半干旱區封育措施下植物生態位研究
2012-07-10 08:49:04殷建
湖南農業科學 2012年11期
殷 建
(新疆生產建設兵團水土保持監測總站,新疆 烏魯木齊 830002)
生態位理論是指種群在時間、空間的位置以及種群在群落的地位和功能作用[1-2]。生態位理論在種間關系、群落結構、生物多樣性及種群進化等研究中的廣泛應用,使其在現代生態學中占有愈來愈重要的地位[3]。研究種群生態位、了解各種群在群落中的地位和作用及種群間的相互關系,對植被資源保護、可持續利用和恢復重建等具有重要意義[4-5]。該研究結合寧夏鹽池荒漠化監測項目,以荒漠化草原人工封育區各植物種為研究對象,研究封育措施下的植物生態位情況,為沙化草地植被恢復提供理論依據,為制定科學合理的草場管理體系提供參考。
1 研究區概況
鹽池縣位于寧夏回族自治區東部,地理坐標北緯 37°04′~38°10′,東經 106°30′~107°41′。鹽池縣北與毛烏素沙漠相連,南靠黃土高原,在地理位置上屬于典型的過渡地帶。這種地理上的過渡性,造就了該縣自然條件資源多樣性和脆弱性的特點。
鹽池縣主要為剝蝕的準平原地形,全縣地勢南高北低,海拔在1 295~1 951 m之間。該縣屬于典型中溫帶大陸性氣候,年均氣溫為8.1℃,極端最高溫度為34.9℃,極端最低溫度為-24.2℃,年均無霜期為165 d,年降水量僅250~350 mm。土壤類型以灰鈣土為主,其次是黑壚土和風沙土,此外有黃土、少量的鹽土和白漿土等。植被類型有灌叢、草原、草甸、沙地植被和荒漠植被。
2 研究方法
2.1 樣地布設及調查方法
