中國濃香型白酒窖池糟醅中微生物群落演替分析
2012-06-01 09:25:08張大鳳劉若塵邢亞閣李明元車振明向文良
食品科學 2012年15期
關鍵詞:生態
張大鳳,李 可,劉 森,劉若塵,邢亞閣,李明元,車振明,向文良,*
(1.西華大學生物工程學院 四川省食品生物技術重點實驗室,四川 成都 610039;2.西華大學古法發酵(釀造)生物技術研究所,四川 成都 610039)
中國濃香型白酒窖池糟醅中微生物群落演替分析
張大鳳1,2,李 可1,2,劉 森1,2,劉若塵1,邢亞閣1,李明元1,2,車振明1,向文良1,2,*
(1.西華大學生物工程學院 四川省食品生物技術重點實驗室,四川 成都 610039;2.西華大學古法發酵(釀造)生物技術研究所,四川 成都 610039)
為探索中國濃香型白酒發酵糟醅中微生物群落演替規律,利用克隆分析,從時間和空間兩個層面對糟醅微生物區系的消長變化進行較為全面的調查。根據有效克隆子分布數據計算的群落多樣性參數表明:窖池糟醅微生物群落多樣性在時間和空間上存在差異。應用CANOCO 4.5 軟件對物種和生態因子進行典范對應分析(CCA),應用CANODRAW 4.0作種類、樣方分布與生態因子關系的二維排序圖,排序圖上清楚地反映了窖池糟醅微生物種類、群落分布與生態因子的關系。結果表明:酸度、乙醇含量、水分含量是影響微生物群落結構組成的主要微生態因子;大部分微生物適宜在低酸度條件下生長,少數耐酸微生物能在高酸度條件下生長;隨著發酵的進行,生態因子發生變化,微生物群落也進行相應的演替,向耐高酒精含量、高酸度的群落發展。
中國濃香型白酒;發酵糟醅;微生物演替
中國濃香型白酒的生產以泥窖窖池為基礎,發酵過程是棲息在窖池糟醅、窖泥中的龐大微生物區系在糟醅固、液、氣三相界面的復雜的物質能量代謝過程[1-2]。在這一過程中,窖池生態因子如窖池溫度、糟醅酸度、總糖及還原糖含量、糟醅水分含量等與窖池中的微生物彼此聯系、互相促進、互相制約、協調發展,共同構成了窖池特定的微生物生態系統,并在“三流運轉”規律的交互作用下構成了發酵過程中窖池微生態的動態發展和特定階段的動態平衡[3-4]。由于環境信息的傳遞對于調控窖池微生物種類的變化、代謝產物的生成與積累至關重要,并決定著窖池物質代謝和能量代謝的走向。因此,環境生態因子的變化對微生物的生長、代謝影響非常大。在窖池不同空間位置和微生物生長代謝的不同階段,微生物對窖池生態因子的需求都有所不同[5]。隨著發酵的進行,微生物的演替活動將引起窖池環境生態因子不斷發生變化,而這些變化又將反過來作用于窖池微生物,從而影響窖池發酵,最終導致窖池中各代謝產物成分和比例的變化,從而影響窖池濃香型白酒的產量及品質[6-7]。所以,研究窖池微生物的動態變化對于把握窖池發酵情況是很有必要。正因如此,白酒窖池生態系統中微生物的演替引起了廣大學者的極大興趣,但系統分析其演替過程,還未見報道。
本研究從生態學角度審視傳統固態發酵,利用生態分析相關軟件,對中國白酒窖池糟醅中微生物群落和生態因子等相關數據進行數理統計,以揭示伴隨發酵進行,窖池不同空間層次糟醅中微生物群落物種多樣性變化趨勢和演替規律以及在窖池不同空間層次和不同發酵時間的糟醅中微生物群落結構與窖池微生態因子之間的關系,尋找對糟醅微生物群落結構起關鍵作用的生態因子,探討影響微生物群落演替的機制,為理解中國濃香型白酒發酵機理、特征風味形成機制提供參考。
