RNAi沉默PRDXⅢ基因表達誘導U251膠質瘤細胞凋亡的實驗研究
2012-01-20 09:02:26左書浩焦慶芳焦保華
河北醫藥 2012年16期
關鍵詞:檢測
左書浩 焦慶芳 焦保華
本文通過RNAi技術沉默人U251膠質母細胞瘤細胞系中PRDXⅢ的表達,探討其對膠質瘤細胞凋亡和生長的影響,為膠質瘤治療提供新的選擇。
1 材料與方法
1.1 材料 人腦膠質瘤U251細胞株購自ATCC公司,RPMI-1640培養基購自GIBCOBRL公司,RT-PCR二步法試劑盒購自MBI公司,LipofectamineTM 2000購自Invitrogen公司,Opti-MEM I Reduced Serum Medium購自Invitrogen公司,凋亡檢測試劑盒購自晶美公司,DNA Ladder試劑盒購自碧云天生物技術研究所。PRDXⅢ引物1對,內參照GAPDH引物1對,均由上海英駿生物技術有限公司合成。引物序列為:PRDXⅢ:上游引物:5’-CTTAAGAAGATGGCGGCTGCTGCTGTAGGAC-3’下游引物:5’-CTCGAGCATGGGTGATCTACTGATTTACCTT-3’GAPDH:上游引物:5’-ACCACATGCCCAGAGGGTCC-3’下游引物:5’-AGGAGGCTGGGCTGTCTGTA-3’基因特異干擾序列表達載體pGenesil-1-PRDXⅢ-1、pGenesil-1-PRDXⅢ-2、pGenesil-1-PRDXⅢ-3由中國武漢晶賽生物有限公司進行設計與合成。
1.2 方法
1.2.1 U251細胞培養、傳代與保種:U251細胞用RPMI-1640培養液,置于37℃、5%CO2培養箱培養,每2~3天換液1次,培養細胞生長至80%融合。加入新鮮的0.25%胰酶消化細胞,離心沉淀,然后分置細胞于新的培養瓶中,傳代比例為1∶3。最后按照每個凍存管2×106個細胞保種。細胞梯度冷凍,最后置于液氮罐中保存。
1.2.2 細胞轉染:以0.25%胰酶消化收集細胞,轉種于6孔板,每孔1×105個細胞,加入2 ml含小牛血清和抗生素的培養基繼續孵育。當細胞達到80%的融合時移去6孔板內完全培養基,加入1 ml無血清無抗生素培養基,加入轉染復合物(含質粒及脂質體),混勻,培養4~6 h后,換有血清有抗生素的完全培養基2 ml。轉染24 h后,熒光顯微鏡下觀察6孔板內熒光以檢測基因轉染效率。以未轉染的空白對照組,空載體轉染組,無關序列轉染組為對照組。
1.2.3 PRDXⅢ的分離、擴增:按照Trizol試劑說明書步驟一步法提取總 RNA。合成 cDNA,通過 RT-PCR法擴增 PRDXⅢ(95℃預變性 3 min,95℃變性 30 s,61.4℃退火 35 s,72℃延伸1 min,共33個循環,最后72℃延伸10 min)和內參基因GAPDH(95℃預變性 5 min 后,95℃變性45 s,56℃退火40 s,72℃延伸40 s,共35個循環后,72℃延伸10 min終止反應),篩選有效的基因特異干擾序列。
1.2.4 Western blot檢測膠質瘤細胞中PRDXⅢ蛋白的表達:轉染后48 h,以含0.25%胰蛋白酶消化收集細胞(細胞經預冷的PBS漂洗3次);加入450 μl RIPA裂解液裂解細胞(冰上操作),在振蕩器上混勻4~15 min,14 000 g離心15 min(4℃),棄沉淀;處理后的樣品加入樣品緩沖液,煮沸5 min,12 000 g離心5 min,取上清液,用分光光度計對蛋白質進行定量檢測。將蛋白樣本進行凝膠電泳,然后電轉移至PVDF膜。TBS洗膜3次后裝入密封袋,加入1∶4 000稀釋的一抗,4°C過夜,緩慢搖動。棄一抗,TBS/T緩沖液室溫潤洗,5 min×4次。裝入新的密封袋,加入1∶5 000稀釋的的二抗,室溫孵育1 h,緩慢搖動。棄二抗,TBS/T緩沖液室溫潤洗,10 min×3次,浸泡于TBS中待檢測。加入DAB顯色液,顯色5 min。以β-actin為內參,其一抗稀釋度為1∶3 000。在凝膠成像系統上分析處理蛋白條帶,拍照。以PRDXⅢ蛋白條帶OD值與內參照OD值的比值來表示蛋白表達的量。
