Smad4對(duì)小鼠眼瞼發(fā)育的影響
2012-07-31 14:06:26顧欣祖LUMichaelDINGChuanqing
中國(guó)病理生理雜志 2012年2期
劉 瑛,顧欣祖,LU Michael,DING Chuanqing
(1中山大學(xué)中山眼科中心,國(guó)家重點(diǎn)眼科學(xué)實(shí)驗(yàn)室,廣東 廣州 510060;2Department of Ophthalmology and Doheny Eye Institute;3Department of Cell and Neurobiology,Keck School of Medicine,University of Southern California,Los Angeles,CA 90033,USA)
哺乳類動(dòng)物其眼瞼的發(fā)育源自表皮外胚層及其下的眼周中胚層,包括眼瞼特質(zhì)化、生長(zhǎng)、閉合及重開(kāi)4個(gè)連續(xù)的過(guò)程。小鼠眼瞼的特質(zhì)化始于E9.0,此時(shí)轉(zhuǎn)錄因子Foxl2表達(dá),確定眼瞼相對(duì)于眼球的位置[1]。在E11.5,背側(cè)及腹側(cè)的眼周外胚層內(nèi)陷,形成所謂的眼瞼褶(eyelid fold),此后到E15.5,兩側(cè)的眼瞼褶開(kāi)始相向生長(zhǎng),形成上下瞼。在E15.5左右,上下瞼緣的外表皮層開(kāi)始向角膜方向延伸,最終于E16.5左右匯合并融合。上下瞼在生后12 d左右開(kāi)始分離,大約2 d 后眼瞼重新睜開(kāi)[2-3]。
眼瞼的閉合和重開(kāi)是所有哺乳類動(dòng)物的必經(jīng)之路,眼瞼的正常閉合與開(kāi)放對(duì)于角結(jié)膜的發(fā)育及維持眼表的健康至關(guān)重要,當(dāng)眼瞼的生長(zhǎng)或融合發(fā)生缺陷時(shí),便可出現(xiàn)眼瞼在出生時(shí)保持開(kāi)放狀態(tài)(eye open at birth,EOB),眼瞼的閉合涉及到很多基因和信號(hào)通路的調(diào)控,這些因素的破壞或改變可能都會(huì)引起EOB的發(fā)生[4]。我們通過(guò)對(duì)Smad4條件性基因敲除小鼠的研究發(fā)現(xiàn),該基因敲除鼠除了具有小眼球的表型之外也出現(xiàn)了EOB的表型,因此我們進(jìn)行了更進(jìn)一步的研究,試圖更深入探討Smad4對(duì)眼瞼閉合的影響及其可能的調(diào)控機(jī)制。我們所采用的Smad4條件性基因敲除鼠借助Cre/LoxP系統(tǒng),通過(guò)Pax6引導(dǎo)的Cre轉(zhuǎn)基因表達(dá)產(chǎn)生的Cre重組酶介導(dǎo)靶基因Smad4兩側(cè)的LoxP間發(fā)生切除反應(yīng),從而獲得特定于眼部某些組織包括眼瞼的Smad4的敲除,以便觀察Smad4對(duì)于某些特定組織器官發(fā)育的影響。
材料和方法
1 動(dòng)物
所用動(dòng)物遵循眼及視覺(jué)科學(xué)研究動(dòng)物使用聲明。Smad4fl/fl小鼠由美國(guó)南加州大學(xué)Dr.Xun Xu實(shí)驗(yàn)室提供,Pax6啟動(dòng)子控制下的Le-Cre轉(zhuǎn)基因小鼠來(lái)自美國(guó)華盛頓大學(xué)Dr.David Beebe實(shí)驗(yàn)室,ROSA26報(bào)告基因小鼠R26R品系購(gòu)自Jackson。Le-Cre的小鼠與 Smad4fl/fl小鼠交配產(chǎn)生 Le-Cre/Smad4fl/+小鼠,Le-Cre;Smad4fl/+小鼠繼而再與Smad4fl/fl小鼠交配產(chǎn)生 Le-Cre/Smad4fl/fl小鼠,即我們所需要的條件性Smad4基因敲除小鼠——Smad4 CKO小鼠。Smad4fl/fl小鼠作為對(duì)照,即Smad4野生鼠(Smad4 WT)。胎齡以陰部栓子的檢查為基礎(chǔ)確定,該日中午12點(diǎn)為孕0.5,記為E0.5。
