et al, 2009), 但將種間相互作用運用于水鳥遷徙動態分析的研究仍較少(Zhang et al, 2004; Yang et al, 2009)。
杭州灣地處長江三角洲經濟發達的滬、杭、甬金三角地帶, 其保護與利用的矛盾非常突出。在該區域開展水鳥資源調查, 以了解其群落組成和季節動態, 并研究遷徙水鳥的種間相關性, 將為該區域水鳥資源監測、保護和合理利用濕地資源提供重要的基礎資料和理論依據。
1 研究區域概況
杭州灣位于我國東部沿海的浙江省東北部,為喇叭狀河口灣, 西界與錢塘江相連, 東界為蘆漕港閘與長跳咀邊線。濕地總面積5.86×104hm2,其中淺海水域3.18×104hm2、潮間淤泥海灘2.05×104hm2、潮間鹽水沼澤6 244 hm2, 海拔?5~5 m(Zhejiang Forestry Administration, 2002)。除錢塘江河口外, 灣內還包含曹娥江、甬江等重要河口,其南岸屬淤漲型海岸, 三北淺灘是浙江最大的海涂,而北岸則屬侵蝕型海岸。濕地類型以淺海水域和期間淤泥海灘為豐, 其它尚有潮間鹽水沼澤和巖石性海岸。杭州灣為一喇叭口形狀的河口海灣, 灣內海域水深都小于l0 m, 水下地形平坦, 中北部至口門為杭州灣水下淺灘; 灣內有大小島嶼69個, 島嶼附近發育有潮流深槽、沖刷深潭及潮流沙脊; 海岸線長258.49 km, 以人工海岸為主。
近年來持續的大規模圍墾使得杭州灣灘涂濕地急劇減少, 其中2004—2008年間每年減少10%以上(National Marine Environmental Monitoring Center, 2009), 由于普遍采用了中、低灘圍墾方式,現有自然灘涂上的植被狀況較差, 以低灘鹽藻光灘為主, 狹窄帶狀分布的植被覆蓋區域較少。為了解決圍墾區生活用水和工農業用水問題, 杭州灣南岸目前有中小型海涂水庫10余個, 水深均在3 m以上,面積最大的四灶浦水庫, 達5.12 km2, 水深5.7 m。
2 研究方法
2007年7月—2008年6月, 對杭州灣上虞曹娥江口至鎮海甬江口之間潮間帶及其附近海涂水庫的水鳥群落進行逐月統計。在隨后的2008年7月—2010年11月繼續開展季度監測。我們將一年分為4期:12—次年2月(越冬期)、3—5月(北遷期)、6—8月(繁殖期)和9—11月(南遷期)。季度監測是指每年每期選擇其中一個月進行調查, 為了更好地掌握研究區域內的鳥類資源組成和動態, 第二年的同期調查時間順延至下一個月進行, 循環往復。
采用直接計數法, 直接記錄水鳥種類和數量,計數方法以精確記數為主, 估算為輔, 并結合“集團統計法”, 推算水鳥的總數以及群落中各物種所占百分比(Howes & Endagama, 1995)。調查時用Kowa BD42-8GR雙筒望遠鏡和Zeiss Diascope 85T單筒望遠鏡觀察鳥類。鳥類分類系統參考《中國鳥類分類與分布名錄》(Zheng, 2005), 中文鳥名主要參考《中國鳥類野外手冊》(Mackinnon et al, 2000)。
調查時間, 潮間帶鳥類調查根據潮汛情況, 選擇在高潮點前后2 h內進行。為避免調查數據重復,在鳥類數量統計中采用最大值保留法(Howes & Bakewell, 1989), 并將種群數量超過10%的種群定為優勢種。
對水鳥物種數的月動態分析采用了2007年7月—2010年11月的所有監測數據。利用2007年7月—2008年6月的逐月調查數據, 對水鳥數量的年動態進行分析, 并對記錄的鸻鷸類和鴨類分別進行兩兩配對的種間數量隨時間變化的Spearman相關性分析。為了提高分析的準確性, 在各遷徙階段中種群數量低于1%的物種未納入上述分析。數據處理采用SPSS10.0進行。
3 結 果
3.1 水鳥種類組成