1.2.5 DNA Ladder:每 106個細胞中加入 500 μl添加了蛋白酶K的樣品裂解液,Vortex混勻,充分裂解細胞,50℃水浴消化過夜。加入500 μl Tris平衡苯酚,劇烈震蕩10~30 s,以使酚相和水相充分相互作用以抽提樣品。吸出約300 μl上清液,加入60 μl 10 mol/L醋酸銨和600 μl無水乙醇,顛倒數次混勻,此時可見DNA沉淀產生,-20℃凍存1 h,以充分沉淀小片斷DNA。12 000 g離心10 min,棄上清,加入70%乙醇洗滌 DNA沉淀1次。盡量吸除殘余的乙醇,加入50~100 μl TE溶解DNA,取部分抽提得到的基因組DNA,瓊脂糖凝膠電泳分析。
1.2.6 PI染色:轉染后48 h PI染色觀察細胞凋亡。將蓋玻片取出,滴上1滴PI液,熒光顯微鏡下觀察紅色熒光。實驗分組同上:干擾質粒實驗組,無關序列對照組,空白載體對照組及未處理的空白對照組。
1.2.7 流式細胞儀(FCM)檢測細胞凋亡:轉染后48 h收集細胞約5 ×106個,離心,1 000 r/min,5 min,棄去培養液;PBS洗滌2次;離心,1 000 r/min,5 min,去上清液;加入冰預冷的70%的乙醇固定,4℃,1 ~2 h;離心,1 000 r/min,5 min,棄去固定液;PBS重懸細胞,400目的篩網過濾1次,1 000 r/min,離心5 min,棄去PBS;用1 ml PI染液染色,4℃避光30 min;流式細胞儀檢測。
1.2.8 MTT比色分析轉染后細胞增殖活性:將U251細胞接種到96孔板,每孔2×103細胞,待細胞達80%融合后,選擇基因特異干擾序列3由脂質體包裹進行轉染。轉染后48 h,每孔加入10 ~15 μl MTT(5mg/ml),5%CO2培養箱中孵育4 h,將孔內液體全部吸去,每孔加入100 μl DMSO,10 min后用酶標儀在490 nm波長處讀取吸收值。計算干擾后對細胞的生存抑制率。以空白組、空載體轉染組和無關序列轉染組作對照。
1.3 統計學分析應用SPSS 11.5統計軟件,計量資料以±s表示,采用單因素方差分析,P<0.05為差異有統計學意義。
2 結果
2.1 RT-PCR分析轉染后PRDXⅢmRNA水平變化 半定量RT-PCR檢測示,空載體組和無關序列組PRDXⅢmRNA表達無明顯變化,干擾序列1、3使PRDXⅢmRNA的表達明顯降低,尤以干擾序列3的效果最顯著,對PRDXⅢmRNA的表達抑制率為76%。見表1、圖1。
表1 干擾后4組PRDXⅢmRNA表達水平的變化±s

表1 干擾后4組PRDXⅢmRNA表達水平的變化±s
注:與空白對照比較,*P <0.05
類別 PRDXⅢ/GAPDH 抑制率(%) P值0.84 ±0.025 0.00干擾序列1 0.26 ±0.024* 69干擾序列2 0.68 ±0.020 19干擾序列3 0.20 ±0.047*空白對照76
2.2 Western Blotting分析轉染后PRDXⅢ蛋白表達變化Western Blotting結果顯示,空載體組和無關序列組PRDXⅢ蛋白表達無明顯變化,干擾序列1、3對PRDXⅢ蛋白表達有明顯抑制作用,以干擾序列3作用最顯著,抑制率達到63.9%。見表 2、圖 2。

圖1 PRDXⅢ基因有效特異干擾序列篩選分析
表2 干擾后各組PRDXⅢ蛋白表達水平±s

表2 干擾后各組PRDXⅢ蛋白表達水平±s
注:與空白對照比較,*P <0.05
類別 PRDXⅢ/β-actin 抑制率(%) P值0.72 ±0.015 0.00干擾序列1 0.33 ±0.020* 54.2干擾序列2 0.56 ±0.026 22.2干擾序列3 0.26 ±0.020*空白對照63.9

圖2 Western Blotting檢測干擾后PRDXⅢ蛋白表達
2.3 DNA ladder 細胞凋亡時主要的生化特征是其染色質發生濃縮,染色質DNA在核小體單位之間的連接處斷裂,形成180-200bp整數倍的寡核苷酸片段,在凝膠電泳上表現為梯形電泳圖譜(DNA ladder)。試驗結果顯示干擾組出現明顯梯形電泳條帶,說明有較明顯的細胞凋亡,而空白對照組,空載體和無關序列組未見梯形電泳條帶。見圖3。

圖3 DNA ladder顯示干擾組DNA梯形電泳條帶
2.4 PI染色 空白對照組,空脂質體組和空載體組細胞未見明顯凋亡;而干擾組細胞染色質出現濃縮狀態,細胞核的染色質高度凝聚,部分細胞核裂解為碎塊,產生凋亡小體,符合凋亡的形態學改變。見圖4。