2 方法
2.1 組織學(xué)檢查 于解剖顯微鏡下獲取適當(dāng)胎齡的小鼠胚胎觀察其眼瞼變化并拍照,隨后于4%多聚甲醛中4℃固定過(guò)夜。梯度乙醇脫水,石蠟包埋,切片7.5 μm厚,常規(guī)HE染色觀察形態(tài)學(xué)。
2.2 免疫組化 石蠟切片常規(guī)脫蠟至水,10 mmol/L枸櫞酸鈉緩沖溶液(pH6.0)微波抗原修復(fù),3%甲醇-過(guò)氧化氫溶液室溫下孵育10 min阻斷內(nèi)源性過(guò)氧化物酶,5%BSA/0.1%Triton X-100室溫阻斷30 min,抗小鼠 I抗[Smad4、phalloidin(Invitrogen);表皮生長(zhǎng)因子受體(epidermal growth factor receptor,EGFR,1∶100),c - Jun(1∶100)、p - c - Jun(1∶100,Cell Signaling)]4℃過(guò)夜孵育;再用熒光Ⅱ抗(Jackson Immunoresearch)或者 Histostain SP Broad Spectrum(HRP)試劑盒(Invitrogen)并以DAB(Zymed)顯色。
2.3 肌動(dòng)蛋白的檢測(cè) 鋪片及切片染色采用E15.5眼瞼組織或切片,與熒光I抗Alexa Fluor 568 phalloidin(Invitrogen)4℃過(guò)夜孵育,封片后鏡檢。
2.4 BrdU標(biāo)記 (1)配制 BrdU溶液10 g/L,取特定孕期小鼠,稱重,按100 μg BrdU/g BW腹腔注射,1 h后解剖顯微鏡下取胚胎,Carnoy(30%氯仿,10%冰醋酸,60%甲醛)固定液固定過(guò)夜;(2)切片經(jīng)二甲苯與梯度乙醇脫蠟至水,PBST浸洗;(3)3%過(guò)氧化氫室溫孵育10 min,PBST浸洗3次,每次5 min;(4)20 mg/L的蛋白酶K于37℃消化18~20 min,PBST浸洗3次,每次5 min;(5)1 mol/L HCl中37℃變性45 min,PBST浸洗3次,每次5 min;(6)Cas block阻斷液孵育30 min,PBST浸洗3次,每次5 min;(7)抗BrdU I抗室溫孵育1 h,PBST洗3次,每次5 min;(8)余步驟同常規(guī)免疫組化。
2.5 TUNEL染色 (1)石蠟切片脫蠟至水;(2)蛋白酶K室溫下孵育10 min;PBST洗2次每次5 min;(3)取100 μL標(biāo)記液用于陰性對(duì)照,按1∶9取標(biāo)記液和酶液混勻成TUNEL反應(yīng)液,切片以反應(yīng)液37℃水浴溫箱避光孵育1 h;PBST洗3次,每次5 min;(4)DAPI工作液孵育3~5 min,PBST洗3次,每次5 min;(5)透明封片劑封片,熒光顯微鏡下觀察并照相。
結(jié) 果
1 Le-Cre條件性Smad4基因敲除鼠眼瞼上皮的延伸受損
在E14.5,兩者都表現(xiàn)為同樣的眼瞼開(kāi)放及外形,無(wú)明顯的形態(tài)學(xué)差別,見(jiàn)圖1 A1、B1。從E15.5后,兩者眼瞼表型出現(xiàn)顯著差別。在野生型小鼠胚,上下瞼緣出現(xiàn)突起,并從突起處開(kāi)始向相對(duì)方向延伸移行,并最終融合,見(jiàn)圖 1 C1、E1、G1、G2。相比之下,變異型小鼠自始至終其瞼緣上皮未見(jiàn)延伸,上下瞼緣也一直未見(jiàn)閉合,眼表一直處于暴露狀態(tài),見(jiàn)圖1 D1、F1、H1、J1。該結(jié)果也暗示變異小鼠的 EOB表型是因?yàn)檠鄄€未能閉合造成,而不是過(guò)早的眼瞼開(kāi)放。