1 材料與方法
1.1 數據采集
分析所用的微生物群落和糟醅生態因子數據均由西華大學古法發酵(釀造)生物技術研究所提供。樣品采集于四川某著名濃香型白酒企業48號窖池,分別為窖池上、中、下層中心發酵時間1、7、6 0 d的糟醅。其中,上層糟醅樣分別記為:T01、T07、T60;中層糟醅樣分別記為:M01、M07、M60;下層糟醅樣分別記為:B01、B07、B60。所有樣品中的pH值、酸度、還原糖、總糖、乙醇、溫度、水分、乙醛、酯和蛋白質等微生態因子依據沈怡方等[8]的相關方法測定。糟醅中微生物群落的分析參考向文良[7]、Zhang Wenxue[9]等的方法,PCR分別擴增原核和真核微生物的16S rRNA基因和18S rRNA基因。擴增的16S rRNA基因和18S rRNA基因再分別與pGM-T載體連接成重組質粒后,分別導入感受態的E.coli DH5α構建窖池糟醅中原核微生物的16S rRNA基因文庫和真核微生物的18S rRNA基因文庫,提取重組質粒,基因測序分析窖池糟醅中微生物的分類學地位。窖池糟醅的生態因子數據見表1。窖池糟醅中微生物群落數據見表2,其中原核微生物代表克隆子的16S rRNA基因在GenBank(http://www. ncbi.nlm.nih.gov/genbank/)中的接受號:JQ693078~JQ693100;真核微生物代表克隆子的18S rRNA基因在GenBank中的接受號:JQ693061~JQ693077。
1.2 數據分析方法
微生物群落關系的演替關系利用Origin8.0繪制。群落多樣性參數Simpson指數(D)、Shannon-Wiener多樣性指數(H)、均勻度(J)的計算公式分別如下:

式中:pi為第i種物種個體數占群落總個體數的比例;S為物種總數;Hmax為H的最大值,等于lnS[10]。Shannon-Wiener 多樣性指數隨時間的動態變化圖用SPSS for Windows 13.0繪制。窖池糟醅中微生物群落與環境的相互作用關系利用典范對應分析(canonical correspondence analysis, CCA)排序[11]。
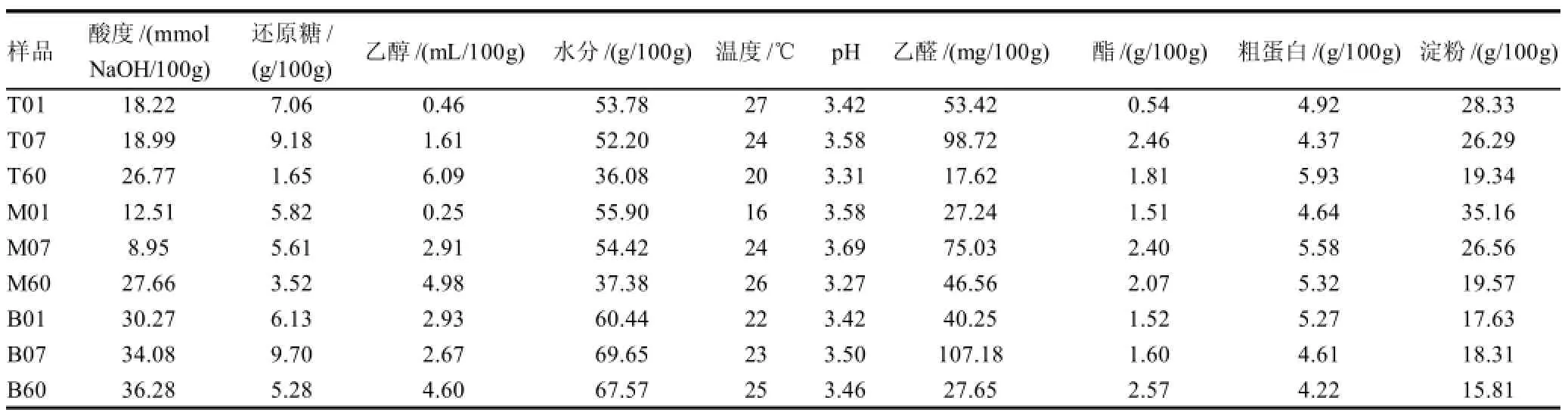
表1 窖池糟醅的生態因子Table 1 Chemical composition of lees (wet weight basis)

表2 窖池糟醅樣中微生物16S rRNA和18S rRNA基因的克隆分布Table 2 Microbial categorization and abundance in fermentation lees based on the 16S rRNA and 18S rRNA analysis
2 結果與分析
2.116 S rRNA和18S rRNA基因的克
隆分析
2.1.1 上層糟醅中微生物分析
