甘藍SCR、THL與SRK交替相互作用的酵母雙雜交檢測
2012-08-08 01:23:22楊永軍張賀翠薛麗琰常登龍朱利泉
中國蔬菜 2012年24期
楊永軍 張賀翠 楊 昆 薛麗琰 常登龍 朱利泉
(西南大學農學與生物科技學院植物生理生化實驗室,重慶 400716)
植物的自交不親和性(self-incompatibility,SI)是其為了避免自花受精而進化出的重要遺傳機制。甘藍(Brassica oleraceaL.)SI的分子過程是由存在于花粉中的S-位點富含半胱氨酸蛋白(S-locus cystein-rich protein,SCR),亦稱為S位點蛋白11(S-locus protein11,SP11)識別結合柱頭上的S受體激酶基因(Sreceptor kinase gene,SRK)啟動,將原本結合于SRK上的負調控因子類硫氧還蛋白(thioredoxin-like protein,THL)釋放出來,然后通過目前尚未完全知曉的分子過程實現自交不親和性。SRK由S結構域(S域)、跨膜結構域以及激酶結構域3個結構域組成(Stein et al.,1991),SRK的胞外區域與SCR相識別,且基因序列與SLG具有較高的同源性(Takayama et al.,2001);其本身又具有較高的多態性,與S位點緊密連鎖,呈單倍型一致性。根據S-位點上SLG基因的結構,蕓薹屬自交不親和復等位基因可分為兩大類:class-Ⅰ自交不親和復等位基因,SLG沒有內含子;class-Ⅱ型S等位基因,SLG基因的3′末端附近有1個內含子(Hatakeyama et al.,1998a;Kusaba & Nishio,1999)。Ⅰ型和Ⅱ型的S基因的分化較早,差異較大(Kusaba et al.,1997),Ⅱ型不同物種的SLG基因之間同源性在86%以上,而Ⅰ型和Ⅱ型SLG基因之間同源性僅60%~70%。Nishio等(1996)根據這兩類SLG基因序列特點設計特異引物,通過PCR擴增就能將這兩類SLG基因區分開來。Stein等(1991)發現,在同一單倍型中S結構域中的SRK與SLG表現出極高的相似性,擁有很相似的單倍型和相似的S位點特異性,在序列上SLG可能與其他相似的SRK單倍型聯系更加緊密。然而甘藍S-2和S-6的單倍型數據顯示,Ⅱ型與Ⅰ型序列差別很大(Kusaba et al.,1997)。研究還發現許多甘藍類作物S等位基因兼有class-Ⅰ和class-Ⅱ型(Chen & Nasrallah,1990;Tantikanjana et al.,1993),在Ⅰ型和Ⅱ型兼有的植物中,所有Ⅱ型S復等位基因都屬于花粉隱性表現型,包括B. rapa中的S-29、S-40、S-44和S-60單元型(Hatakeyama et al.,1998b;Takasaki et al.,2000)和B. oleracea中的S-2、S-5和S-15單元型(Chen & Nasrallah,1990;Scutt & Croy,1992;Cabrillaca et al.,1999),而大多數 class-Ⅰ型S復等位基因屬于花粉顯性表現型(Nasrallah et al.,1991);研究發現Ⅰ型的SCR基因為顯性基因,Ⅱ型的SCR基因為隱性基因,在轉錄過程中Ⅱ型的SCR基因的轉錄受到了抑制,因此植物表現顯隱性的關系決定于S位點基因的轉錄情況(Shiba et al.,2002)。