在E14.5左右的野生鼠胚,眼瞼上皮僅由1層基底細(xì)胞構(gòu)成,見(jiàn)圖1 A2,在E15.5左右因?yàn)榛准?xì)胞的增殖和分化而形成多層結(jié)構(gòu),此后上下眼瞼的上皮層自其尖端相對(duì)移行,兩上皮層在眼表中央相遇而最后融合在一起,見(jiàn)圖1 C2、E2、G2、I2。E14.5的變異鼠胚眼瞼的形態(tài)學(xué)與同齡野生鼠胚相比并無(wú)顯著差異,見(jiàn)圖1 B2,而且在E15.5其眼瞼上皮也增厚而形成多層結(jié)構(gòu),但其眼瞼上皮層一直未出現(xiàn)延伸,直到出生,其眼瞼始終處于開(kāi)放狀態(tài),見(jiàn)圖1 D2、F2、H2、J2。

Figure 1.Histological and morphological changes of embryonic eyelids in Smad4 CKO mice(HE,×200).A1,A2:in wild-type embryos at E14.5,upper and lower eyelids kept seperated,and the eyelid epithelium comprised a single layer of basal cells;B1,B2:mutant eyelids at E14.5 appeared similar to wild-type eyelids;C1,C2:at E15.5,eyelid epithelium in wild-type embryos became multilayered and the eyelid epithelial sheet started to extend from both top and bottom eyelid tips;D1,D2:at E15.5,eyelid epithelium in mutant embryos became multilayered but epithelial sheet didn't extend;E1,E2:at E16.0,the eyelid epithelial sheet in wild-type embryos continued to extend,meet and cover the ocular surface;F1,F(xiàn)2:at E16.0,no extension of epithelial sheet was observed;G1,G2:at E16.5,top and bottom eyelids almost fused;H1,H2:no migration of eyelid epithelium was observed;I1,I2:eyelid closure in wild - type newborn;J1,J2:eyelids remained open in mutant newborn.圖1 變異小鼠胚胎眼瞼的組織形態(tài)學(xué)改變
2 Le-Cre條件性Smad4基因敲除鼠眼瞼上皮細(xì)胞內(nèi)肌動(dòng)蛋白束形成受損
