魚露中生物胺降解菌的篩選及其特性
2012-06-01 10:42:34楊利昆付湘晉胡葉碧周其中
食品科學 2012年11期
關鍵詞:生物
楊利昆,付湘晉,*,胡葉碧,周其中,李 濱
(1.中南林業科技大學 糧油深加工與品質控制湖南省重點實驗室,湖南 長沙 410004;2.中南林業科技大學食品科學與工程學院,湖南 長沙 410004)
魚露中生物胺降解菌的篩選及其特性
楊利昆1,2,付湘晉1,2,*,胡葉碧1,2,周其中2,李 濱1,2
(1.中南林業科技大學 糧油深加工與品質控制湖南省重點實驗室,湖南 長沙 410004;2.中南林業科技大學食品科學與工程學院,湖南 長沙 410004)
從天然發酵魚露中篩選具有生物胺降解活性的微生物,并研究發酵時間和發酵溫度對其降解生物胺的影響。以天然發酵3個月的魚露為分離材料,對魚露中的生物胺降解菌進行分離純化,獲得10株具有生物胺降解活性的微生物。分離到的10株菌株在30℃發酵72h組胺降解率在17.3%~62.2%,酪胺降解率在12.4%~57.3%。其中M8菌株生物胺降解活性最高,經26S rDNA測序及在線序列比對,M8菌株鑒定為奧默柯達酵母。奧默柯達酵母M8在30℃條件下降解生物胺活性最強;其降解生物胺的時間變化規律為發酵第1天生物胺含量降低,第2天和第3天生物胺含量升高,第4天開始生物胺含量持續降低,30℃發酵9d后,組胺和酪胺降解率分別為69.6%和79.2%。奧默柯達酵母M8在25%食鹽條件下生物胺降解效果最好,組胺和酪胺的降解率分別為64.4%和72.4%。
魚露;生物胺;奧默柯達酵母
魚露,又稱魚醬油,是一種風味獨特的水產調味品,其味咸、極鮮美、營養豐富,含有所有的必需氨基酸和牛磺酸、鈣、碘等多種礦物質和維生素,還含有大量生物活性肽[1]。近年來,魚露工業發展迅速,目前國內產量每年約10萬t以上[2]。
魚露加工最主要的問題之一是生物胺含量較高[3]。生物胺是一系列含氮低分子有機堿,可分為單胺和多胺兩類,包括酪胺、組胺、腐胺、尸胺、苯乙胺、色胺、精胺和亞精胺等多種物質。生物胺多存在于蛋白質含量高的食品中,如發酵香腸、魚露、咸肉、咸魚、干酪等[4]。生物胺在人體積累到較高數量時,會產生毒害,如:外部血管膨脹,導致高血壓和頭痛,以及腸部痙攣、腹瀉和嘔吐等。生物胺是造成食品中毒的重要因素之一,引起魚類食品中毒的主要是組胺,引起干酪和發酵香腸中毒主要是組胺和酪胺。而且尸胺和腐胺還可以與亞硝酸鹽反應生成致癌物質亞硝胺[5]。所以,生物胺是影響肉制品特別是發酵肉制品食用安全性的重要因素。
目前,魚露中生物胺的研究主要集中在生物胺含量的檢測[6],魚露發酵過程中生物胺含量變化規律[7],產品加工條件對產品生物胺含量的影響等方面[8]。食品中生物胺主要是由微生物氨基酸脫羧酶催化氨基酸脫羧生成,食品加工條件控制不當和外源性微生物污染是引起生物胺在食品中的積累而達到或超過安全限值的主要原因。如鮮魚制品、魚露制品中的生物胺就是由于外源微生物污染所致[9-10]。