Ⅰ型和Ⅱ型S等位基因存在較高的保守區,同時也存在著較大的差異(Kusaba et al.,1997),這些差異涉及從授粉到受精過程中發生的花粉配體和雌蕊受體之間的相互作用以及相互作用的強弱關系(Kachroo et al.,2002;Ivanov et al.,2010)。S位點的基因結構不同而使Ⅰ型和Ⅱ型產生差異的分子機制尚不清楚,甘藍兩種類型的SRK與其下游的識別因子(SCR和THL1)之間的相互作用是否有區別,以及它們之間相互作用的強度又有什么關系還未見報道。因此建立一個適用于 class-Ⅱ型配體和受體之間,以及與其下游的THL1/THL2之間的蛋白質相互作用平臺,進行蛋白質間相互作用機理的研究,對控制整個自交不親和信號傳導途徑,以及確定class-Ⅰ和class-Ⅱ型自交不親和程度具有重要意義。本試驗擬構建class-Ⅱ型SCR2、eSRK2酵母雙雜交表達載體和class-Ⅰ型SRKJ激酶域,THL1酵母雙雜交表達載體,通過其毒性和自激活檢測來確定自交不親和性表型所依賴的3個起始信號傳導元件之間識別程度的差異,進而通過激活酵母雙雜交的報告基因所表達的多少來確定其相互作用程度。并通過生物信息學分析確定其Ⅰ型和Ⅱ型歸屬,以期能確定class-Ⅱ型內部的eSRK-SCR的互作及其與class-Ⅰ型之間THL、SCR與SRK存在的相互作用及其作用程度,為深入研究甘藍自交不親和性信號傳導分子過程提供參考。
1 材料與方法
1.1 材料
試驗于2011年7月~2012年1月在西南大學農學與生物科技學院植物生理生化實驗室進行。甘藍 B3、E1、D3是西南大學蔬菜學實驗室選育的材料。大腸桿菌(Escherichia coli)DH5α菌株為本實驗室保存。EcoRⅠ、SmaⅠ和BamHⅠ限制性內切酶,T4DNA連接酶購自 TaKaRa公司。RNA提取試劑盒、反轉錄試劑盒、TaqDNA聚合酶、DNA膠回收試劑盒及質粒提取試劑盒購自TransGen公司。酵母雙雜交系統(Matchmaker? Gold Yeast Two-Hybrid System)、x-α-gal、Aureobasidin A(以下簡稱AbA)及酵母培養基購自Clontech公司。DMSO購自Sigma公司。
1.2 SCR2、eSRK2、THL1的擴增
根據SCR2、eSRK2和THL1基因的結構特點,分別構建SCR2、eSRK2、THL1基因序列片段。依據GenBank中編號為AB067447的SP11-2序列和編碼為AJ306585的SRK序列分析和酶切圖譜,結合載體pGBKT7和pGADT7的多克隆位點,利用primer premier 6.0設計相應的RT-PCR引物。SCR2擴增的是第2個外顯子。eSRK2正向引物從成熟肽第1個氨基酸開始,并去掉信號肽;反向引物是依據eSRK-3、eSRK-7、eSRK-15、eSRK-28、eSRK-6、eSRK-36、eSRK-18、eSRK-12、eSRK-60、eSRK-2設計的特異引物(表1)。另外,參照劉東等(2003)擴增的引物和條件,擴增THL1編碼序列。SRK6(SRKJ)激酶域、eSRK28分別來自張賀翠等(2011)、楊紅等(2011)。