結(jié)果顯示在野生型眼瞼上皮中觀察到明顯的F肌動(dòng)蛋白束,位于眼瞼上皮層的進(jìn)行緣,呈環(huán)形包繞眼瞼邊界,見(jiàn)圖2 A、B。相反,在變異型其眼瞼上皮前緣未能檢測(cè)到此種F肌動(dòng)蛋白纖維,見(jiàn)圖2 C、D。眼瞼冰凍切片的phalloidin染色也顯示,野生型在其眼瞼上皮層的各層細(xì)胞內(nèi)均有顯著的F肌動(dòng)蛋白染色,見(jiàn)圖2 E,而在變異型中很少有該肌動(dòng)蛋白的裝配,見(jiàn)圖2 F。這些結(jié)果顯示在Smad4基因敲除鼠胚中,眼瞼未能閉合是因?yàn)檠鄄€上皮內(nèi)肌動(dòng)蛋白束的裝配缺陷所致。
3 Smad4為c-Jun磷酸化及EGFR核轉(zhuǎn)位所必需
E15.5的野生小鼠胚眼瞼,c-Jun染色存在于瞼緣細(xì)胞胞核,見(jiàn)圖3 A,p-c-Jun染色亦呈強(qiáng)陽(yáng)性,見(jiàn)圖3 C,而變異小鼠有著類似的c-Jun染色模式,見(jiàn)圖3 B,但p-c-Jun染色明顯變?nèi)酰胰旧?xì)胞顯著減少,僅有少數(shù)靠近瞼結(jié)膜的細(xì)胞染色陽(yáng)性,見(jiàn)圖3 D。EGFR染色顯示,在Smad4野生小鼠,EGFR主要存在于瞼緣細(xì)胞細(xì)胞核內(nèi),見(jiàn)圖3 E,而在Smad4變異小鼠,EGFR染色出現(xiàn)在瞼緣細(xì)胞的核周胞漿,見(jiàn)圖3 F。我們推測(cè),Smad4在c-Jun的完全活化中起了重要的作用,Smad4的缺失導(dǎo)致其磷酸化過(guò)程受損,而無(wú)法發(fā)揮其足夠的轉(zhuǎn)錄因子功能;同時(shí)受損的還有 EGFR的核轉(zhuǎn)位,因此可能Smad4在EGF信號(hào)通路中也發(fā)揮了一定的作用。
4 Le-Cre條件性Smad4基因敲除鼠眼瞼閉合不能與細(xì)胞增殖或凋亡無(wú)關(guān)
在E15.5,TUNEL標(biāo)記的凋亡細(xì)胞在野生型和變異小鼠眼瞼瞼緣無(wú)明顯差別,見(jiàn)圖4A、B、C、D,E14.5的BrdU標(biāo)記在兩者間也相似,見(jiàn)圖3 E、F,因此可推測(cè)Smad4的缺失導(dǎo)致的眼瞼閉合不能不涉及細(xì)胞增殖的減少或細(xì)胞死亡的增加。
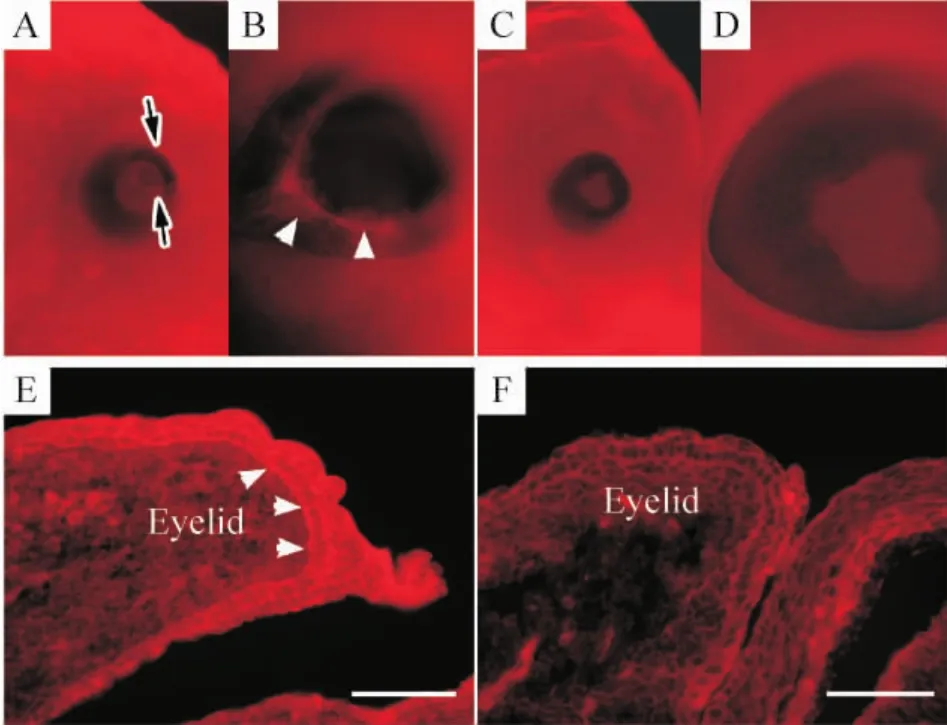
Figure 2.Absence of F - actin surrounding the mutant eyelids.A,B:whole mount phalloidin staining of eyelids from a wild-type embryo at E15.5 showed F-actin bundles(white arrowheads)projecting latitudinally around the edge of the eyelids(black arrows);C,D:the F-actin bundles were not present in the eyelids of the Smad4 CKO mutant;E:phalloidin intensely interacted with F-actin in the eyelid epithelium(arrowheads)of the wild-type embryo;F:F-actin bundles were not detected in the eyelid epithelium of the mutant embryo.Scale bar:100 μm.圖2 肌動(dòng)蛋白在變異眼瞼的缺失
討 論
胚胎發(fā)育中眼瞼的閉合通過(guò)上皮細(xì)胞移行來(lái)實(shí)現(xiàn),而眼瞼上皮細(xì)胞形態(tài)的改變,以及眼瞼上皮內(nèi)肌動(dòng)蛋白應(yīng)力纖維和絲足的形成這些都是上皮細(xì)胞移行的關(guān)鍵特征[5]。上皮移行一旦受損,將會(huì)導(dǎo)致EOB的發(fā)生。