決定生物胺含量的另外一個重要因素是微生物胺氧化酶。胺氧化酶可以降解生物胺,避免生物胺的累積[11]。一般認為,在肉制品發酵初期,肉中游離氨基酸在細菌氨基酸脫羧酶的作用下,分解成生物胺。而隨著發酵的進行,鹽分逐漸滲入肉體,并分布均勻,逐漸升高的鹽濃度和肉制品自身pH值的變化,抑制了生物胺產生菌的生長,而產胺分解酶的細菌卻增加,從而使得生物胺含量降低。隨著產胺分解酶的細菌的生長,兩種細菌達到了一種平衡,胺含量趨于穩定。如發酵魚露中組胺的變化趨勢表明,在魚露前期發酵過程中,組胺含量起初會有所升高,而后下降直至穩定[9]。有關食品中胺氧化酶及其產生菌的研究還很少,僅僅是對微生物進行初步的分離鑒定。如Dapkevicius等[11]從魚露中分離到77株奧默柯達酵母菌,其中48株有胺氧化酶活力,特別是其中2株能消除魚漿中50%以上的組胺。Fadda等[12]從發酵香腸中分離到的干酪乳桿菌CRL705能降解98%的酪胺。Leuschner等[13]發現,64株奧默柯達酵母菌中,27株有較低的胺氧化酶活力;32株亞麻短桿菌(Brevibacterium linens)中,21株有胺氧化酶活力;20株肉葡萄球菌(Staphylococcus carnosus)中未發現有胺氧化酶活力;9株白地霉(Geotrichum candidum)中有1株顯示輕微降解酪胺的活力;44株微球菌屬(Micrococcus sp.)中,17株有胺氧化酶活力。但生物胺降解菌降解生物胺的特性還未見研究報道。
本實驗從自然發酵魚露中篩選分離具有生物胺降解活性的微生物,并研究培養溫度、食鹽質量分數對其降解生物胺的影響,使其能更好的在魚露中運用,提高魚露的食用安全性。同時,對其他生物胺含量高的食品(魚露、發酵香腸、咸肉、咸魚、酸奶、泡菜及葡萄酒等)的加工也有較高的參考價值。
1 材料與方法
1.1 材料、試劑與儀器
自制發酵3個月的魚露(食鹽質量分數40%,料液比1:1,室溫條件下發酵)。
甲醇、組胺、酪胺 美國Sigma公司;苯甲酰氯國藥集團化學試劑有限公司;所有試劑均為分析純。
LC24A高效液相色譜儀 日本島津公司。
1.2 方法
1.2.1 菌種分離及鑒定
分離培養基(MRS):胰蛋白胨10.0g、磷酸氫二鉀2.0g、葡萄糖20.0g、無水醋酸鈉3.0g、牛肉浸膏10.0g、檸檬酸三銨2.0g、酵母浸膏5.0g、七水硫酸鎂0.2g、食鹽10.0g、吐溫-80 1.0mL、L-半胱氨酸0.5g、水1000mL,pH6.8,121℃滅菌15min。