杭州灣及錢塘江河口濕地共記錄水鳥9目18科128種(表1)。候鳥119種, 占總數的93.0%〔其中, 冬候鳥64種(50.0%), 旅鳥34種(26.6%), 夏候鳥17種(13.3%)及迷鳥4種(3.1%)〕。繁殖鳥(夏候鳥和留鳥)26種, 占該地水鳥總數的20.3%。鸻形目、雁形目、鸛形目和鷗形目為記錄物種最多的4個目,合計占該地水鳥總數的87.5%。本區域的優勢種隨遷徙階段的變化而變動, 自然潮間帶的優勢種有黑腹濱鷸(Calidris alpina)、黑尾塍鷸(Limosa limosa)、紅頸濱鷸(Calidris ruficollis)等12種, 海涂水庫的優勢種包括羅紋鴨(Anas falcata)、斑嘴鴨(Anas poecilorhyncha)等5種。
杭州灣及錢塘江河口濕地共記錄列入國家重點保護野生動物名錄的水鳥有11種, 包括國家I級重點保護鳥類東方白鸛(Ciconia boyciana)、中華秋沙鴨(Mergus squamatus)和遺鷗(Larus relictus), 列入IUCN中國鳥類受威脅狀況名錄的鳥類數量17種, 包括瀕危EN等級的中華秋沙鴨、青頭潛鴨(Aythya baeri)、東方白鸛和黑臉琵鷺(Platalea minor)。此外, 杭州灣及錢塘江河口共有88種水鳥列入中日候鳥保護名錄, 42種列為中澳候鳥保護名錄。
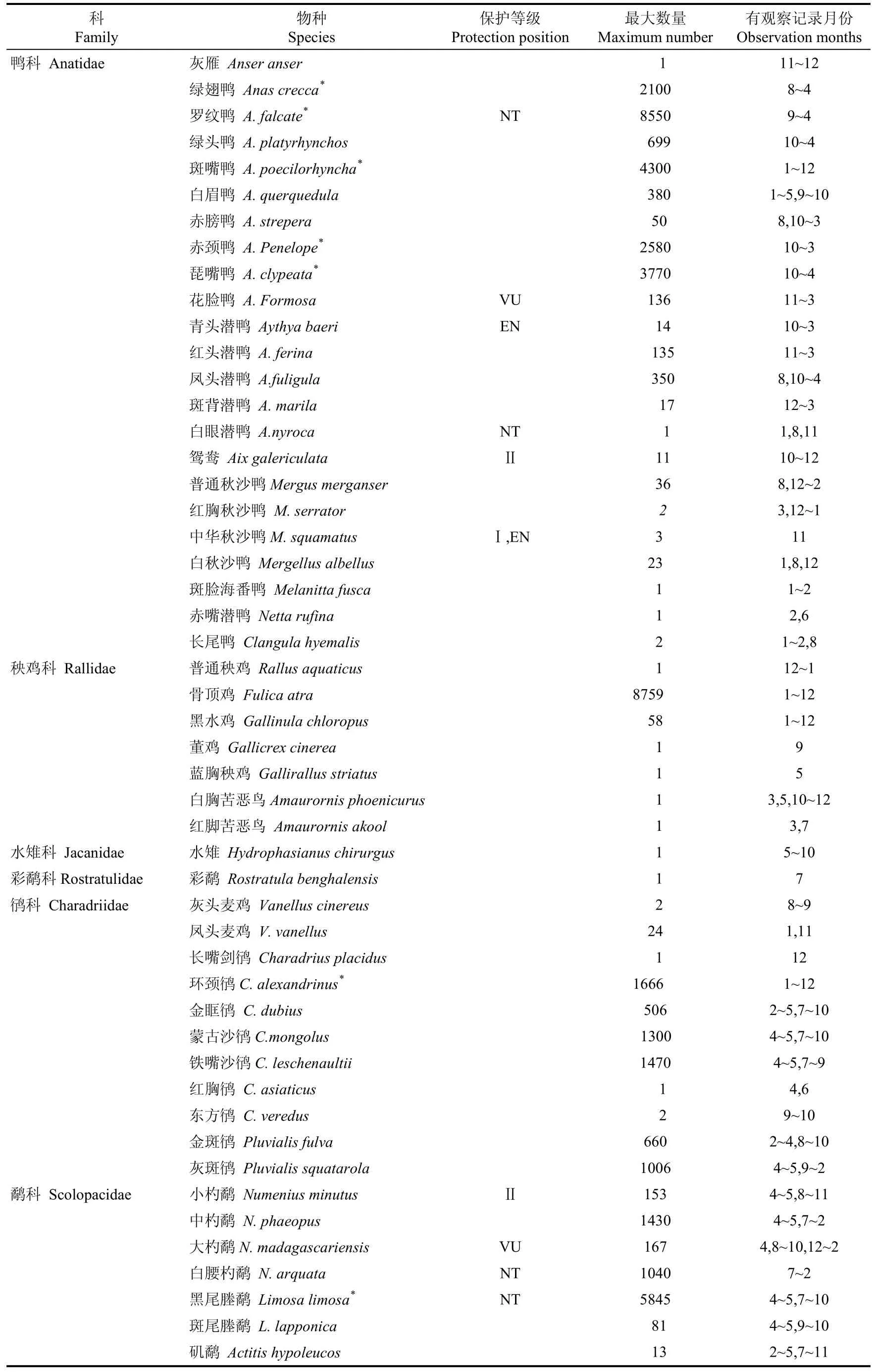
科Family物種Species保護等級Protection position最大數量Maximum number有觀察記錄月份Observation months鴨科 Anatidae 灰雁 Anser anser 1 11~12綠翅鴨 Anas crecca* 2100 8~4羅紋鴨 A. falcate* NT 8550 9~4綠頭鴨 A. platyrhynchos 699 10~4斑嘴鴨 A. poecilorhyncha* 4300 1~12白眉鴨 A. querquedula 380 1~5,9~10赤膀鴨 A. strepera 50 8,10~3赤頸鴨 A. Penelope* 2580 10~3琵嘴鴨 A. clypeata* 3770 10~4花臉鴨 A. Formosa VU 136 11~3青頭潛鴨 Aythya baeri EN 14 10~3紅頭潛鴨 A. ferina 135 11~3鳳頭潛鴨 A.fuligula 350 8,10~4斑背潛鴨 A. marila 17 12~3白眼潛鴨 A.nyroca NT 1 1,8,11鴛鴦 Aix galericulata Ⅱ 11 10~12普通秋沙鴨Mergus merganser 36 8,12~2紅胸秋沙鴨 M. serrator 2 3,12~1中華秋沙鴨M. squamatus Ⅰ,EN 3 11白秋沙鴨 Mergellus albellus 23 1,8,12斑臉海番鴨 Melanitta fusca 1 1~2赤嘴潛鴨 Netta rufina 1 2,6長尾鴨 Clangula hyemalis 2 1~2,8秧雞科 Rallidae 普通秧雞 Rallus aquaticus 1 12~1骨頂雞 Fulica atra 8759 1~12黑水雞 Gallinula chloropus 58 1~12董雞 Gallicrex cinerea 1 9藍胸秧雞 Gallirallus striatus 1 5白胸苦惡鳥Amaurornis phoenicurus 1 3,5,10~12紅腳苦惡鳥 Amaurornis akool 1 3,7水雉科 Jacanidae 水雉 Hydrophasianus chirurgus 1 5~10彩鷸科Rostratulidae 彩鷸 Rostratula benghalensis 1 7鸻科 Charadriidae 灰頭麥雞 Vanellus cinereus 2 8~9鳳頭麥雞 V. vanellus 24 1,11長嘴劍鸻 Charadrius placidus 1 12環頸鸻C. alexandrinus* 1666 1~12金眶鸻 C. dubius 506 2~5,7~10蒙古沙鸻C.mongolus 1300 4~5,7~10鐵嘴沙鸻C. leschenaultii 