2.1.1 樣地布設 研究區域選在鹽池縣的鄂爾多斯緩坡丘陵區內,即毛烏素沙地西南緣的柳林堡鄉。研究設3個處理:老封育區、新封育區和對照區,其中老封育區始于1991年第一批全國防沙治沙試驗示范,采用鐵絲網圍欄,完全排除野生動物和家畜的采食;新封育區自2002年以來采取封育措施;對照區自2002年以來也采取了封育措施,但還是受到一些人為干擾和放牧的影響。3種處理方式在一條直線上,相距不遠,自然條件基本相同。
2.1.2 調查方法 研究于2011年7月進行野外調查。調查方法為從老封育區開始沿著樣帶的方向隨機布設1 m×1 m的樣方,老封育區、新封育區和對照區內各布設10個樣方。調查內容包括:植物名稱、植物種數、株數、蓋度、高度、生物量等。
2.2 數據計算
2.2.1 重要值計測[6]重要值=(相對多度+相對頻度+相對蓋度+相對高度+相對生物量)/5
2.2.2 生態位寬度計測 采用Levins生態位寬度計算公式[7]:
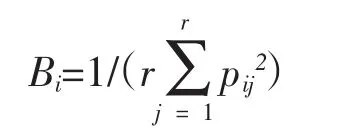
式中:Bi為物種i的生態位寬度;Pij是物種i在第j資源位上的重要值占它所在全部資源位上重要值的比例;r為樣方數。
2.2.3 生態位重疊度計測 采用Pianka生態位重疊指數[8]:
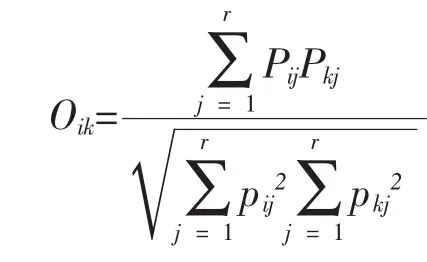
式中:Oik為物種i與物種k的重疊指數,其它符號含義同生態位寬度計算公式。
3 結果與分析
3.1 生態位寬度
從表1中可以看出,老封育區阿爾泰狗哇花、刺沙蓬、隱子草和狗尾草的生態位寬度最大,分別為0.61、0.60、0.52和0.52。這說明,這4種植物對老封育區資源環境的利用能力較強,是老封育區的優勢種。而角蒿、砂珍棘豆和砂藍刺頭的生態位寬度最小,分別為0.15、0.14和0.14,且它們都只在一個樣方中出現過,是老封育區的退化種。