以反轉錄的花藥cDNA為模板擴增SCR2、eSRK2和THL1片段,eSRK2片段反應參數為94 ℃2 min;94 ℃ 2 min,55 ℃ 2 min,72 ℃ 45 s,30 個循環;72 ℃ 10 min,25 ℃ 1 min 終止。以反轉錄的柱頭cDNA為模板擴增eSRK2片段,反應參數為94 ℃ 2 min;94 ℃ 2 min,59 ℃ 60 s,30 個循環;72 ℃ 10 min,25 ℃ 1 min 終止。THL1片段的反應參數為 94 ℃ 2 min;94 ℃ 2 min,52 ℃ 2 min,72 ℃ 45 s,30個循環;72 ℃ 10 min,25 ℃ 1 min終止。擴增產物經1%瓊脂糖凝膠電泳檢測后,以DNA膠回收試劑盒純化回收。

表1 擴增SRK的S結構域和SCR的引物
1.3 酵母雙雜交重組表達載體的構建
用EcoRⅠ和SmaⅠ對擴增產物SCR2、THL1和酵母表達質粒pGBKT7進行雙酶切;用EcoRⅠ和BamHⅠ對eSRK2、SRKJ和酵母表達質粒pGADT7進行雙酶切;SCR2、THL1片段與pGBKT7連接,轉化E. coliDH5α感受態細胞,涂于LB平板上(含Kan+抗性);eSRK2、SRKJ片段與pGADT7連接,轉化E. coliDH5α感受態細胞,涂于LB平板上(含Amp+抗性)。37 ℃培養過夜,篩選陽性重組子,并進行菌液PCR和酶切鑒定,將含有插入目的片段的陽性克隆送北京華大基因研究中心測序。序列正確的重組質粒分別命名為pGBKT7-SCR2、pGBKT7-THL1、pGADT7-eSRK2和 pGADT7-SRKJ。
1.4 重組酵母雙雜交表達質粒的毒性檢測
根據酵母雙雜交系統使用說明書(Matchmaker? Gold Yeast Two-Hybrid System User Manual),采用LiAc法制備感受態酵母菌Y2HGold和Y187。用PEG/LiAc法將pGBKT7空載和重組誘餌載體pGBKT7-SCR2、pGBKT7-THL1同時轉化 Y2HGold酵母感受態細胞,30 ℃培養 3~5 d,觀察其在SD/-Trp篩選平板上的生長情況。采用同樣的方法將pGADT7空載和重組AD質粒pGADT7-eSRK2、pGADT7-SRKJ轉化Y187感受態細胞,觀察其在SD/-Leu平板上的生長情況。
1.5 重組誘餌載體的自激活檢測
用PEG/LiAc法將pGBKT7-SCR2轉化Y2HGold酵母感受態細胞,分別涂布于SD/-Trp、SD/-Trp/xα-gal、SD/-Trp/x-α-gal/AbA平板上;將 pGADT7-eSRK2轉化 Y187感受態細胞,分別涂布于SD/-Leu、SD/-Leu/x-α-gal平板上,30 ℃倒置培養 3~5 d,觀察 pGBKT7-SCR2、pGBKT7-THL1在選擇平板上的生長情況。以 pGBKT7-Lam×pGADT7-T為陰性對照,以 pGBKT7-p53×pGADT7-T為陽性對照。
1.6 SRKJ與 THL1、SCR2與 eSRK2、eSRK28與 SCR2、eSRK28與 SCR28相互作用檢測