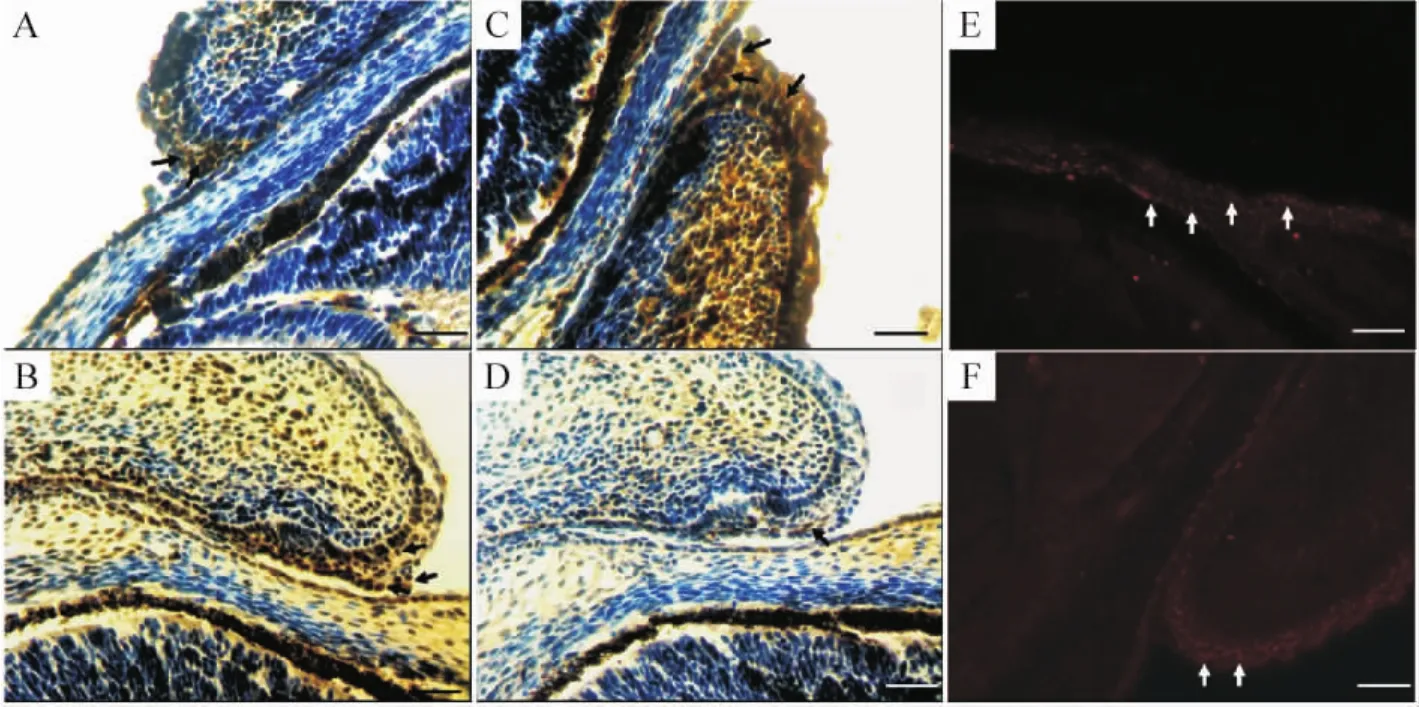
Figure 3.Immunohistochemical staining of eyelids(×400).A,B:strong nuclear staining for c-Jun in the eyelids of wild-type(A)and mutant(B)embryos at E15.5,no difference was observed;C,D:staining for embryo:in the Smad4 wild-type embryo,strong nuclear staining was observed in majority of epithelial cells in eyelids(C),but only a few cells were stained in Smad4 CKO embryo;E,F(xiàn):staining for EGFR:in the wild - type embryo,staining was found in the nuclei.Perinuclear staining was observed in the eyelid of mutant embryos with weaker immunoreactivity.Scale bar:100 μm.圖3 眼瞼免疫組化染色
基因敲除技術(shù)的發(fā)展為人們探尋眼瞼閉合的機(jī)制提供了一個(gè)良好的工具,借此也發(fā)現(xiàn)了很多基因敲除鼠可導(dǎo)致EOB表型,如纖維生長(zhǎng)因子10(fiber growth factor 10,F(xiàn)GF10)及其受體 FGFR2(fiber growth factor receptor 2)、激活素 β -B、MEK激酶1(MEKK1)、c-Jun N末端激酶(c-Jun N-terminal protein kinase,JNK)、c- Jun、EGFR 家族成員的 HB-EGF(heparin-binding EGF-like growth factor)和轉(zhuǎn)化生長(zhǎng)因子 α(transforming growth factor α,TGF-α)及其受體EGFR(epidermal growth factor receptor)、叉頭轉(zhuǎn)錄因子 Foxc1及 Foxc2、骨形態(tài)形成蛋白(bone morphogenetic protein,BMP)I型受體、BMP 相關(guān)的 R - Smads、Wnt拮抗劑 Dkk2 等[4,6-21],這些信號(hào)分子基本上歸屬于3個(gè)信號(hào)通路:TGFβ/激活素-MEKK1-JNK/p38信號(hào)通路、TGFα/EGFR-ERK信號(hào)通路及經(jīng)典的BMP信號(hào)通路,因此目前已經(jīng)明確胚胎眼瞼閉合至少需要3個(gè)信號(hào)通路的調(diào)控,涉及到眾多的基因,這些基因或信號(hào)通路的破壞或改變都可能會(huì)引起EOB的發(fā)生,普遍認(rèn)為FGF10是眼瞼發(fā)育的始動(dòng)因素,通過(guò)調(diào)控激活素 β-B、TGFα/HB-EGF以及BMP信號(hào)通路來(lái)控制眼瞼上皮的增生和移行,從而確保眼瞼的適當(dāng)發(fā)育[4]。
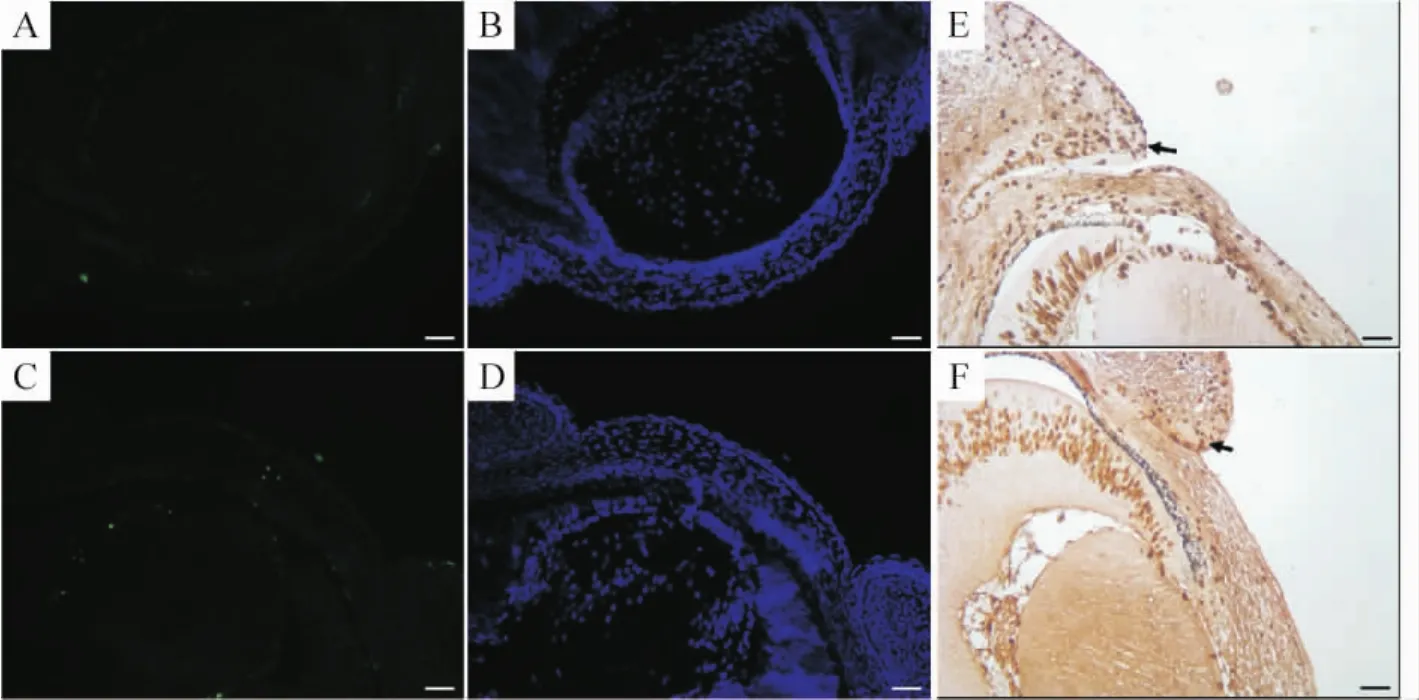