1470 4~5,7~9紅胸鸻 C. asiaticus 1 4,6東方鸻 C. veredus 2 9~10金斑鸻 Pluvialis fulva 660 2~4,8~10灰斑鸻 Pluvialis squatarola 1006 4~5,9~2鷸科 Scolopacidae 小杓鷸 Numenius minutus Ⅱ 153 4~5,8~11中杓鷸 N. phaeopus 1430 4~5,7~2大杓鷸N. madagascariensis VU 167 4,8~10,12~2白腰杓鷸 N. arquata NT 1040 7~2黑尾塍鷸 Limosa limosa* NT 5845 4~5,7~10斑尾塍鷸 L. lapponica 81 4~5,9~10磯鷸 Actitis hypoleucos 13 2~5,7~11
* 代表該物種是某個遷徙階段的優勢種;* Represents a dominant species in the waterbird community of one or more migration stages;I和Ⅱ:分別代表國家一和二級重點保護野生動物;I represents the national first-grade protection of wildlife in China and II represents the second-grade level of protection;CR、EN、VU、NT:世界自然保護聯盟IUCN保護等級IUCN Red List Categories and Criteria, CR:極危(Critically Endangered);EN:瀕危(Endangered);VU:易危(Vulnerable);NT:近危(Near Threatened)。
3.2 水鳥種類及數量動態
杭州灣及錢塘江河口水鳥物種數的年動態曲線總體上呈現兩個峰值(圖1), 分別是10月73種和4月70種, 而6月份水鳥物種數全年最低, 僅記錄24種, 另一個低谷出現在3月(54種)。對鸻形目、雁形目、鸛形目和鷗形目這4種優勢類群物種數動態分析結果顯示, 鸻形目和雁形目物種數年動態波動較大, 而鸛形目和鷗形目物種數波動較小; 鸻形目鳥類物種數年動態呈現“雙峰”曲線, 分別在4和9月份達到最高值36和39種, 其低谷值出現在3和6月, 分別為11和4種。雁形目鳥類物種數動態總體上表現為一單峰曲線, 峰值出現在12月, 記錄25種, 5—7月為低谷期, 僅記錄1~3種。鸛形目和鷗形目物種數的最小值分別出現1—3月和6—7月, 分別僅記錄7和2種。
杭州灣及錢塘江河口水鳥個體數量的年動態曲線與其物種數動態曲線存在較大差異, 總體上呈現3個峰值(圖2), 其中最低值出現在6月, 記錄個體數量為4 316只, 最高值出現在3月初, 記錄水鳥114 395只。鸻形目鳥類數量動態曲線與總曲線較為相似, 其最小值出現在6月(430只), 最高值在3月(101297只); 雁形目鳥類數量動態曲線呈“雙峰”,峰值分別出現在11和3月, 分別記錄10 622和
10 862只; 與前兩者相比, 鸛形目和鷗形目個體數量總體偏少, 波動相對平緩; 鸛形目個體數量高峰期出現在繁殖期6和8月, 分別記錄了3 561和11 367只, 其余月份記錄數量均低于600只; 鷗形目數量動態曲線總體上呈現3個峰值, 分別為9、 11和3月, 分別記錄1 628、4 892和1 348只。
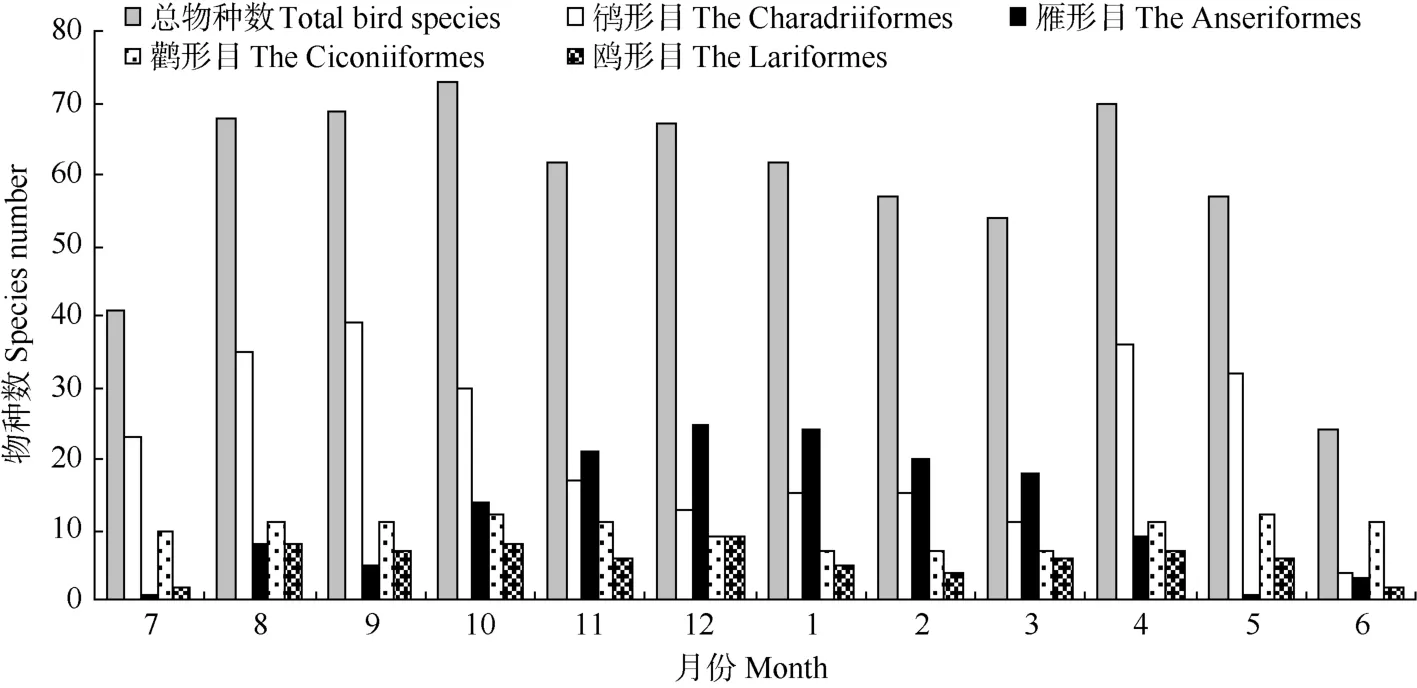
圖1 杭州灣及錢塘江河口水鳥物種數的月動態Fig. 1 Monthly dynamics of waterbirds species in Qiantangjiang River estuary and Hangzhou Bay
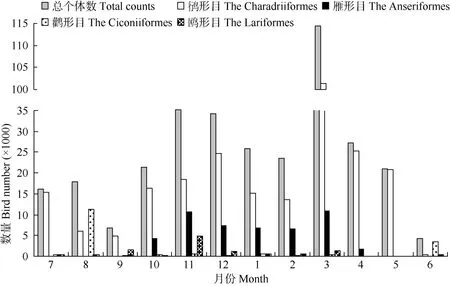
