肌肉免疫DC-EBV-LMP2誘導(dǎo)免疫應(yīng)答的研究
2012-08-04 06:52:14杜海軍尉秀霞葉樹清
中國醫(yī)藥指南 2012年30期
杜海軍 王 湛 尉秀霞 周 玲* 馬 晶 馮 霞 葉樹清 曾 毅*
(中國疾病預(yù)防控制中心 病毒病預(yù)防控制所,傳染病預(yù)防與控制國家重點實驗室,北京100052)
樹突狀細胞(Dendritic cell,DC)是1973年美國洛克菲勒大學(xué)的Steinman和Cohn教授從小鼠的外周淋巴器官首次發(fā)現(xiàn)的[1],是體內(nèi)功能最強的專職抗原提呈細胞(Antigen present cell,APC),也是唯一能激活初始型T細胞的APC,被稱為天然“免疫佐劑”,在抗感染、抗腫瘤、移植排斥等過程中發(fā)揮重要作用[2]。目前以DC為基礎(chǔ)的疫苗在治療腫瘤方面,尤其在某些晚期腫瘤等的臨床治療方面已經(jīng)取得一些的成效[3-7]。鼻咽癌(Nasoparyngeal carcinoma,NPC)是我國南方一些省(自治區(qū))的常見惡性腫瘤之一。目前放射治療是鼻咽癌治療的基本方法,但中晚期鼻咽癌治療后常常復(fù)發(fā)或向遠處轉(zhuǎn)移,對于復(fù)發(fā)和轉(zhuǎn)移的患者在臨床上尚無有效治療方案。EBV-LMP2是NPC細胞表達的病毒抗原之一,是誘導(dǎo)特異性免疫應(yīng)答的理想靶抗原。以DC為基礎(chǔ)的疫苗其免疫方式報道多為靜脈回輸、淋巴結(jié)注射和皮內(nèi)(下)注射[8-11]。我們以肌肉免疫方式注射DC-EBV-LMP2,研究LMP2誘導(dǎo)的特異性CTL。
1 材料與方法
1.1 材料與試劑
Balb/C鼠購自軍事醫(yī)學(xué)科學(xué)院實驗動物中心。優(yōu)級胎牛血清購自Invitrogen公司。rAd-LMP2:深圳清華源興生物醫(yī)藥科技有限公司生產(chǎn)。LMP2單抗購于SantaCruz公司;熒光標記羊抗鼠IgG及HRP標記羊抗兔IgG購自北京中衫生物技術(shù)公司;流式檢測抗體購自Biolegend公司;重組小鼠細胞因子購自Peprotech公司;ELispot Kit購自BD公司;細胞培養(yǎng)液由病毒病預(yù)防控制所配液室提供。
1.2 小鼠骨髓細胞分離與DC的誘導(dǎo)培養(yǎng)
斷頸法處死小鼠,取出小鼠股骨骨髓,制成成單細胞懸液,以小鼠淋巴細胞分離液(達科為)分離骨髓細胞(2000rpm/min,30min)。洗滌2次(1500rpm/min,10min),獲得小鼠骨髓細胞。以4×106/孔濃度接種在6孔培養(yǎng)板中,37℃5%的CO2培養(yǎng)箱中培養(yǎng)3h。輕微晃動培養(yǎng)液,吸掉上清中的懸浮細胞,以無血清1640輕輕沖洗3~4遍,加入含細胞因子(rmGM-CSF200U,rmIL-410U,rmTNF-α100U)10%胎牛血清的RPMI1640,37℃5%CO2培養(yǎng),3d后,半量更換含細胞因子培養(yǎng)液,繼續(xù)培養(yǎng)至7d。
1.3 DC-EBV-LMP2制備
用移液管吹打細胞,收集懸浮細胞團,1500rpm/min,離心10min,獲得小鼠的DC。以100Tcid50的rAd-LMP2感染吸附DC1小時(體積在0.5~1mL內(nèi)),然后在含細胞因子(rmGM-CSF200U,rmIL-410U,rmTNF-α200U)10%胎牛血清的RPMI1640作用下,培養(yǎng)48h。第9天收集成熟的樹突狀細胞(1500rpm/min,10 min)。