將發酵3個月的魚露以1%接種量接種于50mL改良MRS液體培養基中,30℃培養,發酵2d。采用傾注法獲得微生物單菌落。挑單菌落液體培養,培養2d后接種0.5mL菌液于裝有2.5mL液體MRS培養基的無菌離心管中,同時分別加入0.5mL質量濃度為1mg/mL的組胺和酪胺,發酵3 d,離心,收集上清液,測定生物胺含量,從而確定是否具有生物胺降解活性。固體MRS培養基保存具有生物胺降解活性的微生物用于后續研究。采用26S rDNA測序法對分離到的活性菌進行鑒定。
1.2.2 奧默柯達酵母菌降解生物胺的特性
以分離到的生物胺降解活性最強的奧默柯達酵母菌(M8)為菌種,用上述(MRS)液體培養基,分別在30、35、40℃培養11d,每天測定生物胺的含量變化。
1.2.3 食鹽質量分數對降解魚露中生物胺的影響
以天然自制魚露(食鹽質量分數35%)后作為液體培養基,通過加蒸餾水調整魚露的食鹽質量分數分別為1 5%、2 0%、2 5%,滅菌后,接種M8,接種量1%,在30℃條件下培養11d,每天測定生物胺的含量變化。
1.2.4 HPLC法測定生物胺
采用外標法對樣品中的生物胺進行定量。
1.2.4.1 標準溶液的配制
準確稱取標準品酪胺和組胺各100mg,用0.1mol/L鹽酸定容至100mL,制成質量濃度為1000mg/L混合標準儲備液,置于4℃冰箱保存。吸取0.2、0.5、1.0、1.5、2.5mL上述生物胺標準混合使用液(100mg/L),分別置于10mL容量瓶中,用0.1mol/L HCl稀釋至刻度,混勻,配成最終質量濃度分別為20.0、50.0、100.0、150.0、250.0 mg/L的標準溶液。
1.2.4.2 樣品的前處理及衍生化
用移液搶移取3.0mL發酵液置于5.0mL的離心管中,在5000×g離心30min,吸取1.0mL上清液進行衍生化處理。衍生過程參考Hwang[14]、Qzogul[15]等的方法,并稍作修改。具體方法是:取1.0mL樣品,加入1.0mL 2mol/L NaOH調整pH值至堿性,加入10μL苯甲酰氯水浴30℃衍生處理40min,中間渦旋振蕩4次,每次30s。而后加入2.0mL飽和NaCl于60℃水浴5min終止衍生化,加3.0mL乙醚,旋渦振蕩混合2min,靜置分層,移取上層有機相1.0mL至干凈試管,用氮氣吹干后,加1.0mL甲醇溶解,微濾(0.45μm)后用于測定生物胺含量。
1.2.4.3 色譜條件
色譜條件參考孟甜[16]的基礎上,對梯度洗脫程序和柱溫等進行了適當的改進。Waters symmetry C18色譜柱(4.6mm×250mm,5μm),采用等濃度洗脫,流動相甲醇-水(70:30,V/V);流速:1.0mL/min;檢測波長:254nm;進樣量:20μL;柱溫:30℃。
2 結果與分析
2.1 生物胺標準品和發酵樣品的圖譜
