IFN-γ誘導胰腺癌細胞IDO表達及活性的變化
2012-07-21 09:12:22龔勇
山西衛生健康職業學院學報 2012年6期
龔 勇
(長治市衛生學校,山西 長治 046000)
在機體免疫系統與腫瘤細胞相互對抗的過程中,被激活的T細胞與NK細胞分泌的IFN-γ具有直接抑制腫瘤細胞增殖、抑制腫瘤血管生成、調控表面腫瘤細胞的Fas/FasL表達等多種抗腫瘤作用。作為一種重要的雙向免疫調節因子,IFN-γ與免疫的激活和抑制都著密切的聯系。IFN-γ不僅能上調機體內樹突狀細胞的吲哚胺2,3-雙加氧酶(IDO)表達[1],而且能夠誘導許多腫瘤細胞的IDO表達,如骨肉瘤、上皮惡性腫瘤細胞和肺纖維細胞等[2]。而IDO在腫瘤免疫逃逸中起著極其重要的作用[3]。本研究旨在觀察IFN-γ對胰腺癌細胞株Panc02中IDO表達及活性的影響,為深入了解胰腺癌免疫逃逸機制提供實驗基礎。
1 材料與方法
1.1 材料
RPMll640培養基、胎牛血清購自美國Hyclone公司,重組鼠γ干擾素(IFN-γ)購自美國Pepro Tech公司,BCA試劑盒、PVDF Transfer Membranes購自美國Pierce公司,Bradford蛋白定量試劑盒購自南京建成生物工程研究所,辣根酶標記山羊抗兔IgG購自北京中杉金橋公司,Super ECL Plus超敏發光液購自普利萊基因技術有限公司,預染低分子量蛋白質Marker購自Solarbio公司,Actin(1-19)購自 SANTA CRUE,氨基酸色柱購自天津四友公司,犬尿氨酸標準品、內標3-硝基酪氨酸標準品購自中國藥品生物制品檢定所,胰腺癌細胞株Panc02購自美國ATCC公司。
1.2 實驗分組及方法
按每個培養瓶為一個處理對象,隨機分為4個處理組:空白對照組(IFN-γ 0U/mL)和 IFN-γ 10 U/mL、20 U/mL、50 U/mL組。待細胞生長至60%匯合時,IFN-γ處理組分別加入終濃度為10 U/mL、20 U/mL、50 U/mL的IFN-γ,空白對照組加入等量PBS,繼續培養18 h,收集半貼壁細胞,并抽提細胞胞漿蛋白進行Western Blot分析以觀察各組IDO的蛋白表達。每組樣本例數均為6例。
1.3 Western blot檢測 IFN-γ作用胰腺癌細胞株Panc02后IDO蛋白的表達
采用胞質胞核蛋白抽提試劑盒,操作步驟嚴格按照試劑盒說明書進行。提取的蛋白樣品經變性、電泳、轉膜及封閉后,分別與小鼠抗大鼠單克隆抗體和辣根過氧化物酶標記的二抗孵育。暗室中采用ECL方法對反應產物進行檢測,當條帶達所需亮度時立即顯影定影。采用Quantity one圖像分析系統進行各條帶平均灰度值(A)分析,以目的條帶的A值與內參照β-actin A值的比值表示IDO的相對表達量。
1.4 高壓液相測定培養液上清中犬尿氨酸(Kyn)的濃度以評價IDO的分子活性
取4個處理組的培養液上清,利用高壓液相測定其中Kyn的濃度來評價IDO的分子活性。色譜條件:色譜柱為waters RP-C18柱,流動相為15 mmol/L乙酸-乙 酸 鈉 溶 液 (0.2 mmol/L, pH3.6),流 速 為1.0 mL/min,紫外檢測波長為225 nm,內標為 3-硝基酪氨酸(3-n)。
1.5 統計學方法
數據均以均數±標準差表示,應用SPSS11.5統計軟件進行單因素方差分析,組間差異用SNK-q檢驗,兩兩比較采用LSD的方法,以P<0.05具有統計學意義。
2 結果
2.1 IFN-γ誘導胰腺癌細胞株Panc02中IDO的表達增強
IDO、β-actin特異性免疫印跡條帶的相對分子質量分別為42 KD和43 KD。不同濃度IFN-γ誘導Panc02的IDO蛋白表達量見表1及圖1。經過分析可知,4組的IDO/β-actin值之間差異具有統計學意義(F=21.416,P <0.05),可以認為 4 組 IDO 蛋白的表達量存在差別,進一步采用LSD的方法進行兩兩比較,比較結果顯示,除10 U/mL組與20 U/mL組差別無統計學意義(P=0.238)之外,其余各組間均存在差異(P均小于0.05)。與空白對照組比較,IFN-γ作用后胰腺癌細胞株Panc02的IDO蛋白表達量明顯增加,且隨著IFN-γ作用濃度的增大其IDO的表達量逐漸增加,以50 U/mL組為著。
表1 4 組 IDO/β-actin 值比較(±s)Table 1 Comparison of IDO/β-actin Value in Four Groups(ˉx ± s)
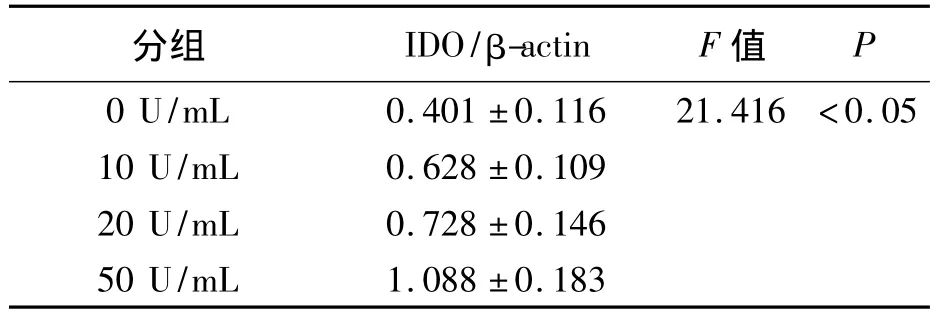