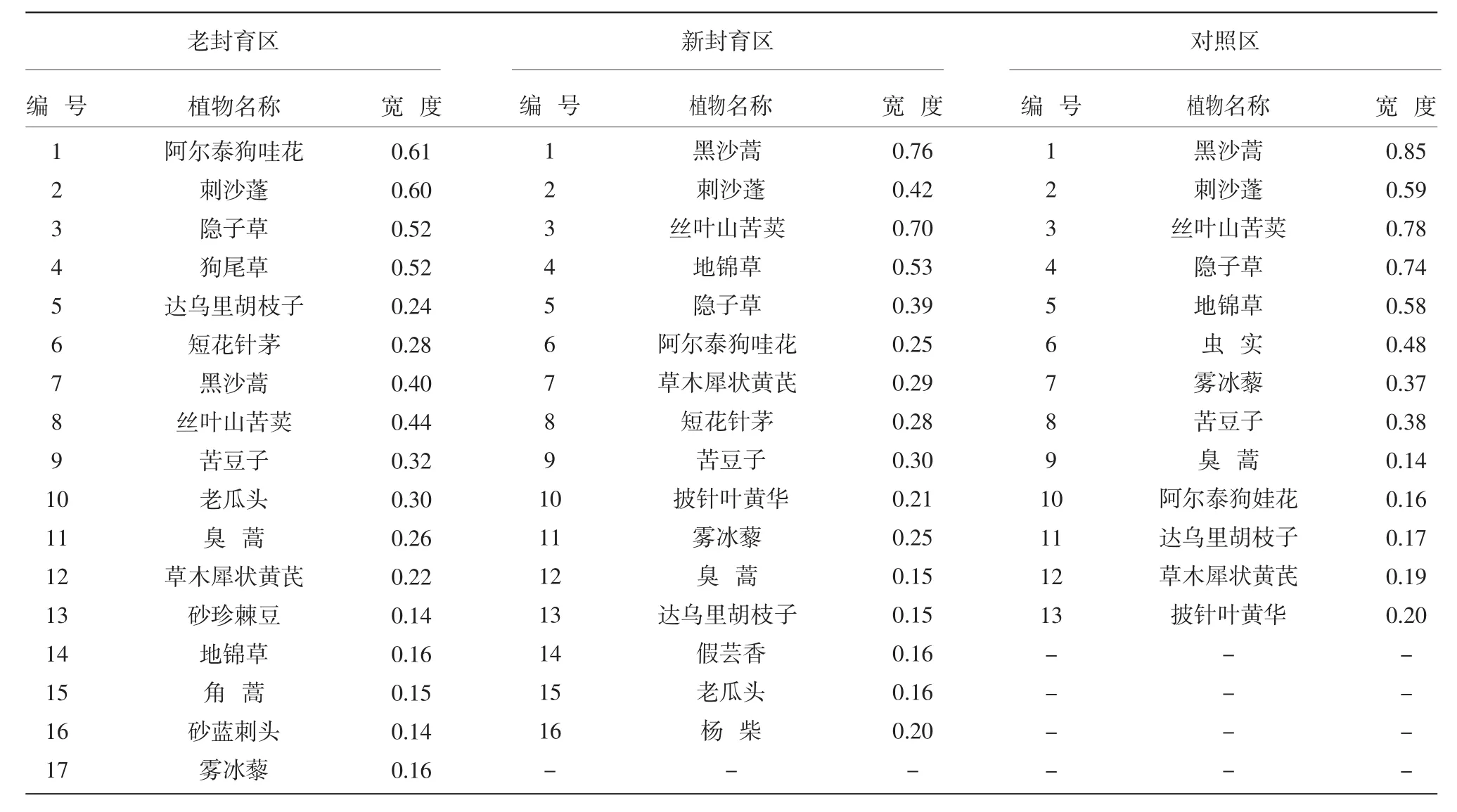
表1 封育區和對照區生態位寬度
新封育區黑沙蒿、絲葉山苦荬、地錦草3種植物的生態位寬度最大,分別為0.76、0.70和0.53。黑沙蒿為菊科植物,在我國北方沙區分布甚廣,從干草原、荒漠草原至草原化荒漠皆有分布。假蕓香、老瓜頭、臭蒿、達烏里胡枝子4種植物的生態位寬度最小,分別為 0.16、0.16、0.15、0.15,它們都只在一個樣方中出現過,說明這幾種植物對新封育區的資源環境利用能力很弱。
對照區黑沙蒿、絲葉山苦荬、隱子草3種植物的生態位寬度最大,分別為0.85、0.78、0.74;達烏里胡枝子、阿爾泰狗娃花、臭蒿3種植物的生態位寬度最小,分別為 0.17、0.16、0.14。
在3個處理中,老封育區共出現17種植物,新封育區共出現16種植物,對照區共出現13種植物。對比3個區域物種的生態位寬度值可以發現,老封育區物種的生態位寬度較其他兩區的相對集中。這說明老封育區的植物分布均勻,生物多樣性較好,群落結構穩定。新封育區植物生態位寬度較對照區的相對集中,說明封育措施對沙化草地植被的恢復有積極作用。對照區由于受人畜的影響較大,對環境適應能力弱的物種的生長受到限制,導致對照區內生物多樣性較差,群落結構單一。
3.2 生態位重疊度