2.5 流式細胞儀檢測細胞凋亡 流式細胞儀檢測顯示空白對照組、空載體組、無關序列組和干擾組細胞凋亡率分別為(2.3±0.6)%、(2.7 ±0.7)%、(3.5 ±0.6)%和(35.9 ±4.8)%,干擾組細胞調亡率明顯高于空白對照組、空載體組、無關序列組(F=219.76,P <0.01),而空白對照、空載體組和無關序列組之間促凋亡率的差異無統計學意義(P<0.05)。見圖5。
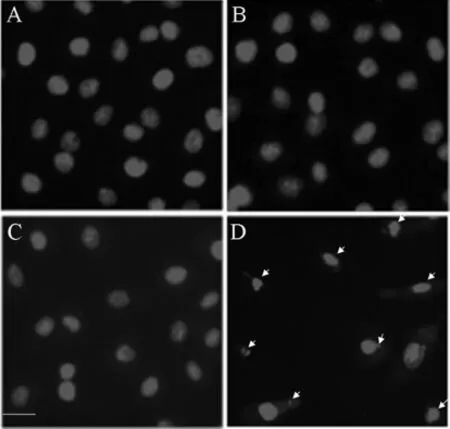
圖4 PI染色

圖5 FCM檢測干擾后48 hU251細胞凋亡率
2.6 MTT檢測RNAi后U251細胞生長活性 MTT比色實驗結果顯示空白對照組、空載體組、無關序列組和干擾組轉染后48小時的光吸收值分別為(1.26 ±0.06)、(1.22 ±0.09)、(1.19 ±0.02)和(0.70 ±0.07),干擾組光吸收值明顯低于空白對照組、空載體組和無關序列組,而空白對照組、空載體組和無關序列組相互間差異無統計學意義(P>0.05)。干擾組對U251細胞生長的抑制率為44.4%,明顯減慢了細胞生長(F=49.9,P <0.01)。
3 討論
RNA 干擾(RNA interference,RNAi)[1]是雙鏈 RNA(double-stranded RNA,dsRNA)介導的特異性基因表達沉默現象,是指當細胞中導入與內源性mRNA編碼區同源的dsRNAs時,該mRNA發生降解而導致基因表達沉默的現象。由于這種現象發生在轉錄后水平,故又稱為轉錄后基因沉默(post transcriptional gene silencing,PTGS)。
RNAi包含至少下列幾個連續的步驟[2]:(1)外源性或內源性的dsRNA在細胞內與一種RNA酶Ⅲ(RNAⅢendonucleasedicer)結合,隨即被切割成含有3’端單鏈尾巴及磷酸化的5’端的21-23nt的短鏈 dsRNA,是 RNAi的起始誘導物,即 siRNA[3]。(2)siRNA與相關蛋白形成RNA引導的沉默復合體(RNA induced silencing complex,RISC)[4],RISC 具有核酸酶活性,它可以在siRNA的引導下特異降解相應的mRNA。(3)siRNA按照堿基互補原則識別靶基因轉錄出的mRNA,并引導RICS復合體結合mRNA。(4)siRNA與mRNA在復合體中換位,核酸酶Dicer將mRNA切割成21-23nt的片斷,特異性地抑制靶基因地表達。而新形成的dsRNA可再次形成RISC復合體,繼續降解mRNA,從而產生級聯放大效應。因此,每個細胞僅需要幾個siRNA就能引起強烈的RNAi反應。siRNA還可以在RNA依賴性RNA聚合酶(RNA dependent RNA polymerase,RdRp)的作用下大量擴增,并轉運出細胞,使RNA擴散到整個機體并可以傳代[5]。最近的研究也有顯示,RNAi不僅在轉錄后水平發揮作用,而且在轉錄水平和翻譯水平也有一定程度的抑制特異基因表達的作用。針對靶基因設計出特異性的具有高效抑制作用的siRNA序列是RNAi成功的關鍵。在siRNA序列的選擇上應注意:(1)siRNA序列的長度。序列太短則不足以誘導特異性的基因抑制,太長又會激活哺乳動物細胞的抗病毒防御機制。21nt的siRNA既可在哺乳動物細胞介導基因特異性的抑制作用,又不會引起宿主細胞非特異性的抗病毒反應,所以目前多數的siRNA是21nt的雙鏈。另外,在3’對稱懸掛2nt的堿基(通常為UU)的siRNA降解靶基因mRNA最為有效[6],尤其是在反義鏈3’懸掛2ntUU對誘導哺乳動物細胞中有效的RNAi尤為重要[7]。(2)靶序列的選擇。在目標分子的作用部位選擇上,一般認為在cDNA的轉錄起始位置下游50~100 bp處較好,要避開轉錄的非翻譯區。在靶基因的成熟 mRNA序列AUG起始密碼子后尋找到以“AA”開頭的序列,再向下游取19nt作為正義序列。(3)干擾分子的GC含量一般在30% ~70%,最好50%左右。我們按照上述原則設計了3條siRNA序列,利用真核表達載體轉染U251細胞。轉染后24和48 h分別以RT-PCR和Westtern blotting檢測PRDXⅢmRNA和蛋白的表達,發現第1、3條siRNA序列較好地抑制了PRDXⅢmRNA和蛋白的表達,尤以第3條最佳,對PRDXⅢmRNA和蛋白表達的抑制率分別達到76%和63.9%。