基于16S rRNA和18S rRNA分析技術表明:入窖樣獲得了22個有效克隆,發酵7d樣獲得了53個有效克隆,發酵60d樣獲得了42個有效克隆,這些克隆子代表的微生物伴隨發酵時間發生的群落演替如圖1所示。上層糟醅對原核微生物而言,入窖樣中未檢出古細菌,發酵7d樣和6 0 d樣檢出的古菌為M e t h a n o c u l l e u s屬和Methanospirillum屬;其中Methanospirillum屬只在發酵60d樣被檢出。入窖樣中,細菌、霉菌、酵母的種類和數量差異不是很大。隨著發酵進行和糟醅理化因子的變化,糟醅中微生物群落也發生明顯變化,特別是細菌,其種類和數量發生明顯變化(圖2)。入窖樣中未出現的Streptococcus 屬、Acetobacter屬、Arthrobacter屬、Bacillus屬、Microbacterium屬、Erwinia屬和Rhodococcus屬均在發酵7d樣中檢出。在60d樣中,已檢測不到酵母了,因為這時發酵已結束,此時窖池環境惡劣,如乙醇含量度高、嚴格厭氧、酸度高,酵母無法生存[12-13],此時在60d樣品中只有甲烷菌、霉菌、細菌。

圖1 窖池上層糟醅中微生物16S rRNA和18S rRNA基因的克隆分布圖Fig.1 Microbial categorization and abundance in the top layer of fermented lees based on the 16S rRNA and 18S rRNA analysis
2.1.2 中層糟醅中微生物分析
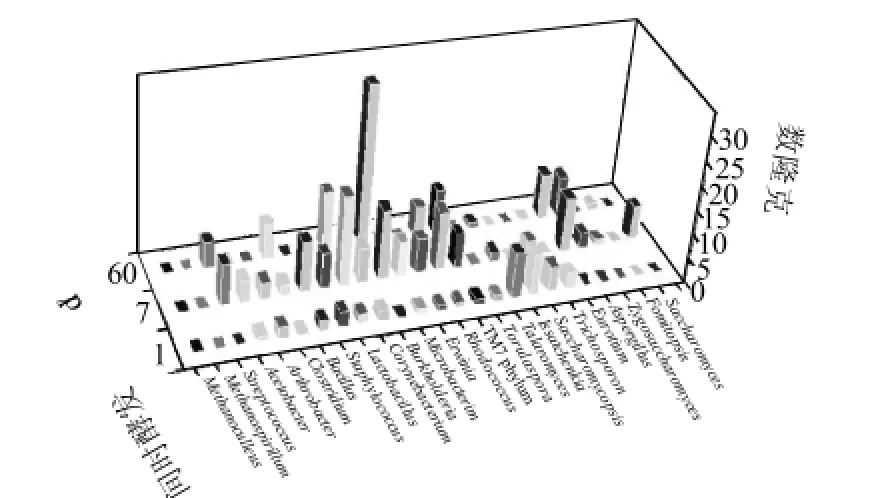
圖2 窖池中層糟醅中微生物16S rRNA和18S rRNA基因的克隆分布圖Fig.2 Microbial categorization and abundance in the middle layer of fermented lees based on the 16S rRNA and 18S rRNA analysis
中層樣主要是母糟,包括紅糟和糧糟,是發酵最活躍的區域[14],微生物的種類和數量在此糟醅中發生劇烈演替。由圖2可知,入窖樣獲得了42個有效克隆,發酵7d樣獲得了133個有效克隆,發酵60d樣獲得了90個有效克隆,其數量明顯多于上層樣和下層樣。上層中的Methanoculleus屬和Methanospirillum屬在中層中沒有檢出,其他古菌也未檢出。Burkholderia屬在上層樣中演替并不活躍,但中層糟醅中盡管入窖時中未檢出,但是當其在發酵7d時Burkholderia屬的克隆數迅速達到14個,在發酵60d時其進一步上升,達到32個克隆,占發酵60d總克隆數的35.6%,此時Burkholderia屬成為優勢屬。Talaromyces屬在上層樣中比較活躍,一度成為上層發酵7d糟醅中的優勢菌。在中層糟醅中隨著發酵的進行,Talaromyces屬中微生物數量一直遞減,在發酵7d樣和60d樣中分別只能檢測出2個和1個克隆子。Eurotium 屬、Aspergillus屬在中層中含量也比較豐富,這與中層樣中含有的糧糟有關。同樣,在中層糟醅發酵60d時也未檢出酵母。
2.1.3 下層糟醅中微生物分析

圖3 窖池下層糟醅中微生物16S rRNA和18S rRNA基因的克隆分布圖Fig.3 Microbial categorization and abundance in the bottom layer of fermented lees based on the 16S rRNA and 18S rRNA analysis