生態位重疊度反映種群之間對資源利用的相似程度和競爭關系。較高的生態位重疊意味著種群之間對環境資源具有相似的生態學要求,因而可能存在著激烈的競爭[9]。由表2可知,老封育區生態位重疊度最大值出現在砂珍棘豆和地錦草、砂珍棘豆和角蒿、地錦草和角蒿、砂珍棘豆和砂藍刺頭、角蒿和砂藍刺頭之間,其重疊度值都為1(事實上只是它們對資源的利用無限接近,而不可能完全一樣,所以重疊值只是接近1而不能達到1)。研究結果表明,生態位寬度較大的阿爾泰狗哇花、刺沙蓬、隱子草和狗尾草與其他物種的生態位重疊度并不高,最高的是阿爾泰狗哇花和隱子草,也僅為0.77。
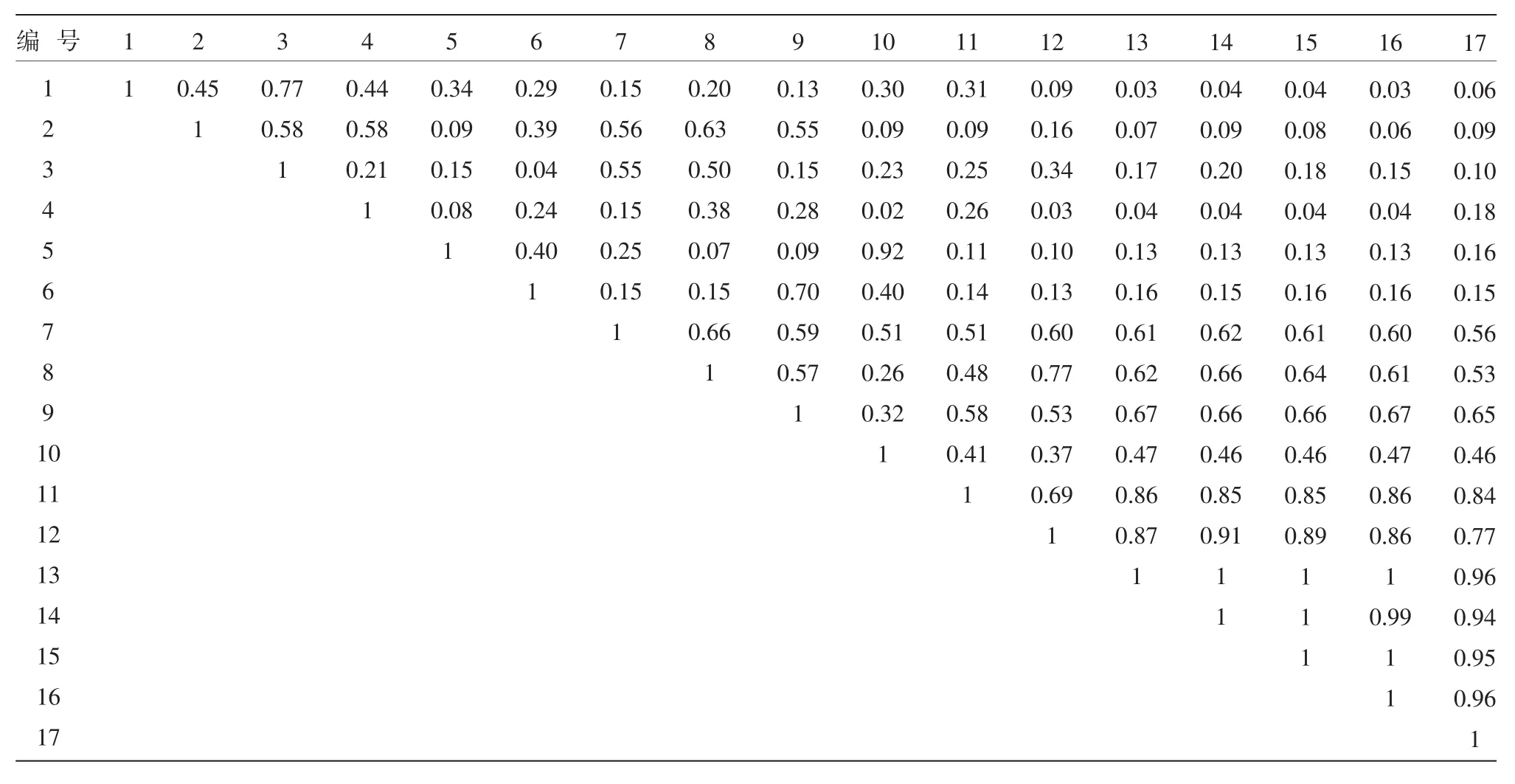
表2 老封育區生態位重疊度
從表3中可以看出,新封育區生態位重疊度最大值出現在達烏里胡枝子和假蕓香、霧冰藜和楊柴、臭蒿和達烏里胡枝子之間,分別為1、0.98和0.93。生態位寬度最大的黑沙蒿、絲葉山苦荬和地錦草與其他物種的重疊度,除了黑沙蒿和絲葉山苦荬為0.81外,均不高。由表4可知,對照區的情況與老封育區和新封育區相似,生態位重疊度最大值均不是出現在生態位寬度最大的幾個物種之間,而是出現在生態位寬度較小的臭蒿和阿爾泰狗娃花之間(0.99)、達烏里胡枝子和草木犀狀黃芪之間(0.98)、臭蒿和達烏里胡枝子之間(0.91)。綜合以上分析可以得出,生態位寬度大的物種不一定和其他物種有大的重疊指數,反之亦然。
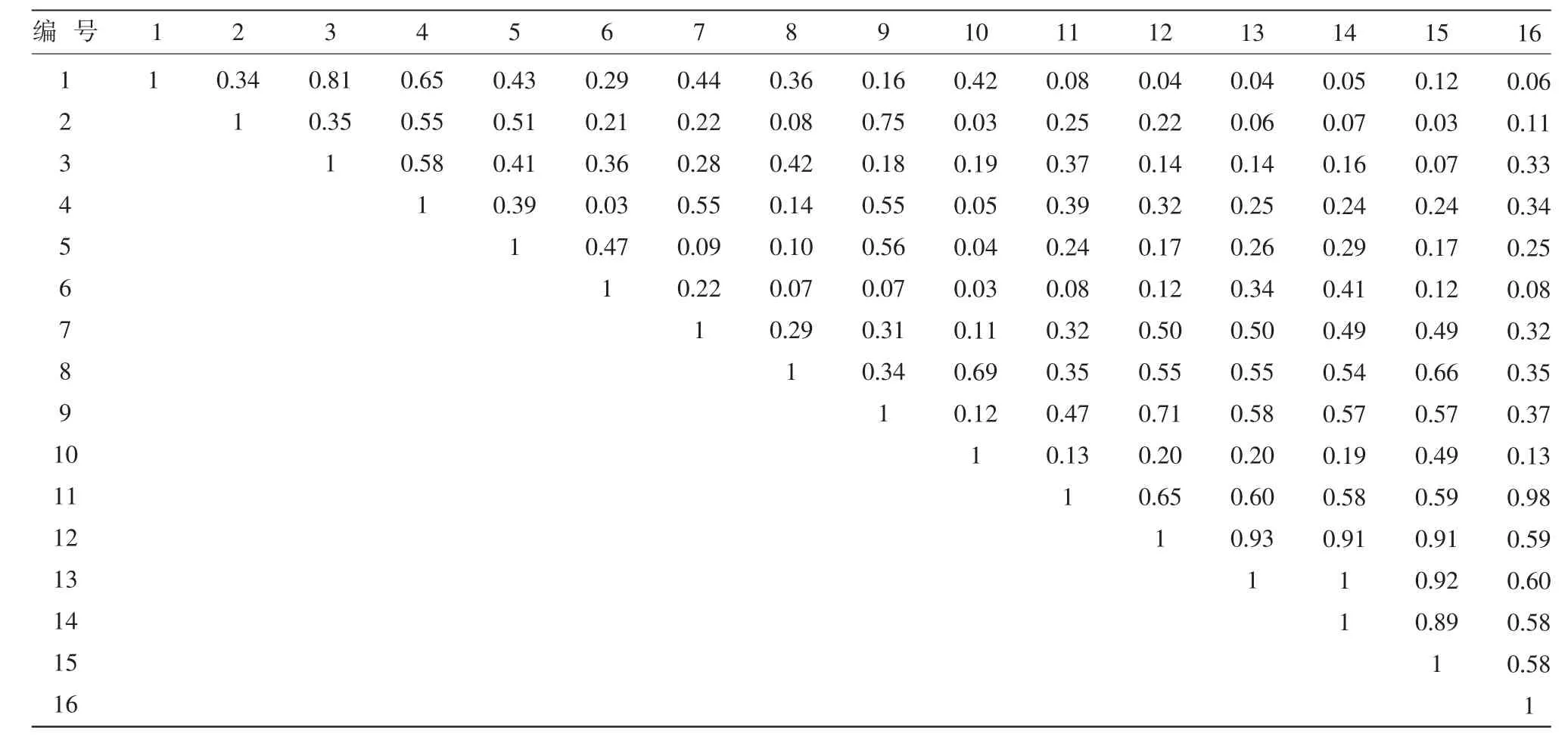
表3 新封育區生態位重疊度
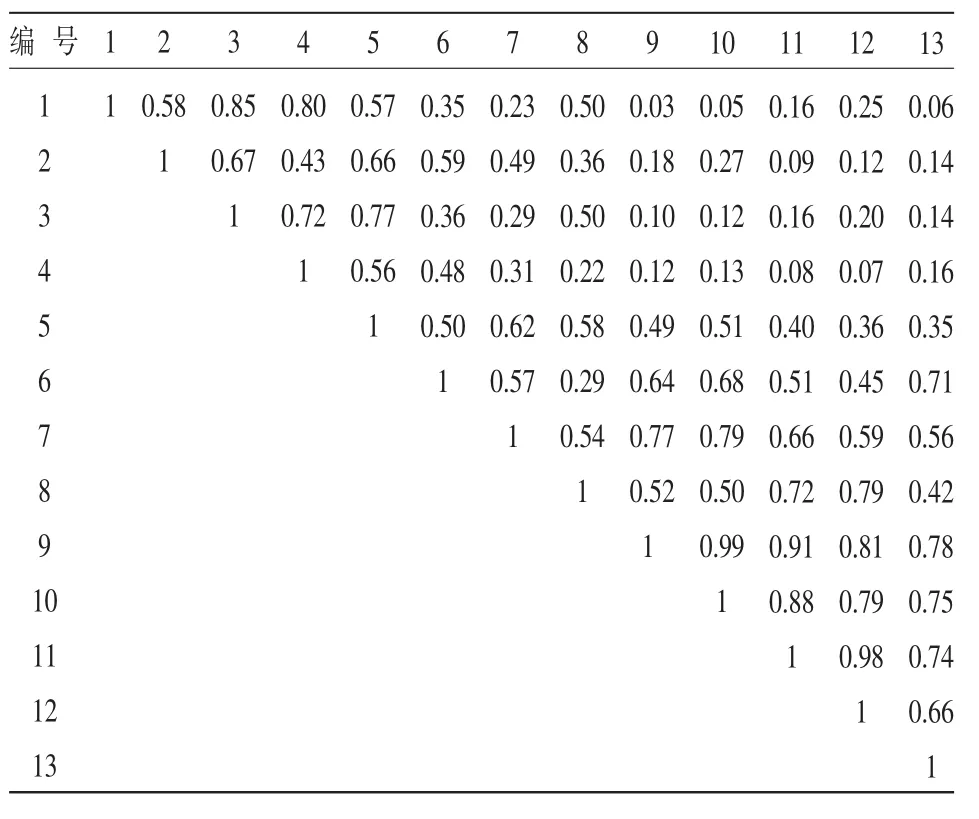
表4 對照區生態位重疊度
4 結論
(1)封育措施對沙化草地植被的恢復具有明顯的促進作用。封育措施避免了人畜對植被的干擾和破壞,給植被提供了較好的生長環境,提高了生物多樣性,增強了群落穩定性。但長期的完全封育狀態不一定有良好的植被恢復效果,要注意適當利用。