1.4 DC表達LMP2的檢測:
取5μL DC-EBV-LMP2懸液滴在載玻片的小孔內(nèi),晾干后,冷丙酮4℃固定。分別以LMP2單抗(1?100稀釋)、生物素標記標IgG抗體(1∶100稀釋)、辣根過氧化物酶標記標抗生物素的IgG抗體(1∶100稀釋)與涂片細胞作用(37℃,40min)。每次PBS沖洗3遍。最后將細胞片浸泡于聯(lián)苯二胺顯色液中1min,然后在顯微鏡下觀察LMP2在DC中表達。
1.5 DC-EBV-LMP2免疫Balb/C鼠實驗
將Balb/C鼠30只,分為3組,按(表1)的設(shè)計進行小鼠免疫。在初次免疫后的第5周和第8周,處死小鼠(每次每組5只),取脾臟分離淋巴細胞進行LMP2特異性細胞免疫檢測。
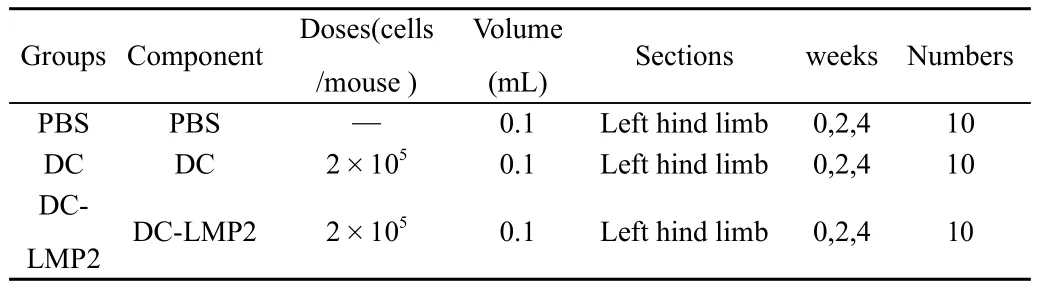
表1 小鼠免疫方案設(shè)計
1.6 小鼠IFN-γElispot的檢測
按照試BD劑盒使用說明,將包被抗體,以2.5×105/孔接種細胞,LMP2多肽(AAALALLASLIL和ASCFTASVSTVV各2μL/孔)刺激物5μg/mL,設(shè)陽性對照1×105/孔(加PHA或ConA刺激1μg/孔)、陰性對照2.5×105/孔、空白對照(加100μL/孔培養(yǎng)液)。按說明書步驟依次加入檢測抗體、酶標的鏈霉親和素和顯色液。用BiosysBioreader4000PRO自動讀板儀讀取斑點,并作統(tǒng)計分析。
1.7 CD8+T/CD3+T亞群檢測
按說明書在淋巴細胞中加入熒光標記CD8和CD3抗體,同時設(shè)置陰性對照(空白)、單色陽性對照及多色檢測樣本,室溫孵育20min。PBS洗滌2次,200目濾網(wǎng)過濾后,流式細胞儀檢測。
2 結(jié)果
2.1 DC制備與LMP2在DC中的表達
在細胞因子誘導(dǎo)下,小鼠骨髓分離的單核細胞逐步分化成樹突狀細胞。以100 MOI rAd-LMP2感染DC 48h后,免疫酶法可以檢測到LMP2在DC中表達(圖1)。
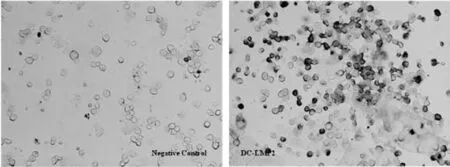
圖1 LMP2在小鼠骨髓來源DC中表達檢測
2.2 DC-EBV-LMP2誘導(dǎo)特異性免疫應(yīng)答
經(jīng)過0、2、4周免疫后,用IFN-γElispot法測定LMP2特異性CTL,結(jié)果顯示DC-EBV-LMP2免疫組特異性CTL(308±167)數(shù)量明顯高于PBS組(18±6)和DC組(26±12)。統(tǒng)計學(xué)分析:P<0.01具有顯著差異(圖2)。在免疫后的第8周仍然檢測到,只是較第5周有所降低(圖3)。
2.3 免疫后細胞亞群變化
以流式細胞儀檢測各實驗組中CD8+T/CD3+T細胞亞群的比值,數(shù)據(jù)顯示在小鼠0,2,4周免疫后的第5周,DC-EBV-LMP2免疫組CD8+T/CD3+T細胞亞群的比值較PBS和DC組有所升高,但統(tǒng)計學(xué)軟件分析P>0.05,不具有統(tǒng)計學(xué)意義。而在第八周再次檢測CD8+T/CD3+T細胞亞群的比值時是顯示,DC-EBV-LMP2免疫組CD8+T/CD3+T細胞亞群的比值低于PBS和DC組。
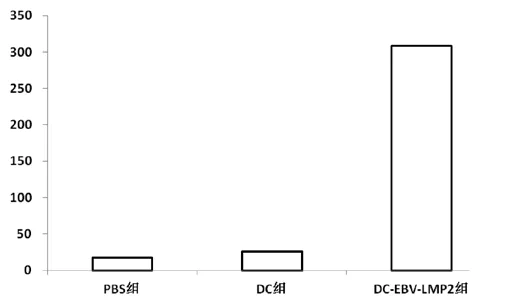
圖2 小鼠0、2、4周免疫后第五周IFN-γElispot檢測結(jié)果
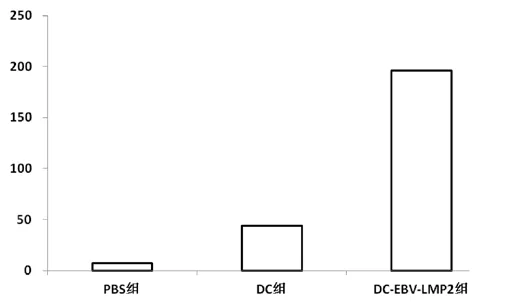
圖3 小鼠免疫后第八周IFN-γ Elispot檢測結(jié)果
3 討論