由圖3可知,在下層糟醅中入窖樣獲得了27個有效克隆,發酵7d樣獲得了44個有效克隆,發酵60d樣獲得了38個有效克隆。下層樣是多輪底糟,其酸度非常高,所以微生物種類和數量都相對較少,在發酵過程中微生物種類和數量的波動比較平緩。下層樣另一特點是古菌含量豐富,包括甲烷菌M e t h a n o c u l l e u s屬和Methanospirillum屬,入窖糟醅中古菌占總菌屬的29.6%,7d為29.5%,60d為57.9%。發酵結束后,只有幾種耐酸性的細菌存在,主要為細菌中的Lactobacillus屬和TM7 phylum;霉菌和酵母在此時未被檢出。
2.2 窖池中微生物群落多樣性分析

圖4 Shannon-Wiener多樣性指數隨發酵時間的變化Fig.4 Temporal changes of microbial Shannon-Wiener diversity
窖池糟醅上、中、下層的入窖樣、發酵7d樣、發酵60d樣共9個樣方中分布的微生物菌群多樣性分析結果見圖4。Shannon-Wiener多樣性指數在上、中、下層糟醅中表現出同樣的變化趨勢:隨著發酵的開始,群落多樣性增加,到發酵結束時,群落多樣性降低。發酵開始時,糟醅原料提供了豐富的營養物質和適宜的生長條件,各類微生物能快速生長。隨著氧氣和營養物質的消耗,代謝產物的積累,窖池環境變得越來越不適宜微生物的生長。下層樣由于酸度最高,微生物種類最少,主要是一些耐酸的微生物能生長,故其Shannon-Wiener多樣性指數在入窖樣、發酵7d樣、發酵60d樣中均低于上層樣和中層樣。中層樣是發酵最活躍的區域,也是微生物種類和數量最豐富的區域,其多樣性指數在入窖樣和發酵7d樣中均大于上層樣和下層樣。但中層樣在發酵60d樣中卻低于上層樣,可能是由于中層樣與下層樣間通過黃水的物質交換比較頻繁,中層發酵60d樣中的酸度也比較高,導致微生物多樣性低于上層樣。
2.3 微生物群落演替與生態因子之間的關系

表3 窖池糟醅的微生態因子Table 3 Microecological factors in fermentation less

表4 CCA分析的統計信息Table 4 Statistical analysis of CCA analysis results
9個樣方的生態因子經過處理后(表3)與微生物的種類用于CCA排序。CCA排序結果(表4)表明:軸1和軸2兩個排序軸的本征值分別為0.398和0.296;生態因子與微生物種類相關性分別為0.998和0.868;物種數據的方差累計百分數分別為30.7和53.6;總本征值為1.294,總典范本征值為1.036,解釋量為80.06%;蒙特卡羅檢驗值小于等于0.05,所以排序結果較好的反映了微生物種類組成與生態因子間的相互關系[15-16]。從CCA排序圖來看,酸度梯度是所有因子中對微生物群落分布起決定性作用的生態因子,大多數物種都分布在低酸度區域,如圖5所示。低酸度區域的物種多樣性遠遠高于高酸度區域[17]。Methanoculleus(1)和Methanospirillum(2)屬對酸度的要求高,是糟醅中最耐酸的微生物,且對水分的要求也高。TM7 phylum(15)、Lactobacillus(9)和Clostridium(6)屬也能耐一定的酸度。