通過分別對組胺和酪胺衍生化圖譜,組胺和酪胺未衍生化圖譜以及生物胺標準溶液的衍生化圖譜的對照,確定組胺和酪胺的出峰位置。同時采用等濃度洗脫,生物胺標準品衍生物圖譜見圖1A。組胺(5.957min)和酪胺(8.190min)分離出峰速度快,10min內所有的生物胺衍生物都被洗脫出來。從生物胺降解菌發酵前后典型圖譜(圖1B、C)可以看出,各生物胺衍生物峰形對稱,且無雜質干擾,因此該流動相等濃度洗脫是可行的。

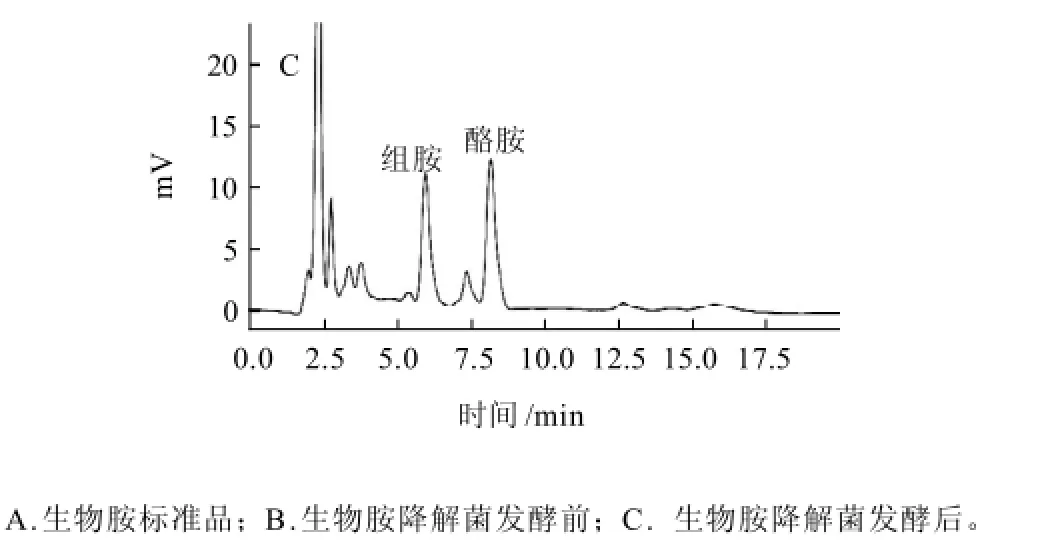
圖1 經生物胺降解菌發酵前后的衍生物色譜圖Fig.1 Chromatograms of biogenic amine standards in MRS medium before and after degradation/fermentation
2.2 HPLC法測定生物胺峰面積與生物胺含量的線性關系
從1.2.4.1節的標準溶液中分別吸取1.0mL混合標準溶液,采用上述方法衍生、測定,每個質量濃度重復測定5次。采用外標法,將兩種生物胺峰面積(y)與相應質量濃度(χ)進行線性回歸,繪制校準曲線,組胺:y=10691χ-49392(R2=0.9965);酪胺:y=15518χ+4979.7(R2=0.9964),表明這2種生物胺標準品在20.0~250.0mg/L質量濃度范圍內線性關系良好(R2>0.995),證明該方法對測定2種生物胺具有良好的可靠性。
2.3 魚露中生物胺降解菌的篩選結果
通過對魚露中奧默柯達酵母菌的分離和純化,獲得46株單一菌種,對其進行單一菌種發酵后,HPLC法測定生物胺降解量。結果分離到10株具有生物胺降解能力的單一菌種,分別編號為M1、M2、M3、M4、M5、M6、M7、M8、M9、M10,將10株具有生物胺降解活性的單一菌株分別接種到含有組胺(18.5mg/L)和酪胺(18.5mg/L)的MRS液體培養基中30℃培養4d,以無菌條件下組胺和酪胺含量均為18.5mg/L的MRS液體培養基作為空白組,然后測定生物胺的含量,結果見表1。
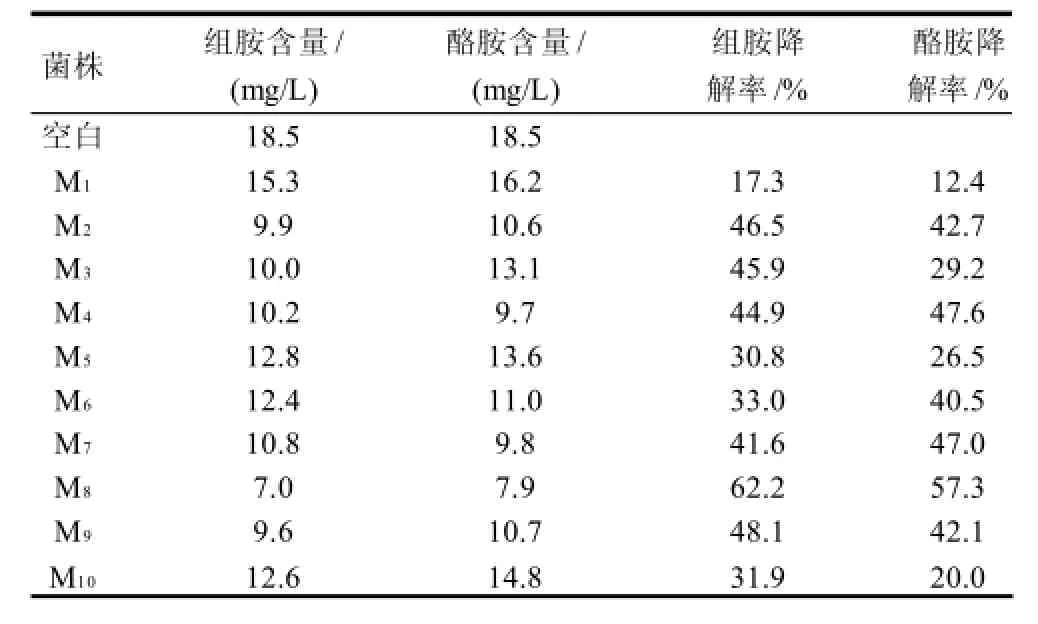
表1 魚露中生物胺降解生物胺菌株篩選結果Table 1 Screening of strains with degradation activity for biogenic amines
由表1可知,組胺含量范圍在7.0~15.3mg/L,酪胺含量范圍在7.9~16.2mg/L。奧默柯達酵母菌對組胺的降解率在17.3%~62.2%之間。奧默柯達酵母菌對酪胺的降解率在12.4%~57.3%之間。其中M8菌株對組胺和酪胺的降解率均為最高,分別達到62.2%和57.3%。
2.4 生物胺降解菌的鑒定
對篩選到降解活性最高的M8菌株進行26S rDNA測序,經Blast在線比對,在線比對結果見表2。