Figure 4.TUNEL staining and BrdU labeling of eyelids(×400).TUNEL staining in the eyelid epithelium of Smad4 wild-type(A,B)and CKO(C,D)embryos at E15.0 showed no difference in the number of apoptotic cells.BrdU labeling in the eyelid epithelium of Smad4 wild-type(E)and CKO(F)embryos at E14.5 also showed no difference in the number of BrdU-labeled cells.Scale bar:100 μm.圖4 眼瞼凋亡染色及BrdU標(biāo)記
有研究表明,在眼瞼發(fā)育過(guò)程中,眼瞼間質(zhì)內(nèi)的FGF10向眼瞼外胚層發(fā)出信號(hào),后者增殖并動(dòng)員表皮生長(zhǎng)因子(epidermal growth factor,EGF)和激活素(activin)信號(hào)通路,而后再激活ERK或MEKK1-JNK信號(hào)級(jí)聯(lián)通路,從而控制細(xì)胞骨架的重組,如誘導(dǎo)肌動(dòng)蛋白絲形成肌動(dòng)蛋白束像壓力纖維(stress fiber),這種纖維在推動(dòng)上皮層的運(yùn)動(dòng)中起主要作用,因此在眼瞼的融合過(guò)程中,肌動(dòng)蛋白重組是眼瞼融合過(guò)程中細(xì)胞遷移所必需而且關(guān)鍵的一個(gè)細(xì)胞活動(dòng)[4-5]。我們的結(jié)果提示在 Smad4基因敲除鼠胚中,眼瞼未能閉合是因?yàn)檠鄄€上皮內(nèi)肌動(dòng)蛋白束的裝配缺陷所致。
c-Jun是眼瞼閉合的關(guān)鍵調(diào)控因素,其磷酸化不僅為BMPs信號(hào)通路和TGFβ/激活素-MEKK1-JNK信號(hào)通路所必需,而且對(duì)于TGFα/EGFR-ERK信號(hào)通路的激活有誘導(dǎo)作用,在眼瞼閉合的分子機(jī)制中不可或缺[4-7]。Huang 等[4]在他們最新的研究中發(fā)現(xiàn),F(xiàn)GF調(diào)控的BMP信號(hào)通路在眼瞼閉合中發(fā)揮必需的作用,他們的結(jié)果提示BMPs通過(guò)經(jīng)典的R-Smad/Smad4通路,與激活素 β-B通路共同促進(jìn)c-Jun的最大磷酸化并介導(dǎo)磷酸化p-c-Jun的核轉(zhuǎn)位,同時(shí)該信號(hào)通路還激活了眼瞼閉合所必需的轉(zhuǎn)錄因子Foxc1和Foxc2的表達(dá),從而調(diào)控眼瞼發(fā)育的閉合過(guò)程。他們還認(rèn)為FGF調(diào)控的TGF α通路與BMP信號(hào)通路之間也存在偶合,且激活素 β-B,TGFα/HB -EGF,音猬因子(Sonic Hedgehog,Shh)以及BMP信號(hào)通路形成了一個(gè)復(fù)雜的調(diào)控網(wǎng)絡(luò),確保瞼緣上皮細(xì)胞的適當(dāng)遷移和融合。