圖2 杭州灣及錢塘江河口水鳥數量的月動態Fig. 2 Monthly dynamics of waterbirds number in Qiantangjiang River estuary and Hangzhou Bay
3.3 遷徙水鳥種間相關性分析
對杭州灣及錢塘江河口鸻鷸類和鴨類的物種數量變化進行了種間相關性分析結果顯示(表2、表3), 進行兩兩配對相關性分析的13種鸻鷸類中, 有20對為顯著正相關, 其中13對為極顯著正相關, 有1對為顯著負相關。蒙古沙鸻(Charadrius mongolus)、鐵嘴沙鸻(C. leuschenaultii)、黑尾塍鷸、紅頸濱鷸和尖尾濱鷸(Calidris acuminate)之間均極顯著正相關。金眶鸻(C. dubius)與前4種鸻鷸類(不包括紅頸濱鷸)顯著正相關, 且與金斑鸻(Pluvialis dominica)為極顯著正相關, 與青腳鷸為顯著正相關。與金眶鸻類似, 青腳鷸與前5種極顯著正相關鸻鷸旅鳥群中的4種(不包括鐵嘴沙鸻)顯著正相關, 其中與蒙古沙鸻、紅頸濱鷸極顯著相關, 還與金眶鸻顯著相關。灰斑鸻與金斑鸻顯著負相關。
9種鴨類進行了種間相關性分析, 有23對為顯著正相關, 其中10對為顯著正相關。除針尾鴨(Anasacuta)和琵嘴鴨(A. clypeata)之間無顯著相關性外,綠翅鴨(A. crecca)、羅紋鴨、斑嘴鴨、赤頸鴨(A. penelope)、鳳頭潛鴨(Aythya fuligula)、針尾鴨與琵嘴鴨之間均兩兩顯著正相關。綠頭鴨(Anas platyrhynchos)僅與鳳頭潛鴨、針尾鴨和綠翅鴨之間顯著正相關。而白眉鴨(A. querquedula)與其它任一種鴨類均無顯著相關性。
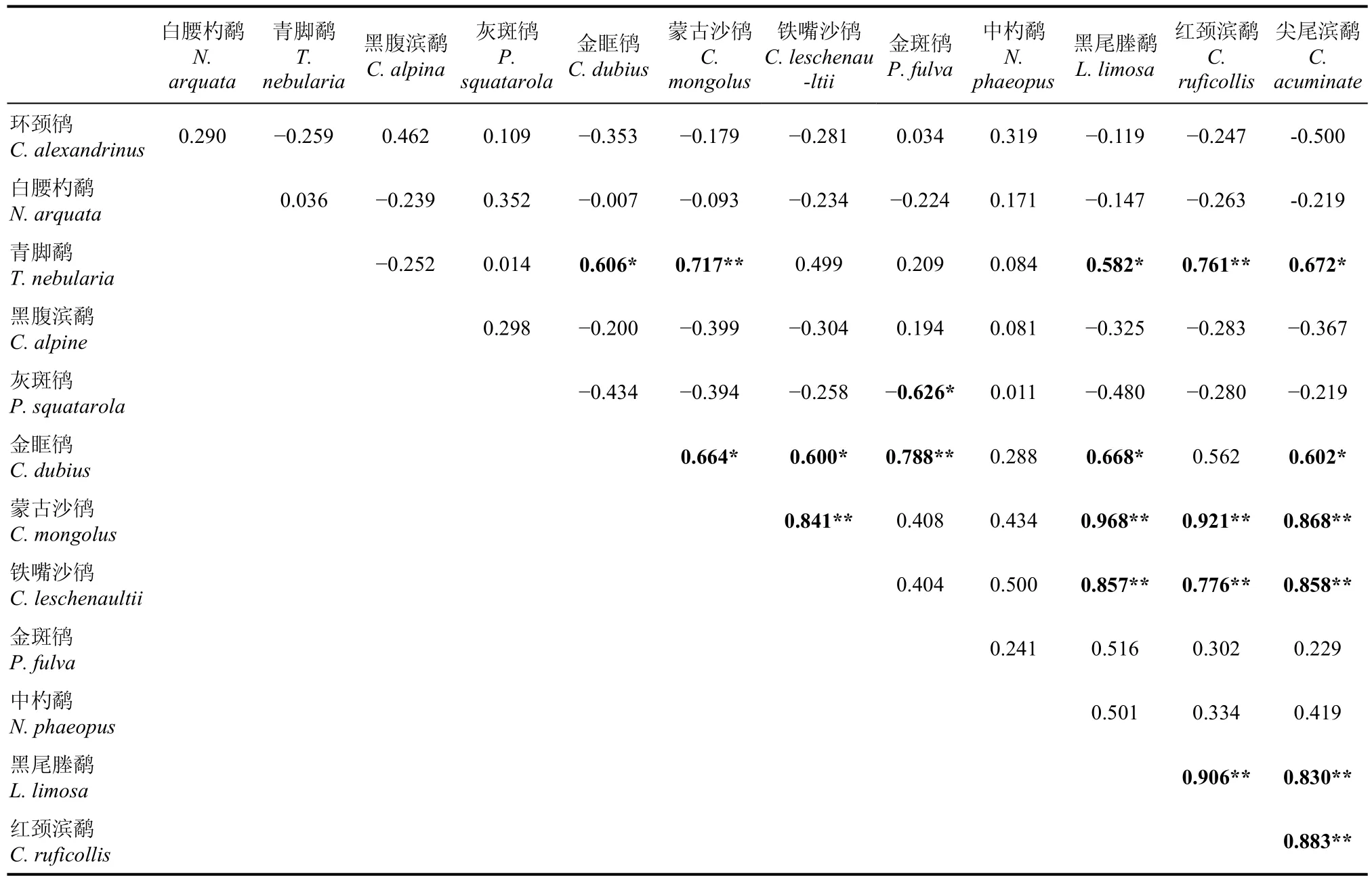
表2 13種鸻鷸類數量變化的相關系數及顯著性矩陣Tab. 2 Correlation coefficient matrix of 13 species of shorebirds in Qiantangjiang River estuary and Hangzhou Bay
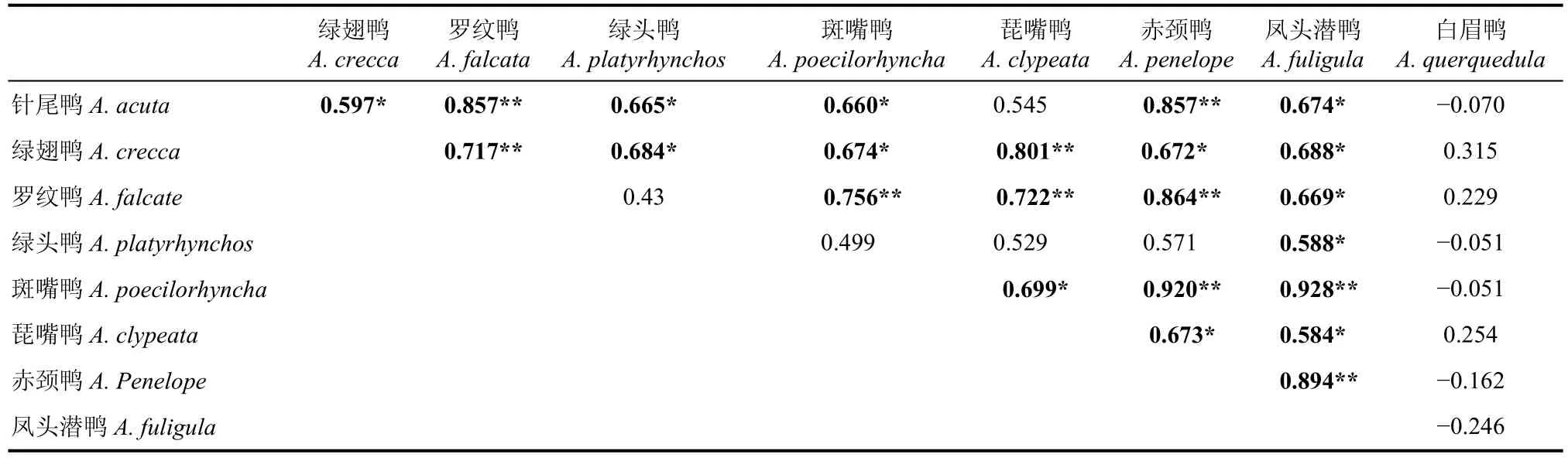
表3 8種鴨類數量變化的相關系數及顯著性矩陣Tab. 3 Correlation coefficient matrix of eight species of duck in Qiantangjiang River estuary and Hangzhou Bay
4 討 論
4.1 種間相關性與水鳥遷徙模式
研究結果顯示, 在杭州灣及錢塘江河口出現的鸻鷸類中, 參與種間相關性分析的7種旅鳥中的5種(包括黑尾塍鷸、紅頸濱鷸、尖尾濱鷸、蒙古沙鸻和鐵嘴沙鸻)之間呈現兩兩顯著正相關, 表明它們的遷徙模式是相同的, 而中杓鷸和金斑鸻與前5者之間及兩者內部均沒建立這種顯著性聯系。這可能表明旅鳥群中至少存在3種遷徙模式。以上述5種顯著相關性鸻鷸類旅鳥為優勢種構成的遷徙水鳥群, 其遷徙時間集中在7月下旬—9月下旬(秋季南遷時期)和3月下旬—5月中旬(春季北遷時期)。參與相關性分析的鸻鷸類冬候鳥種間均無顯著相關性,表明黑腹濱鷸、白腰杓鷸、灰斑鸻、青腳鷸等冬候鳥之間的遷徙模式可能是不同的。金眶鸻、青腳鷸均與由5種旅鳥組成的極顯著相關旅鳥群中的4種顯著正相關, 表明其具有與后者類似的遷徙模式。生態位相近的灰斑鸻與金斑鸻顯著負相關, 兩者的遷徙模式可能截然相反。中杓鷸、白腰杓鷸、黑腹濱鷸和環頸鸻與參與分析的其他13種鸻鷸類之間均無顯著相關性, 表明其具有相對獨立的遷徙節律。
而在該區域越冬的鴨類群落中, 5種優勢種包括綠翅鴨、羅紋鴨、斑嘴鴨、琵嘴鴨和赤頸鴨及鳳頭潛鴨之間顯示了顯著的正相關性, 針尾鴨與前5種鴨類中的4種(不包括琵嘴鴨)建立了種間顯著正相關, 僅白眉鴨與其他鴨類無顯著相關性(表3)。這表明鴨類的遷徙模式相對單一, 大多數鴨類都從10月上旬開始大規模的遷徙至杭州灣, 并于次年4月中旬全部遷離。