表2 26S rDNA序列Blast比對Table 2 Blast analysis of 26S rDNA sequences
該菌的26S rDNA序列與奧默柯達酵母的26S rDNA序列同源性高達100%,所以初步鑒定該菌為奧默柯達酵母。
2.5 生物胺降解菌在不同溫度和時間中降解生物胺情況分析
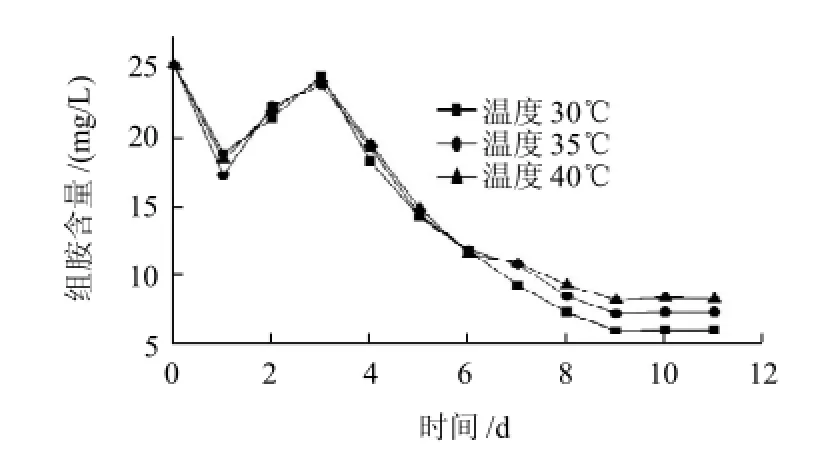
圖2 溫度對M8菌株降解組胺的影響Fig.2 Effect of temperature on the degradation of histamine by strain M8
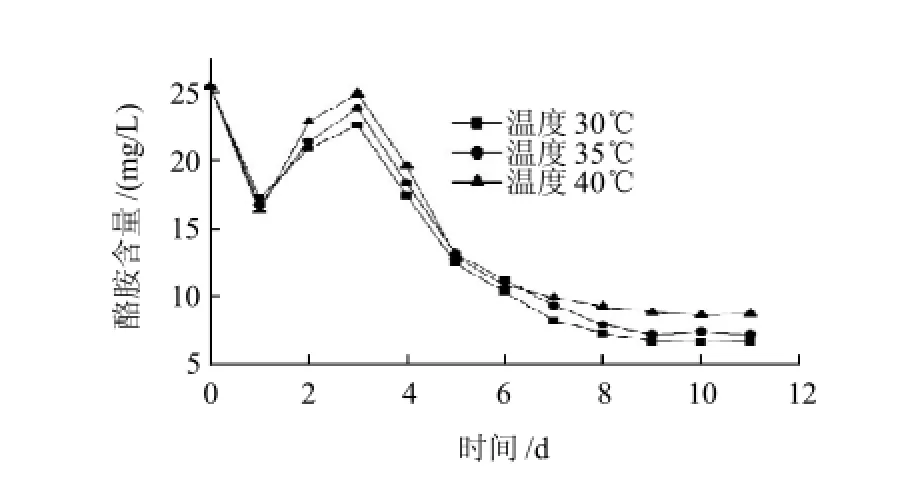
圖3 溫度對M8菌株降解酪胺的影響Fig.3 Effect of temperature on the degradation of tyramine by strain M8
由圖2、3可知,隨著發酵時間的進行,1d時生物胺含量顯著降低,升至3d之后生物胺含量持續降低。在30℃時生物胺的含量最低,其次是35℃,最高的為40℃,在30℃條件下組胺和酪胺的降解率分別達到69.6%和79.2%。但是溫度對生物胺含量變化的影響沒有顯著性。根據生物胺隨發酵時間的變化規律分析,可能在1d時M8菌株生長可能處于適應期,同時其生長可以利用生物胺;2~3d時隨著液體培養基的pH值急劇下降,M8菌株處于對數期,為適應其生長條件的改變,可能會產生生物胺(堿性)以穩定培養液的pH值,所以生物胺含量有所升高;在第4天和第8天M8菌株處于穩定期,菌體數量較大,生物胺降解酶活力較高,生物胺含量迅速降低。第8天開始,M8菌株處于衰退期,對生物胺含量的降解作用變弱直至穩定。
2.6 M8菌株在不同質量分數食鹽魚露中對生物胺降解的情況分析
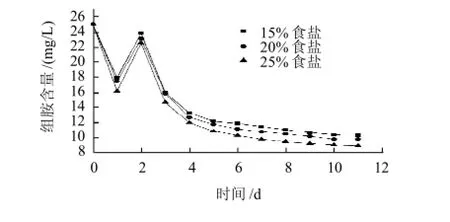
圖4 食鹽質量分數對M8菌株降解組胺的影響Fig.4 Effect of salt concentration on the degradation of histamine by strain M8
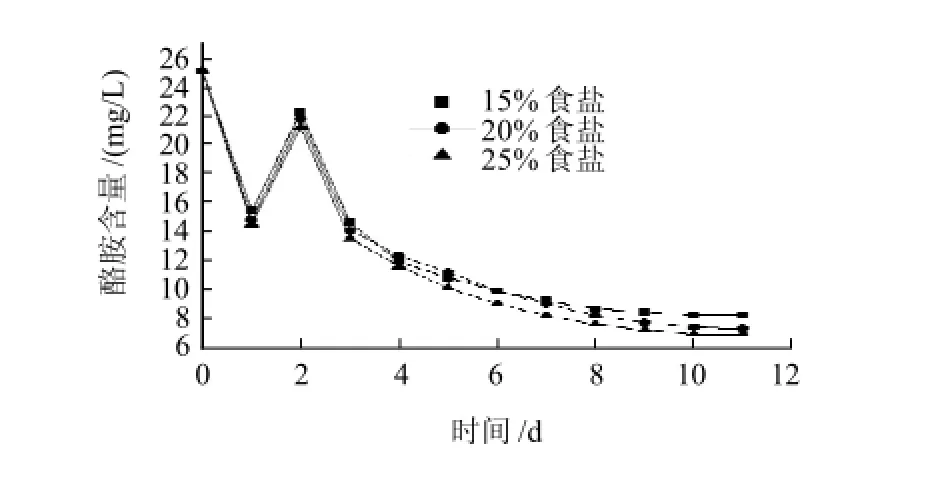
圖5 食鹽質量分數對M8菌株降解酪胺的影響Fig.5 Effect of salt concentration on the degradation of tyramine by strain M8
由圖4、5可知,隨著發酵時間的進行,生物胺含量變化規律為:生物胺含量先降低,然后升高直至穩定。在食鹽質量分數25%條件下生物胺降解效果最好,其次是20%,15%的生物胺降解效果最差。由于M8菌株是從飽和食鹽濃度的魚露中分解得到,M8菌株可能是嗜鹽奧默柯達酵母菌。在食鹽質量分數25%的條件下適宜其生長,在其發酵過程中可能產生了可以氧化降解生物胺的酶或者酶體系,組胺和酪胺的降解率分別達到64.4%和72.4%。