表1 4 組 IDO/β-actin 值比較(±s)Table 1 Comparison of IDO/β-actin Value in Four Groups(ˉx ± s)
分組 IDO/β-actin F 值P 0 U/mL 0.401 ±0.116 21.416 <0.05 10 U/mL 0.628 ±0.109 20 U/mL 0.728 ±0.146 50 U/mL 1.088 ±0.183
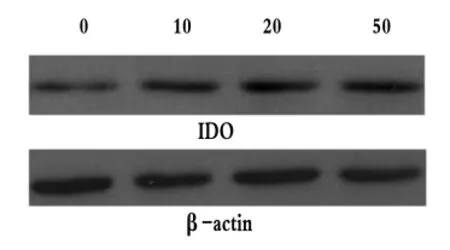
圖1 不同濃度IFN-γ(U/mL)誘導β-actin和IDO蛋白表達量Fig 1 IDO Protein Expression Levels of β-actin Induced by Different Concentrations of IFN-γ(U/mL)
2.2 IFN-γ誘導胰腺癌細胞株Panc02中IDO的活性增強
不同濃度IFN-γ刺激Panc02后細胞培養液上清中Kyn的濃度見表2所示。經過分析可知,4組之間差別具有統計學意義(F=15.404,P <0.05),可以認為4組Kyn濃度存在差別,進一步采用LSD的方法進行兩兩比較,比較結果顯示,除0 U/mL組與10 U/mL組差別無統計學意義(P=0.525)之外,其余各組間都存在差別(均P<0.05)。結果顯示:與空白對照組比較,IFN-γ作用后胰腺癌細胞株Panc02的細胞培養液上清中Kyn的濃度明顯增加,且隨著IFN-γ作用濃度的增大其Kyn的濃度逐漸增加,以50 U/mL組為著。

表2 不同濃度IFN-γ刺激Panc02后細胞培養液上清中Kyn的濃度Table 1 Kyn Concentrations in the Supernatant of Panc02 Cell Culture Media with Different IFN-γ Concentrations
3 討論
IDO是細胞內含亞鐵血紅素的酶,是肝臟外唯一可以沿犬尿酸途徑催化色氨酸分子中吲哚環氧化裂解并進行分解代謝的第一限速酶。作為一種免疫調節酶,IDO有可能為對母嬰耐受的理解、自身免疫疾病的調控以及器官移植排斥反應的抑制等多個領域起重要作用。
近年來大量文獻報道表明,IDO在腫瘤發生發展過程中以及腫瘤免疫逃逸中起著重要作用[4]。研究結果顯示,人類多種腫瘤細胞均高表達IDO[4]。在動物模型中,轉染IDO的腫瘤細胞株可以有效地對抗機體T細胞的保護,在宿主內浸潤性生長[3]。臨床資料也顯示,在卵巢癌和結腸癌的患者中,IDO的表達可以作為一個不良預后的顯著標志[4]。
“色氨酸饑餓”、“代謝產物的毒性作用”以及兩者之間的聯合作用作為解釋IDO在腫瘤免疫逃逸中的作用機制已經被人們廣泛接受。
有研究結果表明,由于對“色氨酸饑餓”的敏感性,處于G1期中期的T細胞缺乏色氨酸的培養環境中,增殖被抑制。而且,這種被抑制的T細胞被再次激活是非常困難的[5]。同時有類似報道,如果培養條件缺乏色氨酸下,小鼠初始 T細胞活化后可以表達CD69和CD25,能夠經歷細胞周期 G0/G1期的前12 h,但不表達細胞周期素依賴性蛋白激酶4、周期素D3以及CD71,而且無cDNA合成,因此不能成功分化為效應T細胞。IDO有可能通過色氨酸饑餓誘導抗原特異性的T細胞耐受,其作用機制與細胞周期被阻斷以誘導T細胞凋亡有關[6]。
有研究結果提示,犬尿氨酸在抗原特異性CD4+T細胞分化中有著重要作用,而哇琳酸、吡啶羥酸、L-犬尿氨酸等也可抑制NK細胞的增殖[7]。色氨酸饑餓和代謝產物毒性作用的聯合作用可以在T細胞水平上下調CD8細胞的TCRζ-鏈并誘導調節性CD4細胞的產生[8,9]。IDO基因的轉錄起始位點的上游包含許多調控元件。其啟動子長1245 bp,含有3個IFN-γ激活序列以及2個干擾素調節因子。它們是IFN-γ誘導IDO表達信號通路中必需的環節。IFN等細胞因子可以通過5'端的調節區域對IDO基因表達進行調控。其中IFN-γ的刺激潛能最強,體外研究表明其激活潛能具有時間、劑量依賴性和種屬特異性。IFN-γ不僅能上調機體內樹突狀細胞的IDO表達,而且能夠誘導許多腫瘤細胞的IDO表達,如骨肉瘤、上皮惡性腫瘤細胞和肺纖維細胞細胞等。IFN-γ可誘導鼻咽癌細胞株中IDO的表達,并抑制患者細胞免疫反應[10]。