在后續的實驗中我們利用第3條干擾序列進行進一步的凋亡研究。PRDXⅢ與細胞凋亡有關,PRDXⅢ過表達可以抑制藥物誘導的凋亡,而降低PRDXⅢ表達使得細胞對凋亡更加敏感[8,9]。我們利用RNAi技術成功地抑制了U251細胞中PRDXⅢ基因和蛋白的表達,轉染后48 h流式細胞儀檢測,干擾組細胞凋亡率明顯高于對照組,差異有統計學意義;DNA ladder顯示干擾組有凋亡典型的梯形條帶,而各對照組未見此現象;PI染色顯示干擾組細胞染色質出現濃縮狀態,細胞核的染色質高度濃染,部分細胞核裂解為碎塊,符合凋亡的形態學改變;MTT顯示干擾組對U251細胞生長的抑制率為44.4%,明顯減慢了細胞生長,而無關序列組、空載體組和空白對照組之間細胞生長無明顯差異。這些結果表明,PRDXⅢ基因在膠質瘤中高表達可以抑制腫瘤細胞凋亡,從而使其增殖加快,而抑制PRDXⅢ基因表達能夠誘導膠質瘤凋亡,減緩腫瘤生長,因此,PRDXⅢ基因有可能成為膠質瘤基因治療的靶基因,為膠質瘤治療提供了一個新的選擇。
1 Hannon GJ.RNA interference.Nature,2002,418:244.
2 Nykanen A,Haley B,Zamore PD.ATP requirements and small interfering RNA structure in the RNA interference pathway.Cell,2001,107:309.
3 Zamore PD,Tuschl T,Sharp PA,et al.RNAi:double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals.Cell,2000,101:25.
4 Bass BL.Double-stranded RNA asa template for gene silencing.Cell,2000,101:235.
5 Dalmay T,Hamilton A,Rudd S,et al.An RNA-dependent RNA polymerase gene in Arabidopsis is required for-transcriptional gene silencing mediated by a transgene but not by a virus.Cell,2000,101:543.
6 Elbashir SM,Harborth J,Weber K,et al.Analysis of gene function in somatic mammalian cells using small interfering RNAs.Method,2002,26:199.
7 Hohjoh H.RNA interference(RNAi)induction with various types of synthetic oligonucleotide duplexes in cultured human cells.FEBS Letters,2002,521:195.
8 Nonn L,Berggren M,Powis G.Increased Expression of Mitochondrial Peroxiredoxin-3(Thioredoxin Peroxidase-2)Protects Cancer Cells Against Hypoxia and Drug-Induced Hydrogen Peroxide-Dependent Apoptosis.Molecular Cancer Research,2003,1:682.
9 Chang TS,Cho CS,Park S,et al.Peroxiredoxin III,a Mitochondrion-specific Peroxidase,Regulates Apoptotic Signaling by Mitochondria.The Journal of Biological Chemistry,2004,279:41975.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