3 結 論
以天然發酵3個月的魚露作為菌種分離材料,研究單一菌株對生物胺降解的影響,篩選到10株具有生物胺降解活性的菌株,證明了魚露中擁有降解生物胺的微生物。其中以M8菌株降解活性最高,對其進行測序,經過在線比對,初步鑒定M8菌株為奧默柯達酵母。
本實驗重點研究了溫度和食鹽質量分數對M8菌株降解生物胺的影響:30℃條件下生物胺的含量最低,其次是35℃,含量最高的為40℃;在30℃條件下,組胺和酪胺的降解率分別達到69.6%和79.2%。25%食鹽質量分數條件下生物胺的含量最低,其次是20%,含量最高的為15%;在食鹽質量分數25%的條件下,組胺和酪胺的降解率分別達到64.4%和72.4%。
篩選到的奧默柯達酵母菌菌能有效降解培養液和魚露中的組胺和酪胺,但篩選到的奧默柯達酵母對魚露品質的影響還需要進一步研究。本研究對其他生物胺含量高的食品(干酪、發酵香腸、咸肉、咸魚、酸奶、泡菜及葡萄酒等)的加工也有較高的參考價值。
[1]陳瑜珠, 陶紅麗, 曾慶孝, 等. 利用羅非魚加工下腳料發酵魚露的研究[J]. 現代食品科技, 2008, 24(5): 441-443.
[2]李陽. 2009年湖南省漁業生產快速增長[EB/OL]. [2011-03-26]. http:// www.yangzhi.com/news/201003/2010_03_26_332221.htmL.
[3]朱志偉, 曾慶孝, 阮征, 等. 魚露及加工技術研究進展[J]. 食品與發酵工業, 2006, 32(5): 96-100.
[4]TSAI Y H, LIN C Y, CHIEN L T. Histamine contents of fermented fish products in Taiwan and isolation of hista-amine forming bacteria[J]. Food Chemistry, 2006, 98: 64-70.
[5]SEN N P, SEAMAN S W. Formation of N-nitroso-N-methylurea in various samples of smoked/dried fish, fish sauce, seafoods, and ethnic fermented/pickled vegetables following incubation with nit rite under acidic conditions[J]. J Agric Food Chem, 2001, 49: 2096-2103.