本研究結果表明,IFN-γ作用后胰腺癌細胞株Panc02的IDO蛋白表達量明顯增加,且隨著IFN-γ作用濃度的增大其IDO的表達量逐漸增加,以50 U/mL組為著。并且Panc02細胞株中IDO的活性在IFN-γ的刺激下顯著增強。因此以下假設是合理的:胰腺癌細胞表達的IDO可以通過色氨酸饑餓及代謝產物的毒性作用等途徑,改變腫瘤局部微環境,使T細胞分化或/和轉化具有抑制表型的Treg。這些細胞分泌諸如:TGF-β,Ⅰ、Ⅱ型干擾素等可溶性細胞因子,募集更多具有抑制或者調節表型的細胞,同時通過反向信號轉導通路刺激IDO表達進一步提高,并誘導CD+8DC向免疫耐受型轉化,提高IDO+DC的數量,從而形成TGF-β-IDO-Treg正反饋環,使得免疫抑制從腫瘤周圍播散開來。因此認為,IFN-γ誘導胰腺癌細胞IDO表達上調且活性增強,這可能是引起胰腺癌免疫逃逸的重要機制之一。
[1]徐 鈞,孔祥順,李元棟.不同濃度γ干擾素誘導大鼠脾臟來源樹突狀細胞表達吲哚胺2,3-雙加氧酶的變化及對T淋巴細胞增殖的影響[J].中華實驗外科雜志,2011,28(10):1686-1689.
[2]Munn,D.H,A.L.Mellor.Indoleamine 2,3-dioxygenase and Tumorinduced Tolerance[J].J Clin Invest,2007,117(5):1147-1154.
[3]Uyttenhove,C,Pilott L,Theate I,et al.Evidence for a Tumoral Immune Resistance Mechanism Based on Typtophan Dgradation by Idoleamine 2,3-dioxygenase[J].Nat Med,2003,9(10):1269-1274.
[4]Brandacher G,Perathoner A,Ladurner R,et al.Prognostic Value of Indoleamine 2,3-dioxygenase Expression in Clorectal Cancer:Effect on Tumor-infiltrating T Cells[J].Clin Cancer Res,2006,12(4):1144-1151.
[5]Bauer T.M,Jiga LP,Chuang JJ,et al.Studying the Immunosuppressive Role of Indoleamine 2,3-dioxygenase:Tryptophan Metabolites Suppress Rat Allogeneic T-cell Responses in Vitro and in Vivo[J].Transpl Int,2005,18(1):95-100.
[6]Lee G.K,Park HJ,Macleod M,et al.Tryptophan Deprivation Sensi-tizes Activated T Cells to Apoptosis Prior to Cell Division[J].Immunology,2002,107(4):452-460.
[7]Frumento G,Rotondo R,Tonetti M,et al.Tryptophan-derived Catabolites are Responsible for Inhibition of T and Natural Killer Cell Proliferation Induced by Indoleamine 2,3-Dioxygenase[J].J Exp Med,2002,196(4):459-468.
[8]Fallarino F,Grohmann U,Vacca C,et al.T cell Apoptosis by Tryptophan Catabolism[J].Cell Death & Differ,2002,9(10):1069-1077.
[9]Fallarino F.The Combined Effects of Tryptophan Starvation and Tryptophan Catabolites Down-regulate T Cell Receptor Zeta-chain and Induce a Regulatory Phenotype in Naive T Cells[J].J Immunol,2006,176(11):6752-6761.
[10]Liu P,Xie Bailu,Cai Shaohui,et al.Expression of Indoleamine 2,3-dioxygenase in Nasopharyngeal Carcinoma Impairs the Cytolytic Function of Peripheral Blood Lymphocytes[J].BMC Cancer,2009,9:416.