Trichosporon(18)、Saccharomycopsis(17)、Torulaspora(23)和Issatchenkia(24)屬主要分布在還原糖/粗蛋白質比值高、水分含量較高、乙醇含量低、低酸度、相對溫度較低的區域,其中Tri-chosporon(18)屬對水分的要求較高 。Talaromyces(16)、Staphylococcus(8)、Arthrobacter(5)、Acetobacter(4)、Bacillus (7)和Corynebacterium(10)屬,多見于具有適宜的水分含量、合適的還原糖/粗蛋白質和低酸度的區域。Sa ccharomyces(25)、Microbacterium(12)、Erwinia(13)、Zygosaccharomyces(21)、Rhodococcus(14)和Streptococcus (3)屬,主要分布在適宜在水分含量低、溫度較高、酸度低和一定含量酒精的糟醅中。Fomitopsis(22)、Aspergillus(20)、Burkholderia(11)、Eurotium(19)和Clostridium(6)屬的微生物與糟醅乙醇含量有著極大地相關性,在較高乙醇含量條件下也能生存。

圖5 微生物物種與生態因子的CCA二維排序Fig.5 Canonical correspondence analysis based on microbial species and ecological factors
3 結 論
3.1 統計分析表明,在發酵不同時間、窖池糟醅不同空間層次中微生物類群和數量存在較大的差異,以中層樣居首。
3.2 通過對多樣性分析表明,窖池糟醅微生物群落多樣性隨空間和發酵時間的變化而異,具體表現為:入窖樣和發酵7d樣的群落多樣性分別為中層>上層>下層,發酵60d樣的群落多樣性為上層>中層>下層。在時間軸上,糟醅微生物群落多樣性呈先上升后下降的趨勢。
3.3 CCA排序結果顯示:酸度、乙醇含量、水分含量是影響微生物群落結構組成的主要微生態因子。除了極少數耐酸的微生物,如甲烷菌,Lactobacillus屬能在較高的酸度下生存,大部分微生物生活在低酸度區域。隨著發酵的進行,乙醇逐漸積累,微生物群落發生相應的演替,向耐高酒精含量、高酸度的群落發展。
[1]張文學, 喬宗偉, 向文良, 等. 中國濃香型白酒窖池微生物生態研究進展[J]. 釀酒, 2004, 31(2): 31-35.
[2]趙發清, 馬海燕. 微生態基本概念的剖析[J]. 中國微生態學雜志, 1997, 7(6): 41-44.
[3]羅海, 劉森, 向文良, 等. 濃香型白酒窖池微生物生態因子的動力學分析[J]. 中國釀造, 2011(7): 108-112.
[4]唐由凱. 論微生態系統及系統分析[J]. 中國微生態學雜志, 2000, 12 (2): 116-118.
[5]張文學. PCR技術對濃香型白酒糟醅細菌菌群的解析[J]. 四川大學學報: 工程科學版, 2005, 37(5): 82-87.
[6]王大珍. 微生物生態學的發展及應用[J]. 科學, 1993(2): 18-20.
[7]向文良, 喬宗偉, 張文學, 等. 中國濃香型白酒窖池中原核微生物的特性及系統發育分析[J]. 四川大學學報: 工程科學版, 2005, 37(1): 39-42.
[8]沈怡方, 高景炎, 康明官, 等. 白酒生產技術大全[M]. 北京: 中國輕工業出版社, 2007: 597-671.