(2)生態位寬度是反映物種對環境資源利用狀況的尺度,生態位寬度越大表明物種對環境的適應能力越強,對各種資源的利用越充分,該物種在群落中往往處于優勢地位。老封育區內阿爾泰狗哇花、刺沙蓬、隱子草和狗尾草的生態位寬度最大,是該區的優勢種。新封育區和對照區內生態位寬度最大的植物都是黑沙蒿和絲葉山苦荬,說明它們對該區的生態環境適應能力最強。
(3)生態位重疊度體現了物種對同等級資源的利用程度及空間配置關系,反映物種之間對資源利用的相似程度和競爭關系[10]。多數關于生態位的研究認為,較大的生態位寬度常常伴隨著較高的生態位重疊度[11-13]。而筆者的研究結果表明,較高的生態位寬度和較高的生態位重疊度之間并不存在直接的線性關系。
[1]Grinnell J.The niche-relationship of the California thrasher[J].Auk,1917,34(10):428-431.
[2]Elton C.Animal ecology[M].London:Sidgewick and Jackson,1927.63-68.
[3]王 剛,趙松嶺,張鵬云,等.關于生態位定義的探討及生態位重疊計測公式改進的研究[J].生態學報,1984,4(2):119-127.
[4]張國斌,李秀芹.嶺南自然保護區常綠闊葉林優勢樹種的生態位研究[J].南京林業大學學報(自然科學版),2007,31(4):46-50.
[5]哀建國,吳谷漢,陳錦宇,等.石垟林場省級森林公園常綠闊葉林種群生態位特征 [J].浙江大學學報(農業與生命科學版),2006,32(3):576-584.
[6]鄭翠玲,曹子龍,趙廷寧,等.渾善達克沙地南緣農牧交錯帶棄耕地植被的演替規律[J].中國水土保持學,2005,3(1):72-76.
[7]魏志琴,李旭光,郝云慶.珍稀瀕危植物群落主要種群生態位特征研究[J].西南農業大學學報(自然科學版),2004,26(1):1-4.
[8]畢潤成,尹文兵,王艷妮.山西南部脫皮榆種群生態位的研究[J].西北植物學報,2003,23(7):1266-1271.
[9]Silvertown J W.Plants in limestone pavements:tests of species interaction and niche separation[J].J Ecol,1983,71:819-828.
[10]Turner R M.Long-term vegetation change at a fully protected Sonoran desert site[J].Ecology,1990,(7):464-477.
[11]Walker B.Conserving biological diversity through ecosystem resilience[J].Conser Biol,1995(9):747-752.
[12]陳 波,周興民.三種蒿草群落中若干植物種的生態位寬度與重疊分析[J].植物生態學報,1995,19(2):158-169.
[13]郭全邦,劉玉成,李旭光.縉云山森林次生演替序列優勢種群生態位[J].西南師范大學學報(自然科學版),1997,22(1):73-78.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