杭州灣是東亞—澳大利西亞水鳥遷徙通道中一個重要的中轉停歇地(Barter, 2002), 而種間吸引作用可能是杭州灣遷徙水鳥群落形成的一個重要驅動因子。鳥類群落中的種間吸引作用已被實驗所證實(Mikko & Jukka, 2002; Robert et al, 2003; Sebastián-González et al, 2010), 但Mikko et al(1999)認為, 種間吸引作用可能僅在種間競爭足夠低時才處于優勢。食物資源是影響鳥類中途停歇地選擇的重要因素之一(Hicklin & Smith, 1979; Wilson, 1990)。但已有研究表明,食物資源可能并不是杭州灣鸻形目鳥類的遷徙規模和數量動態的限制因素。杭州灣潮間帶大型底棲動物功能群的密度和絕大多數種類數分布在季節間差異均不顯著(Li et al, 2007), 崇明東灘的研究結果與此類似(Jing, 2005)。我們的研究結果顯示(圖2), 杭州灣鸻鷸類水鳥群落的數量的季節變動最高值在10萬只左右, 表明除該高峰期外, 杭州灣的食物資源相對于鸻鷸類來說可能是充裕的, 因而鸻鷸類因食物而出現的種內和種間的競爭將處于較低的水平。進入越冬期(12—次年2月)的鴨類群落數量均大幅低于春秋季遷徙季節(圖2), 表明此時的鴨類群落數量處于相對穩定的狀態, 其種間競爭水平可能較低。
出于共同的生境需求, 個體可能將不同種的存在狀況作為評價棲息地斑塊質量的一個指標(Mikko & Jukka, 2002; Jukka & Robert, 2008; Hromada et al, 2008)。這對于剛到達繁殖地的遷徙鳥類來說更為重要(Robert et al, 2003; Jukka & Robert, 2008; Hromada et al, 2008; Jukka et al, 2009)。對于是否停留它們必須快速地做出抉擇, 而留鳥的出現與否及種群情況將是其重要的參考因素(Sebastián-González et al, 2010)。種間吸引假說同時認為, 多物種的群落中的個體能夠降低其被捕食風險(Mikko et al, 1990)。本研究結果中, 7種參與分析的旅鳥中的6種與反映該區域棲息地質量的繁殖鳥金眶鸻或冬候鳥青腳鷸顯著正相關。而5種旅鳥組成的極顯著相關群中的任兩種的最初接觸可能也是基于一方能反映另一方遷徙路線上的中轉停歇地的生境質量, 最終基于共同的生境需求(獲取食物和降低捕食風險), 同時最小化種間排斥作用而有選擇的集群遷徙。Recher(1966)認為遷徙鸻鷸類集群中所有的種間差異和瞬態的集群特征都明顯為了最大限度的減少種間排斥作用。這種觀點也體現在我們的研究結果中, 極顯著相關群中的5種旅鳥的喙長是不同的, 而喙長與取食方式有關(Brown & Wilson, 1956), 不同的喙長意味著食物選擇的分化(Wang & Qian, 1988)。
對于在杭州灣越冬的鴨類來說, 由于地處長江三角洲經濟發達的滬、杭、甬金三角地帶, 降低被捕食(主要為人類的捕食活動)風險將是影響其棲息地選擇尤其是休息場所選擇的重要因素之一。本研究結果中, 鳳頭潛鴨與除白眉鴨外的7種鴨類顯著正相關, 而其也是最先來到杭州灣的鴨類之一, 但其食性以動物性食物為主, 因而其在水庫的出現與否及種群數量大小可能成為后來者評價該水庫生境質量(不包括食物資源)的重要指標。
4.2 杭州灣及錢塘江河口水鳥群落遷徙規律
根據杭州灣及錢塘江河口水鳥物種及數量遷徙動態(圖1、圖2)的研究結果, 本區域水鳥群落整個遷徙過程可分為6個階段。
第一階段從7月下旬開始—9月下旬, 遷徙高峰出現在8月中旬, 這一階段的遷徙水鳥群落的主體是以黑尾塍鷸、紅頸濱鷸及蒙古沙鸻為優勢種群的鸻鷸類旅鳥。在該階段, 鸛形目的鷺鳥正處于繁殖期, 8月中旬大量亞成鳥離巢, 鷺鳥數量達到最高峰, 9月下旬鷺科夏候鳥陸續南遷; 而鸛形目白琵鷺和黑臉琵鷺從7月下旬開始—8月陸續遷飛至此短暫停歇。以白額燕鷗為主體的鷗類繁殖鳥及鷗嘴噪鷗(Gelochelidon nilotica)開始南遷, 同時以西伯利亞銀鷗(Larus vegae)和黑嘴鷗為主體的冬候鳥開始遷飛到達。