[9]ZHANG Wenxue, QIAO Zongwei, TANG Yueqin, et al. Analysis of the fungal community in Zaopei during the production of Chinese Luzhou flavour liquor[J]. Journal of the Institute of Brewing, 2007, 113(1): 21-27.
[10]KEYLOCK C J. Simpson diversity and the Shannon-Wiener index as special cases of a generalized entropy[J]. Oikos, 2005, 109(1): 203-207.
[11]TERAHARA T, IKEDAB S, NORITAKE C, et al. Molecular diversity of bacterial chitinases in arable soils and the effects of environmental factors on the chitinolytic bacterial community[J]. Soil Biol Biochem, 2009, 41(3): 473-480.
[12]向文良. 中國濃香型白酒窖池微生物生態研究[D]. 成都: 四川大學, 2004.
[13]施安輝. 濃香型白酒發酵過程窖中微生物區系的分析[J]. 釀酒, 1986 (4): 24-29.
[14]吳衍庸. 白酒工業生態中的微生物生態學[J]. 釀酒科技, 2001(5): 32-33.
[15]張金屯. 數量生態學[M]. 北京: 科學出版社, 2003: 131-193.
[16]郝占慶. 長白山北坡木本植物分布與環境關系的典范對應分析[J]. 植物生態學報, 2003, 27(6): 733-741.
[17]MACDONALD C A, THOMAS N, ROBINSON L, et al. Physiological, biochemical and molecular responses of the soil microbial community after afforestation of pastures with Pinus radiata[J]. Soil Biol Biochem, 2009, 41(8): 1642-1651.
Microbial Community Succession of Chinese Luzhou-Flavor Liquor Lees
ZHANG Da-feng1,2,LI Ke1,2,LIU Sen1,2,LIU Ruo-chen1,XING Ya-ge1,LI Ming-yuan1,2,CHE Zhen-ming1,XIANG Wen-liang1,2,*
(1. Provincial Key Laboratory of Food Biotechnology of Sichuan, College of Bioengineering, Xihua University, Chengdu 610039, China;2. Biotechnology Institute of Ancient Brewing, Xihua University, Chengdu 610039, China)
In order to explore the succession of microorganism communities in fermented grains from Chinese Luzhou-flavor liquor production, we investigated the temporal and spatial changes of microbial communities by clone analysis. The community diversity parameters calculated based on clone distribution suggested that microbial community diversity showed temporal and spatial variations. The data of species and ecological factors were analyzed by canonical correspondence analysis (CCA) using CANOCO 4.5. The species (or sites)-environment biplot of CCA were automatically mapped using CANODRAW 4.0 and the relationships of the ecological distribution of the fermented grains microorganism species and communities with the ecological factors were clearly revealed on these biplots. The results showed that the acidity, ethanol and humidity had important influences on the microbial community composition. The bulk of microorganism species adapted to grow in low acidity condition and only a few microorganisms could live in high acidity condition. The microorganism communities responded to changes of ecological factors along with fermentation, towards a community tolerant of high levels of ethanol and acidity.
Chinese Luzhou-flavor liquor;fermentation lees;community succession
TS261.1
A
1002-6630(2012)15-0183-05
2011-11-11
教育部春暉計劃項目(Z2010101);四川省食品生物技術重點實驗室開放基金項目(SZJJ2009-014);
西華大學重點科研基金項目(Z0910501)
張大鳳(1973—),女,講師,碩士研究生,研究方向為傳統發酵食品生物技術。E-mail:yhf1998914@163.com
*通信作者:向文良(1973—),男,副教授,博士,研究方向為傳統發酵食品微生物過程學。E-mail:biounicom@mail.edu.cn
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
貴茶(2019年3期)2019-12-02 01:46:32
鄉村地理(2018年3期)2018-11-06 06:51:02
茶葉通訊(2017年2期)2017-07-18 11:38:40
河北城市研究(2015年4期)2015-08-23 11:53:06
浙江工商大學學報(2015年6期)2015-03-01 02:56:40