我們以前研究結(jié)果表明:以pcDNAⅢ-LMP2真核表達質(zhì)粒免疫小鼠可誘發(fā)LMP2特異性細胞和體液免疫[12]。rAd-LMP2以灌胃、肌肉注射和滴鼻的方式感染小鼠,均可誘發(fā)小鼠針對LMP2的特異性體液免疫和細胞免疫[13,14]。EBV-LMP2DNA疫苗、腺相關(guān)病毒(adenoassociated virus,AAV)疫苗、非復(fù)制5型腺病毒疫苗(Ad5)聯(lián)合免疫Balb/C小鼠能夠更好地誘導(dǎo)機體產(chǎn)生特異性細胞免疫應(yīng)答[15]。rAd-LMP2肌肉免疫恒河猴可以誘導(dǎo)EBV-LMP2特異性的細胞免疫應(yīng)答及抗體應(yīng)答,且免疫應(yīng)答水平的高低與病毒劑量的高低相關(guān)[16]。rAd5F35-LMP2重組腺病毒疫苗在恒河猴體內(nèi)誘導(dǎo)EBV-LMP2特異性的免疫應(yīng)答[17]。國內(nèi)外眾多的文獻報道,DC的免疫途徑有皮下、皮內(nèi)、淋巴結(jié)注射和靜脈回輸?shù)龋匆娪屑∪饷庖叩膱蟮馈1狙芯康膶嶒灲Y(jié)果證實DC-EBV-LMP2肌肉免疫小鼠后能夠誘導(dǎo)LMP2特異性的細胞免疫應(yīng)答。由于LMP2刺激產(chǎn)生了較強的細胞免疫應(yīng)答,生成大量的直接具有殺傷效應(yīng)的細胞即細胞毒T細胞,造成了免疫后DC-EBV-LMP2組較PBS和DC組的CD8+T細胞亞群有所增高,DC免疫組與PBS組比較也有所升高,可能為DC刺激所致。隨著特異性CTL與靶細胞結(jié)合后釋放穿孔蛋白使靶細胞破裂,免疫應(yīng)答結(jié)束,大部分細胞死亡,微量變成記憶T細胞長期存在。
由于記憶T細胞存在,使我們在第八周仍能夠檢測到LMP2特異性的免疫應(yīng)答,雖然隨著時間的推移,絕大多數(shù)的CTL已經(jīng)死亡,因而免疫應(yīng)答強度有所降低,流式細胞儀檢測DC-EBV-LMP2組CD8+T細胞亞群比值明顯低于其他兩組,恰好是一個有力的證據(jù)。除此之外,我們注意到免疫后第八周CD8+T細胞亞群水平整體上有所降低,一方面由于小鼠自身CD8+T細胞亞群經(jīng)過擴增后,除記憶細胞外,其余的大部分凋亡細胞所致,另一個原因可能是不同老鼠實驗批次間的差異造成,隨著時間延長小鼠“衰老”也可能起一定的作用。在實驗中我們未檢測CD4+T細胞在小鼠免疫應(yīng)答過程中的變化,尤其是CD25+CD4+T細胞亞群在特異性細胞免疫應(yīng)答過程中,起著維持自身耐受和避免免疫反應(yīng)過度損傷機體的重要作用,也許與CD8+T/CD3+T的比值變化相關(guān)。
從我們目前的實驗結(jié)果分析,雖然DC-EBV-LMP2組誘導(dǎo)的特異性免疫應(yīng)答的水平明顯高于PBS和DC組,但組內(nèi)小鼠間的數(shù)值差異較大,在今后的實驗中需要進一步增加每組小鼠的數(shù)量,減小鼠間差異。免疫后隨著時間的延長,免疫應(yīng)答的強度衰減,這提示我們在今后的臨床免疫治療中,需要再次免疫或與其它疫苗聯(lián)合應(yīng)用來維持細胞免疫的強度。我們正在進行這一方面的探索。
[1] Steinman RM,Cohn ZA.Identification of a novel cell type in peripheral lymphoid organs of mice I Morphology quantitation tissue distribution[J].J Exp Med,1973,11(37):1142-1162.