第二階段從10月上旬—12月中旬, 遷徙水鳥群落以冬候鳥的鸻鷸類和雁鴨類為主。這一階段的鸻鷸類以黑腹濱鷸為主要優勢種。從10月上旬開始, 大規模鴨群陸續到來, 11月下旬鴨類群落數量達到一個小高峰, 并在12月中旬進入相對穩定期。從11上旬開始, 大型水鳥如鸛形目的黑臉琵鷺、白琵鷺、雁鴨類的小天鵝、豆雁(Anser fabalis)以及鵜形目的卷羽鵜鶘等陸續遷飛至此。11下旬, 以黑尾鷗(Larus crassirostris)為優勢種的鷗類冬候鳥個體數量達到了全年的最高值。
第三階段從12月下旬—次年2月中旬, 為越冬穩定期。這時水鳥群落以冬候鳥為主, 群落物種組成與數量相對比較穩定, 鸻鷸類以黑腹濱鷸為絕對優勢種, 其種群數量占總數的80%以上。
第四階段從2月下旬—3月下旬, 為水鳥遷徙期。該階段以黑腹濱鷸為主的冬候鳥遷徙首先開始, 2月下旬—3月上旬, 黑腹濱鷸的種群數量驟升至前期的5倍以上, 之后其數量回落到越冬期相近的水平。3月中旬, 鴨類遷徙高峰到來, 其個體數量在3月下旬達到一個高峰之后迅速下降。3月下旬鸻鷸類旅鳥陸續遷來。小天鵝則在2月下旬就已經遷離。3月中旬鵜形目的卷羽鵜鶘遷飛至此, 短暫停留后即遷離。
第五階段從4月上旬—5月中旬, 為以紅頸濱鷸、尖尾濱鷸及黑尾塍鷸為優勢種的旅鳥群的遷徙高峰期。但該階段, 以黑腹濱鷸為優勢種的冬候鳥群仍為水鳥群落的主體。鴨類在4月中旬基本遷離。3月下旬—4月下旬, 陸續有鸛形目黑臉琵鷺和白琵鷺遷飛至此, 作短暫停留后即遷離。
第六階段從5月下旬—7月中旬, 為候鳥的繁殖季節。此階段的水鳥群落主要由鷺鳥和以白額燕鷗為優勢種的鷗類及普通燕鸻等夏候鳥構成。鷺鳥夏候鳥從2月開始就陸續遷來, 5月至7月進入繁殖高峰期。4月中下旬, 普通燕鸻就已經遷來, 而白額燕鷗也在此時出現在杭州灣, 但前者的個體數量在7月中旬即達到高峰, 而后者則在繁殖季節結束后的8月下旬—9月上旬達到高峰。
從杭州灣及錢塘江河口水鳥季節動態分析中,我們發現該區域春季鸻鷸類遷徙高峰在數量上遠超過秋季遷徙高峰, 達到了后者的4倍以上, 類似的研究結果出現在崇明東灘(Qian et al, 1985; Ma et al, 2002b)。馬志軍等認為, 春季鸻鷸類遷徙高峰數量遠高于秋季的主要原因在于崇明東灘在鸻鷸類遷徙路線中的特殊地理位置(Ma et al, 2002b)。我們的研究結果與此類似, 杭州灣及錢塘江河口水鳥秋季遷徙高峰數量上的巨大差異來自于該區域的特殊地理位置和水鳥群落構成。對春秋季遷徙季節的水鳥群落組成分析結果顯示, 從10月上旬開始—次年3月上旬, 黑腹濱鷸的種群數量占鸻鷸類群落總數的比例均維持在80%以上, 并在3月上旬達到98%, 其后的4—5月, 雖然其種群數量開始下降,但仍維持在總數的60%以上。已有研究表明太平洋東海岸的黑腹濱鷸會采取一系列的短距離輕跳方式, 沿海岸北上到達繁殖地, 而南遷過程中, 則可能直接飛越阿拉斯加海灣到達較南的停歇地或越冬地(Warnock et al, 2004)。位于太平洋西海岸的東亞—澳大利西亞水鳥遷徙通道的黑腹濱鷸也可能采用類似的遷徙方式。崇明東灘的黑腹濱鷸在北遷時具有較高的能量儲備, 可能是為遷徙飛行做準備(Cai, 2009)。由于黑腹濱鷸在中國東南部地區越冬(Barter et al, 2006), 杭州灣所在的區域有可能是其遷飛前的最后一個能量補給站, 因而該區域黑腹濱鷸的種群數量在其遷徙前夕急劇增加, 但在其2月下旬—3月上旬的首輪遷徙高峰結束后, 黑腹濱鷸的種群數量顯著回落。
Barter M. 2002. Shorebirds of the Yellow Sea: Importance, threats and conservation status. Australia[M]//Wetlands International Global Series 9: International Wader Studies 12. Canberra, Australia: [s.n.].