[6]蔡成崗, 謝銘, 張慧, 等. 高效液相色譜法分析魚制品中的生物胺[J].科技通報, 2010, 26(1): 115-119.
[7]徐偉, 石海英, 朱奇, 等. 魷魚加工廢棄物魚醬油發酵過程中的生物胺變化研究[J]. 安徽農業科學, 2010, 38(4): 2055-2057.
[8]江津津, 黎海彬, 曾慶孝, 等. 鹽度和貯藏溫度對潮汕魚露N-亞硝基化合物的影響[J]. 現代食品科技, 2010, 26(3): 234-237.
[9]孫國勇, 曾慶孝, 江津津. 魚露前期發酵過程中組胺變化規律研究[J]. 中國調味品, 2008(6): 64-67.
[10]KALAC P. Biologically active polyamines in beef, pork and meat products: a review[J]. Meat Science, 2006, 73: 1-11.
[11]DAPKEVICIUS M, ROBERT M J N, ROMBOUTS F M. Biogenic amine formation and degradation by potential fish silage starter microorganisms[J]. International Journal of Food Microbiology, 2000, 57: 107-114.
[12]FADDA S, VIGNOLO G, OLIVER G. Tyramine degradation and tyramine/histamine production by Kodamaea ohmeri yeast and kocuria strains[J]. Biotechnology Letters, 2001, 23: 2015-2019.
[13]LEUSCHNER R G, HEIDEL M, HAMMES W P. Histamine and tyramine degradation by food fermenting microorganisms[J]. International Journal of Food Microbiology, 1998, 39: 1-10.
[14]HWANG D F, CHANG S H, SHIUA C Y, et al. High-performance liquid chromatographic determination of biogenic amines in fish implicated in food poisoning[J]. Journal of Chromatog-Raphy B, 1997, 693 (1): 23-29.
[15]QZOGUL F. Production of biogenic amines by Morganella morganii, Klebsiella pneumoniae and Hafnia alvei using a rapid HPLC method[J]. European Food Research and Technology, 2004, 219(5): 465-469.
[16]孟甜. 乳酸菌菌產生物胺的鑒定及食品中生物胺的檢測[D]. 無錫: 江南大學, 2009.
Screening of Strains with Degradation Activity for Biogenic Amines in Fish Sauce
YANG Li-kun1,2,FU Xiang-jin1,2,*,HU Ye-bi1,2,ZHOU Qi-zhong2,LI Bin1,2
(1. Hunan Provincial Key Laboratory of Deeply Processing and Quality Control of Cereals and Oils, Central South University of Forestry and Technology, Changsha 410004, China;2. College of Food Science and Engineering, Central South University of Forestry and Technology, Changsha 410004, China)
In the present study, strains with degradation activity for biogenic amines were screened from naturally fermented fish sauce, and the effects of fermentation time and temperature on the content of biogenic amines were investigated. A total of 10 strains were obtained from natural fish sauce subjected to fermentation for 3 months. After 72 hours of fermentation at 30 ℃, the degradation rate of histamine was in the range of 17.3%-62.2%, and the degradation rate of tyramine was in the range of 12.4%-57.3%. The strain M8 with the highest activity was identified as Kodamaea ohmeri yeast through 26S rDNA sequence and online sequence alignment. Meanwhile, the strain revealed the strongest degradation activity for biogenic amines at 30 ℃. During fermentation, the content of biogenic amines revealed a decrease on the first day and an increase on the second day as well as the highest level on the third day, and then exhibited a decrease trend until the end of fermentation. After 9 days of fermentation at 30 ℃, the contents of histamine and tyramine were reduced to 30.4% and 20.8% of the initial levels, respectively. The strain M8Kodamaea ohmeri yeast could reveal the best degradation activity for biogenic amines in the presence of 25% salt. Under the optimal conditions, histamine and tyramine were reduced to 35.6% and 27.6%, respectively.
fish sauce;biogenic amines;Kodamaea ohmeri
Q939.97;S986.2
A
1002-6630(2012)11-0158-05
2011-05-10
湖南省自然科學基金項目(11JJ5023)
楊利昆(1986—),男,碩士研究生,研究方向為農產品貯藏與加工。E-mail:yanglikun1986@163.com
*通信作者:付湘晉(1980—),男,講師,博士,研究方向為水產品加工。E-mail:yangtzfu@yahoo.com.cn
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