與Huang等的研究類似,我們也在Le-Cre特異性Smad4基因敲除鼠中觀察到了EOB表型,這種EOB表型并非是眼瞼的過(guò)早開(kāi)放,也與細(xì)胞增殖或細(xì)胞凋亡的異常無(wú)關(guān),而是Smad4在眼瞼上皮細(xì)胞失活后,眼瞼上皮細(xì)胞肌動(dòng)蛋白束合成缺陷,上皮細(xì)胞移行受影響,從而導(dǎo)致了眼瞼融合缺陷。我們也發(fā)現(xiàn)Smad4對(duì)c-Jun磷酸化有影響,但并不影響c-Jun本身的核表達(dá)。與Huang等發(fā)現(xiàn)p-c-Jun在Smad4變異小鼠眼瞼上皮細(xì)胞核周表達(dá)不同,我們發(fā)現(xiàn)雖然磷酸化c-Jun的表達(dá)明顯降低,陽(yáng)性細(xì)胞數(shù)減少,但其表達(dá)仍存在于少數(shù)細(xì)胞核內(nèi),這意味著磷酸化c-Jun的核轉(zhuǎn)位并未受到影響,僅Smad4失活不足以完全抑制c-Jun磷酸化,應(yīng)該存在其它信號(hào)通路調(diào)控c-Jun的磷酸化,這與Zhang等[21]的研究結(jié)果相似,他們認(rèn)為c-Jun的磷酸化和功能部分依賴于激活素β-B的非Smads依賴信號(hào)通路,同時(shí)也與眼瞼融合受多個(gè)信號(hào)通路調(diào)控相吻合。我們還觀察到EGFR在Smad4變異小鼠眼瞼上皮的表達(dá)發(fā)生改變,Smad4失活后,EGFR未能出現(xiàn)在核內(nèi),而是位于核周胞漿,且表達(dá)減弱。有研究發(fā)現(xiàn),在眼周皮膚的移行中,EGF家族成員通過(guò)激活ERK信號(hào)通路而促進(jìn)眼周皮膚的移行,而EGF級(jí)聯(lián)上游的c-Jun 磷酸化可上調(diào) EGFR - ERK 通路[5,21];Li等[6]及Zenz等[7]的研究又表明,c-Jun在表皮的失活可影響EGFR信號(hào)通路的激活,因此我們推測(cè)Smad4的失活可能也同時(shí)影響了EGFR信號(hào)通路,亦或與c-Jun磷酸化改變有關(guān),而且Smad4可能不止在控制眼瞼閉合的某一個(gè)信號(hào)通路中起信號(hào)轉(zhuǎn)導(dǎo)作用,而是介入了多個(gè)信號(hào)通路,從而調(diào)控肌動(dòng)蛋白應(yīng)力纖維的裝配和眼瞼上皮的移行。
眼瞼發(fā)育的調(diào)控是一個(gè)相當(dāng)復(fù)雜的過(guò)程,涉及眾多信號(hào)通路,構(gòu)成一個(gè)復(fù)雜的網(wǎng)絡(luò),需要進(jìn)一步的研究來(lái)完全闡明這些通路的功能及相互作用,這些研究不但有利于我們對(duì)眼瞼融合更全面的認(rèn)識(shí),而且對(duì)于我們理解其它依賴于上皮融合的形態(tài)學(xué)發(fā)生如神經(jīng)管、視裂、晶體泡及顎架也大有裨益,同時(shí)對(duì)于尋找上皮性損傷的修復(fù)機(jī)制及治療手段有所借鑒。
[1]Swindell EC,Liu C,Shah R,et al.Eye formation in the absence of retina[J].Dev Biol,2008,322(1):56 -64.
[2]Findlater GS,McDougall RD,Kaufman MH.Eyelid development,fusion and subsequent reopening in the mouse[J].J Anat,1993,183(Pt 1):121 -129.
[3]Kaufman MH.The atlas of mouse development[M].1st ed.London:Academic Press,2002.525.
[4]Huang J,Dattilo LK,Rajagopal R,et al.FGF -regulated BMP signaling is required for eyelid closure and to specify conjunctival epithelial cell fate[J].Development,2009,136(10):1741-1750.