Barter M, Lei G, Cao L. 2006. Waterbird Survey of the Middle and Lower Yangtze River Floodplain in February 2005[M]. Yang Q, Translation. Beijing: China Forestry Publishing House. [馬克·巴特,雷剛,曹壘. 2006.長江中下游水鳥調查報告(2005年2月).楊琴,譯.北京:中國林業出版社.]
Brown WL, Wilson EO. 1956. Character displacement[J]. Syst. Zool,5(2): 49-64.
Cai ZY. 2009. Ecology of Dunlin (Calidris alpine) in Chongming Dongtan[D]. MSc thesis, Fudan University. [蔡志揚. 2009.崇明東灘黑腹濱鷸的生態學研究.碩士學位論文,復旦大學.]
D?nhardt J, Lindstr?m ?.2001. Optimal departure decision of songbirds from an experimental stopover site and the significance of weather[J]. Anim Behav.,62: 235-243.
Ge ZM, Wang TH, Zhou X, Shi WY. 2006. Seasonal change and habitat selection of shorebird community at the South Yangtze River Mouth and North Hangzhou Bay, China[J]. Acta Ecol Sin,26(1):40-47. [葛振鳴,王天厚,施文彧,周曉. 2006.長江口杭州灣鵒形目鳥類群落季節變化和生境選擇.生態學報,26(1): 40-47. ]
Hicklin PW, Smith PC. 1979. The diets of five species of migrant shorebirds in the Bay of Fundy[J]. Proc NS Ins Sci,29: 483-488.
Howes M, Endagama P. 1995. Farmers, Forests and Fuel: Towards a New Biomass Energy Strategy for Sri Lanka[M]. London: Intermediate Technology Publications.
Howes J, Bakewell D. 1989. Shorebird Studies Manual[M]. Malaysia: AWB Publication, NO.55.
Hromada M, Antczak M, Valone TJ, Tryjanowski P. 2008. Settling decisions and heterospecific social information use in shrikes[J]. PLoS ONE,3(12): e3930.
IUCN. 2011. IUCN Red List of Threatened Species.Version 2011.1.[R/OL] http://www.iucnredlist.org.
Jing K. 2005. Stopover ecology of shorebirds in Chongming Dongtan, Shanghai[D]. PH.D. thesis, Fudan University, Shanghai. [敬凱. 2005.上海崇明東灘鸻鷸類中途停歇生態學研究.博士學位論文.復旦大學.]
Jukka TF, Robert LT. 2008. Evidence of information collection from heterospecifics in cavity-nesting birds[J]. Ibis,150: 409-412.
Jukka TF, Ma°rten BH, Lars G.2009. Experimental evidence for the use of density based interspecific social information in forest birds[J]. Ecography,32: 539-545.
Li HH, Bao YX, Hu ZY, Ge BM. 2007. Seasonal dynamics of macrobenthic functional groups and trophic levels in the bridge construction zone at the South Bank of Hangzhou Bay, China[J]. Acta Zool Sin,53(6): 1011-1023. [李歡歡,鮑毅新,胡知淵,葛寶明.2007.杭州灣南岸大橋建設區域潮間帶大型底棲動物功能群及營養等級的季節動態.動物學報,53(6): 1011- 1023.]
Lu JJ, Shi M, Cui ZX. 1988. A preliminary study on the over-wintering wader (Charadriiformes) community in the North Coast of East China Sea[J]. Chn J Ecol,7(6): 19-22. [陸健健,施銘,崔志興. 1988.東海北部沿海越冬鸻鷸群落的初步研究.生態學雜志,7(6): 19-22.]
Mackinnon J, Philipps K, He FQ. 2000. A Field Guide to the Birds of China [M]. Lu HF, Translation. Changsha: Hunan Education Press. [馬敬能,菲利普斯,何芬奇. 2000.中國鳥類野外手冊.盧和芬,譯.長沙:湖南教育出版社.]
Ma ZJ, Jing K, Tang SM, Chen JK. 2002a. Shorebirds in the Eastern intertidal areas of Chongming Island during the 2001 northward migration [J]. Stilt,41: 6-10.
Ma ZJ, Tang SM, Lu F, Chen JK.2002b. Chongming Island: a less important shorebirds stopover site during southward migration? [J]. Stilt,41: 35-37.
Mikko M, Pekka H, Gerald JN, Kent M. 1990. Numerical and behavioural responses of migrant passerines to experimental manipulation of resident tits (Parus spp.): heterospecific attraction in northern breeding bird communities? [J] Oecologia,85: 218-255.
Mikko M, Roger H, Jukka TF, Juha T. 1999 Evolution of heterospecific attraction: Using other species as cues in habitat selection[J]. Evol. Ecol,13: 91-104.
Mikko M, Jukka TF. 2002. Heterospecific attraction among forest birds: a review[J]. Ornithol Sci,1: 41-51.
National Marine Environmental Monitoring Center. 2009. The Marine Environment Quality Bulletin of China 2009 [EB/OL]. http://www. mem.gov.cn/ indexshow.html [國家海洋環境監測中心. 2009. 2009年中國海洋環境質量公報. http://www.mem.gov.cn/ indexshow.html.]
Qian GZ, Cui ZX, Wang TH. 1985. Waders (charadiiformes) community of the Chang-jiang river estuary and the Hang-zhou bay[J]. Acta Zool Sin,31(1): 96-97. [錢國楨,崔志興,王天厚. 1985.長江口杭州灣北部的鸻形目鳥類群落.動物學報,31(1): 96-97.]
Recher HF. 1966. Some aspects of the ecology of migrant shorebirds[J]. Ecology,47(3): 393-407.
Robert LT, Jukka TF, Mikko M. 2003. Positive interactions between migrant and resident birds: testing the heterospecific attraction hypothesis[J]. Oecologia,134: 431-438.
Sebastián-González E, Sánchez-Zapata JA, Botella F, Ovaskainen O. 2010. Testing the heterospecific attraction hypothesis with time-series data on species co-occurrence[J]. Proc R Soc,277: 2983-2990.
Wang TH, Qian GZ. 1988. Waders (Charadiiformes) Community of the Chang-jiang River Estuary and the Hang-zhou Bay[M]. Shanghai: East China Normal University Press. [王天厚,錢國楨. 1988.長江口杭州灣鸻形目鳥類.上海:華東師范大學出版社.]