[2] Steinman RM.The dendritic cell system and its role in immunogenicity[J].Annu Rev Immunol,1991,l9(2):271-296.
[3] Nestle FO,Alijagic S,Gilliet M.tumor lysate-pulsed dendritic cells[J].Nat Med,1998,4(3):328-332.
[4] O`Rourke MG,Johnson MK,Lanaqan CM,et al.Dendritic cell immunotherapy for stage Ⅳ melanoma[J].Melanoma Res,2007,17(5):316-322.
[5] Fong L,Brockstedt D,Benike C,et al.Dendritic cell-based xenoantigen vaccination for prostate cancer immunotherapy[J].J Immunol,2001,67(12): 7150-7156.
[6] Yamaquchi Y,Ohta K,Kawabuchi Y,et al.Feasibility study of adoptive immunotherapy dendritic cell-activated killer(PDAK)cells[J].Anticancer Res,2005,l25(3c):2407-2415.
[7] Avigan D,Vasir B,Gong J,et al.Fusion cell vaccination of patients with metastatic breast[J].Clin Cancer Res,2004,10(14):4699-4708.
[8] Mackensen A,Meidenbauer N,Vogl S,et al.PhaseⅠstudy of adoptive T-cell therapy using antigen-specific[J].J Clin Oncol,2006,24(31):5060-5069.
[9] Lambert LA,Gibson GR,Maloney M,et al.Intranodal immunization with tumor lysate-pulsed [J].Cancer Res,2001,61(2):641-646.
[10] Lin CL,Lo WF,Lee TH,et al.Immunization with Epstein-Barr Virus peptide-pulsed dendritic cells induces carcinoma[J].Cancer Res,2002,62(23):6952-6958.
[11] Zuo JM,Zhou L,Chen ZJ,et al.Induction of cytotoxic T lymphocyte responses nasopharyngeal carcinoma[J].J.Microbiol immunol,2006,l4(1):41-48.
[12] 朱偉嚴,周玲,王琦,等.EB病毒潛伏膜蛋白2DNA疫苗的構(gòu)建及其初步免疫效果觀察[J].中華微生物學(xué)和免疫學(xué)雜志,2002,22(2):185-190.
[13] 姚佳偉,周玲,王琦,等.EB病毒潛伏膜蛋白2重組腺腺病毒的構(gòu)建及其免疫效果的研究[J].中國腫瘤,2003,12(1):45-47.
[14] 左建民,周玲,王曾毅,等.含EBV-LMP2基因重組腺病毒疫苗的構(gòu)建及其誘導(dǎo)CTL應(yīng)答的初步探討[J].中華微生物學(xué)和免疫學(xué)雜志,2003,23(6):446-449.
[15] 楊松梅,王湛,周玲,等.攜帶EBV-LMP2基因的DNA疫苗、腺相關(guān)病毒疫苗和腺病毒疫苗免疫小鼠的特異性細胞免疫應(yīng)答[J].生命科學(xué),2009,39(4):342-345.
[16] 王湛,周玲,吳小兵,等.Ad-LMP2重組腺病毒疫苗在恒河猴體內(nèi)免疫效果的研究[J].中華實驗和臨床病毒學(xué)雜志,2006,20(2):63-65.
[17] 莫武寧,周玲,吳小兵,等.rAd5F35-LMP2重組腺病毒疫苗免疫恒河猴誘導(dǎo)免疫應(yīng)答[J].中華實驗和臨床病毒學(xué)雜志,2007,21(3):226-228.
猜你喜歡
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
湖北師范大學(xué)學(xué)報(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(2014年3期)2014-07-31 18:57:34