[5]Xia Y,Kao WWY.The signaling pathways in tissue morphogenesis:a lesson from mice with eye-open at birth phenotype[J].Biochem Pharmacol,2004,68(6):997 -1001.
[6]Li G,Gustafson-Brown C,Hanks SK,et al.c-Jun is essential for organization of the epidermal leading edge[J].Dev Cell,2003,4(6):865 -877.
[7]Zenz R,Scheuch H,Martin P,et al.c-Jun regulates eyelid closure and skin tumor development through EGFR signaling[J].Dev Cell,2003,4(6):879 -889.
[8]Gage PJ,Qian M,Wu D,et al.The canonical Wnt signaling antagonist DKK2 is an essential effector of PITX2 function during normal eye development[J].Dev Biol,2008,317(1):310-324.
[9]Kidson SH,Kume T,Deng K,et al.The forkhead/winged-h(huán)elix gene,Mf1,is necessary for the normal development of the cornea and formation of the anterior chamber in the mouse eye[J].Dev Biol,1999,211(2):306-322.
[10]Kume T,Deng KY,Winfrey V,et al.The forkhead/winged helix gene Mf1 is disrupted in the pleiotropic mouse mutation congenital hydrocephalus[J].Cell,1998,93(6):985-996.
[11]Li C,Guo H,Xu X,et al.Fibroblast growth factor receptor 2(Fgf2)plays an important role in eyelid and skin formation and patterning[J].Dev Dyn,2001,222(3):471-483.
[12]Luetteke NC,Qiu TH,Peiffer RL,et al.TGF alpha deficiency results in hair follicle and eye abnomalities in targeted and waved -1 mice[J].Cell,1993,73(2):263 -278.
[13]Luetteke NC,Phillips HK,Qiu TH,et al.The mouse waved-2 phenotype results from a point mutation in the EGF receptor tyrosine kinase[J].Genes Dev,1994,8(4):399-413.
[14]Miettinen PJ,Berger JE,Meneses J,et al.Epithelial immaturity and multiorgan failure in mice lacking epidermal growth factor receptor[J].Nature,1995,376(6536):337-341.
[15]Mine N,Iwamoto R,Mekada E.HB-EGF promotes epithelial cell migration in eyelid development[J].Development,2005,132(19):4317 -4326.
[16]Smith RS,Zabaleta A,Kume T,et al.Haploinsufficiency of the transcription factors FOXC1 and FOXC2 results in aberrant ocular development[J].Hum Mol Genet,2000,9(7):1021-1032.
[17]Takatori A,Geh E,Chen L,et al.Differential transmission of MEKK1 morphogenetic signals by JNK1 and JNK2[J].Development,2008,135(1):23 - 32.
[18]Tao H,Shimizu M,Kusumoto R,et al.A dual role of FGF10 in proliferation and coordinated migration of epithelial leading edge cells during mouse eyelid development[J].Development,2005,132(14):3217 -3230.
[19]Vassalli A,Matzuk MM,Gardner HA,et al.Activin/inhibin βB subunit gene disruption leads to defects in eyelid development and female reproduction[J].Genes Dev,1994,8(4):414-427.
[20]Weston CR,Wong A,Hall JP,et al.The c-Jun NH2-terminal kinase is essential for epidermal growth factor expression during epidermal morphogenesis[J].Proc Natl Acad Sci USA,2004,101(39):14114-14119.
[21]Zhang L,Wang W,Hayashi Y,et al.A role for MEK kinase 1 in TGF-beta/activin-induced epithelium movement and embryonic eyelid closure[J].EMBO J,2003,22(17):4443-4454.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
中國(guó)生殖健康(2019年3期)2019-02-01 06:12:26
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
海軍航空大學(xué)學(xué)報(bào)(2015年3期)2015-11-11 17:20:00
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
七彩語(yǔ)文·畫刊(2012年3期)2012-04-29 00:00:00

- 中國(guó)病理生理雜志的其它文章
- 神經(jīng)生長(zhǎng)因子通過(guò)TrkANGFR信號(hào)途徑促進(jìn)肝細(xì)胞增殖*
- 中國(guó)病理生理學(xué)會(huì)2011年度工作總結(jié)
- 小鼠早期完整胚胎誘導(dǎo)子宮內(nèi)膜白血病抑制因子和整合素β3表達(dá)并提高子宮容受性*
- 干擾素誘導(dǎo)蛋白p204表達(dá)變化對(duì)大鼠血管平滑肌細(xì)胞增殖及p21表達(dá)的影響*
- 孕酮對(duì)氧糖剝奪損傷的PC12細(xì)胞的保護(hù)作用*
- 炎癥對(duì)脂肪酸負(fù)荷的腎細(xì)胞FAT/CD36表達(dá)的影響*