Warnock ND, Takekawa JY, Bishop MA. 2004. Migration and stopover strategies of individual Dunlin along the Pacific coast of North America[J]. Can J Zool,82: 1687-1697.
Wilson WH Jr. 1990. Relationship between prey abundance and foraging site selection by semipalmated sandpipers on a Bay of Fundy mudflat[J]. J Field Ornithol,61: 9-19.
Wilson JR, Barter MA. 1998. Identification of potentially important staging area for “long jump” migrant waders in the East Asian-Australian Flyway during northward migration[J]. Stilt,32: 16-26.
Yang CC, Cai Y, Liang W, Zhang GG, Shi HT. 2009. Waterbirds diversity, seasonal dynamics, and interspecific correlation in the wetlands of Beili and Houshui Bays, Hainan[J]. J Biodivesr Sci,17(3): 226-232. [楊燦朝,蔡燕,梁偉,張國鋼,史海濤. 2009.海南北黎灣和后水灣濕地水鳥的季節動態、物種豐富度和種間相關性.生物多樣性,17(3): 226-232.]
Zhang SP, Zhang ZW, Xu JL, Sun QH, Liu DP. 2004. Seasonal changes and interspecific correlation among the migratory waterbird communities in Tianjin[J]. Acta Ecol Sin,42(4): 666-673. [張淑萍,張正旺,徐基良,孫全輝,劉東平. 2004.天津地區遷徙水鳥群落的季節動態及種間相關性分析.生態學報,42(4): 666-673.]
Zhejiang Forestry Administration. 2002. Zhejiang Forestry Natural Resources-Wetland [M]. Beijing: Chinese Agriculture Science and Technology Publishing House. [浙江省林業局. 2002.浙江林業自然資源(濕地卷).北京:中國農業科學技術出版社.]
Zheng GM. 2005. A Checklist on the Classification and Distribution of the Birds of China[M]. Beijing: Science Press. [鄭光美. 2005.中國鳥類分類與分布名錄.北京:科學出版社.]
Zhuge Y, Jiang SR, Zheng ZW, Fang GW. 1986. Preliminary studies on geographical ecology of birds and mammals on some islands of Zhejiang Province[J]. Acta Zool Sin,32(1): 74-85. [諸葛陽,姜仕仁,鄭忠偉,方國偉. 1986.浙江海島鳥獸地理生態學的初步研究.動物學報,32(1): 74-85.]
Zhu X, Yang CJ. 1988. A survey of birds in Zhejiang Province[J].J Zhejiang For Coll,5(3): 243-258. [朱曦,楊春江. 1988.浙江鳥類研究.浙江林學院學報,5(3): 243-258.]
Zhu X. 1989a. Preliminary studies on the ecological types and geographical distribution of birds in Zhejiang[J]. J Zhejiang For Coll,6(3): 283-289. [朱曦.浙江省鳥類的生態地理初步研究. 1989.浙江林學院學報,6(3): 283-289. ]
Zhu X, Xuan ZC, Chen LQ, Gao YJ, Ma XX. 1989b. Preliminary investigation on spring bird in East Coast of Zhejiang Province[J].J Zhejiang For Sci Technol,9(1): 35-40. [朱曦,宣志燦,陳李群,高銀江,麻秀新. 1989.浙江東部沿海春季鳥類初步調查.浙江林業科技,9(1): 35-40.]
Community composition, seasonal dynamics and interspecific correlation of waterbirds in the Qiantangjiang River estuary and Hangzhou Bay
JIANG Ke-Yi, WU Ming, SHAO Xue-Xin
(Research Institute of Subtropical Forestry, CAF; Wetland Ecosystem Research Station of Hangzhou Bay, State Forestry Administration, Fuyang 311400 , China)
Waterbird surveys were conducted regularly in the Qiantangjiang River estuary and Hangzhou Bay from July 2007 to November 2011. A total of 128 species (nine orders and 18 families) were recorded, including 119 migrants which accounted for 93% of the total species; eleven species were listed as National Protected Species. Inter-specific correlation analysis for 13 shorebird populations and nine duck populations recorded over time found that 21 pairs of shorebirds and 23 pairs of ducks were correlated. By looking at seasonal dynamics and migration patterns we were able to divide the migration process into six stages: (1) late July to late September was the migration peak of shorebirds, which were dominated byLimosa limosa,Calidris ruficollisandCharadrius mongolus. (2) Early October to mid-December was the migration peak of wintering migrants of shorebirds and ducks, which were the first two large groups in our study areas. (3) Late December to mid-February was the wintering period of migration waterbirds. (4) Late February to late March was the peak migration of ducks and the winter migrants of shorebirds dominated byCalidris alpina.(5) Early April to mid-May was the migration peak of passage migrants such as,Calidris ruficollis, Calidris acuminate and Limosa limosabut the population size of shorebird winter migrants dominated byCalidris alpinewas still larger than the former. (6) Late May to mid-July was the breeding season of all egrets, summer migrants of gulls and several species of shorebirds. Our surveys show that interaction among species is possibly an important determinant of community composition of shorebirds and wintering ducks during the migration season. It may be the geographical position and community composition of migrant shorebirds across Hangzhou Bay that mean during the northward migration there are far more shorebirds than during the southward migration.
book=32,ebook=110
Migratory waterbirds; Interspecific correlation; Migration pattern
Q959.7;Q958.13
A
0254-5853-(2011)06-0631-10
10.3724/SP.J.1141.2011.06631
2011-06-02;接受日期:2011-10-09
浙江省基金項目“杭州灣濕地景觀格局變化對水鳥多樣性的影響”(Y5080130);國家“十一五”科技攻關專題項目“典型濕地可持續綜合利用技術試驗示范”(2006BAD03A1904);中國林科院亞熱帶林業研究所專項資金項目“杭州灣濕地生物多樣性監測及數據庫構建”(RISF6146)
E-mail: jiangky2005@126.com
book=32,ebook=118
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
學生天地(2019年36期)2019-08-25 08:59:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30

- Zoological Research的其它文章
- New details indicated by different stainings during conjugation of ciliated protozoa Paramecium
- Histological and immunocytochemical study of deferens ducts in the Chinese rat snake (Zaocys dhumnades)
- PTEN在小鼠卵細胞和早期胚胎發育中的功能研究
- A phylogeny of the Tylototriton asperrimus group (Caudata: Salamandridae) based on a mitochondrial study: suggestions for a taxonomic revision
- 云南新平哀牢山西黑冠長臂猿分布與群體數量
- 上海市南匯東灘圍墾后海岸帶濕地冬春季水鳥生境選擇