分別挑取SD/-Trp、SD/-Leu平板上的菌落,將含有SCR2和eSRK2片段的pGBKT7-SCR2×pGADT7-eSRK2轉化菌落組合,同時接種于500 μL的2×YPDA液體培養基中,30 ℃振蕩培養18~20 h,在顯微鏡下檢測 Y2HGold與 Y187交配形成二倍體接合型的情況。以 pGBKT7-Lam×pGADT7-T為陰性對照,以 pGBKT7-p53×pGADT7-T為陽性對照。然后將融合菌液涂布到SD/-Trp-Leu(DDO)、SD/-Trp-Leu/x-α-gal/AbA(DDO/x/A)平板上,30 ℃培養3~5 d。最后將DDO/x/A平板上的藍色克隆劃線至SD/-His-Trp-Leu(TDO)、SD/-His-Trp-Leu/x-α-gal/AbA(TDO/x/A)和 SD/-Trp-Leu-Ade(TDO)、SD/-Trp-Leu-ura/x-α-gal/AbA(TDO/x/A)和SD/-Trp-Leu-Ade-His/x-α-gal/AbA(QDO/x/A)平板,觀察其生長情況。
采用同樣的方法檢測 eSRK2×THL1、eSRK28×SCR2、SRKJ×THL1、eSRK28×SCR28在 DDO、DDO/x/A平板上的生長情況;然后將DDO/x/A平板上的藍色克隆劃線至TDO、TDO/x/A或TDO、TDO/x/A和QDO/x/A平板,觀察其生長情況。
2 結果與分析
2.1 class-Ⅰ型的SCR、eSRK和class-Ⅱ型的SCR、eSRK的基因克隆及序列分析
擴增的eSRK2位于第1個外顯子的74~1321 bp之間,編碼了416個氨基酸。應用Vector NTI Advance分析Ⅰ型和Ⅱ型的SRK氨基酸序列(圖1),發現其序列相似性在48.6%~84.2%之間,class-Ⅱ型的 eSRK之間的蛋白序列相似性要高于 class-Ⅰ型。class-Ⅱ型中的S-2、S-5和S-15之間的序列相似性達到93.0%~97.3%;class-Ⅰ型的單倍型S-1、S-7、S-8、S-14、S-11、S-24和S-29之間蛋白質序列比classⅡ型的序列相似性低。由此推斷,蛋白序列的不同是造成自交不親和性的主要原因之一。比對結果還發現,Ⅰ型和Ⅱ型的氨基酸序列在219~330 bp的氨基酸區間和446~460 bp的氨基酸區間呈現較大的差異,class-Ⅰ型的氨基酸存在部分缺失。
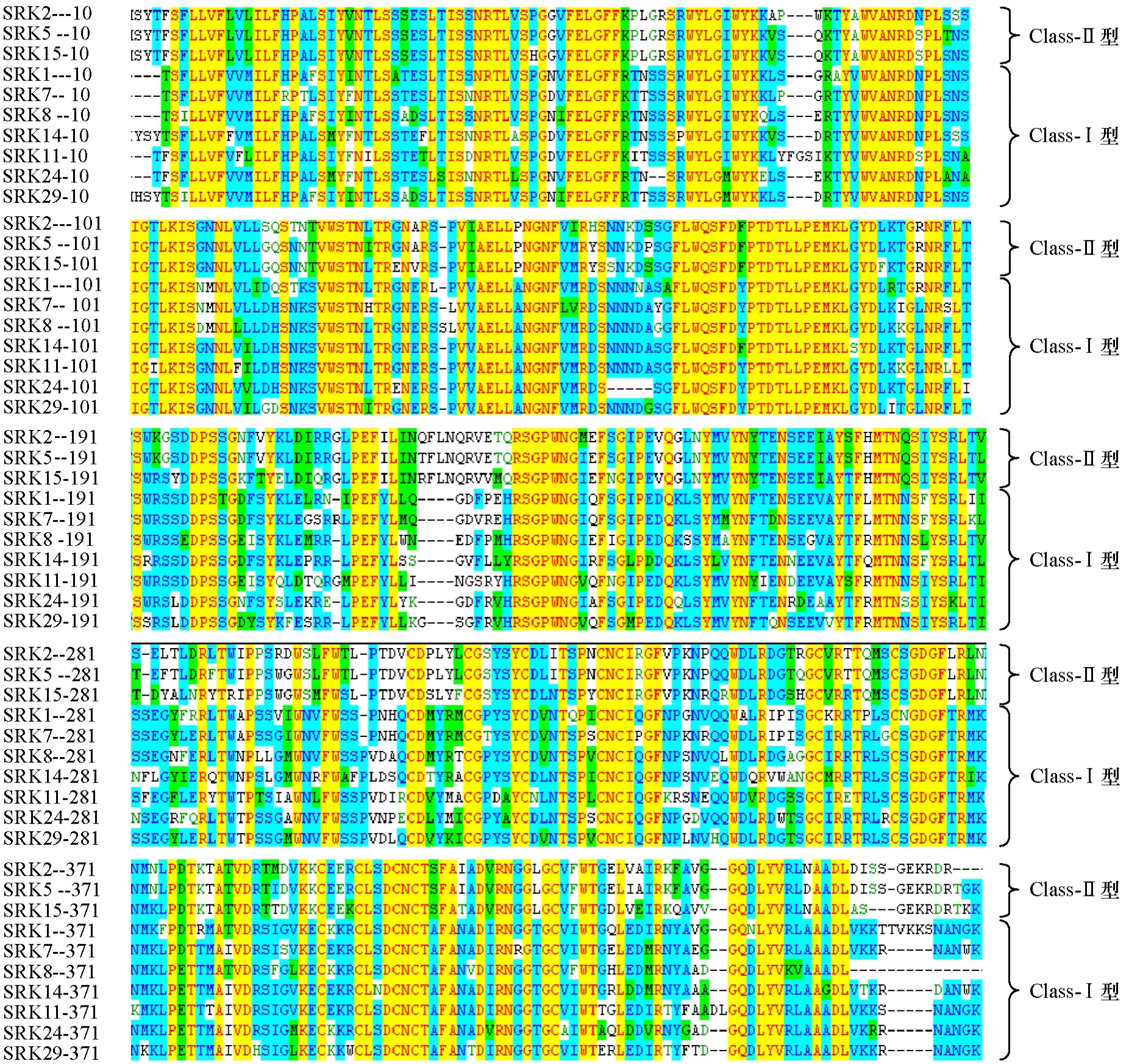
圖1 甘藍class-Ⅰ型和class-Ⅱ型eSRK氨基酸序列比對分析結果
本試驗擴增的是SCR2的第2個外顯子,共編碼65個氨基酸。應用Vector NTI Advance分析SCR氨基酸序列(圖 2)。class-Ⅰ型中不同單倍型 SCR氨基酸的相似度為 20%~76%;class-Ⅱ型中不同單倍型SCR蛋白質序列兩兩之間的相似度達到63%~94%。class-Ⅰ型SCR的4個半胱氨酸殘基高度保守,另外4個半胱氨酸殘基所處的位置略有差異;而class-Ⅱ型SCR的8個半胱氨酸殘基高度保守。Ⅰ型的SCR、eSRK和Ⅱ型的SCR、eSRK在蛋白質序列上存在差異,進而導致了自交不親和蛋白結構和功能的差異,表現為不同的自交不親和強度。
eSRK2、SCR2及THL1的 PCR擴增產物分別約為 1200 bp(圖 3)、200 bp(圖 4)和 400 bp(圖5)。經純化和測序后得出,eSRK2、SCR2和THL1的序列長度分別為195、1248、430 bp,與電泳結果一致。序列分析發現,SCR2與SCR(GB|AB067447|)的序列完全一致;eSRK2與eSRK2D(GB|AJ306585|)的序列完全一致;THL1與劉東等(2003)的序列完全一致。
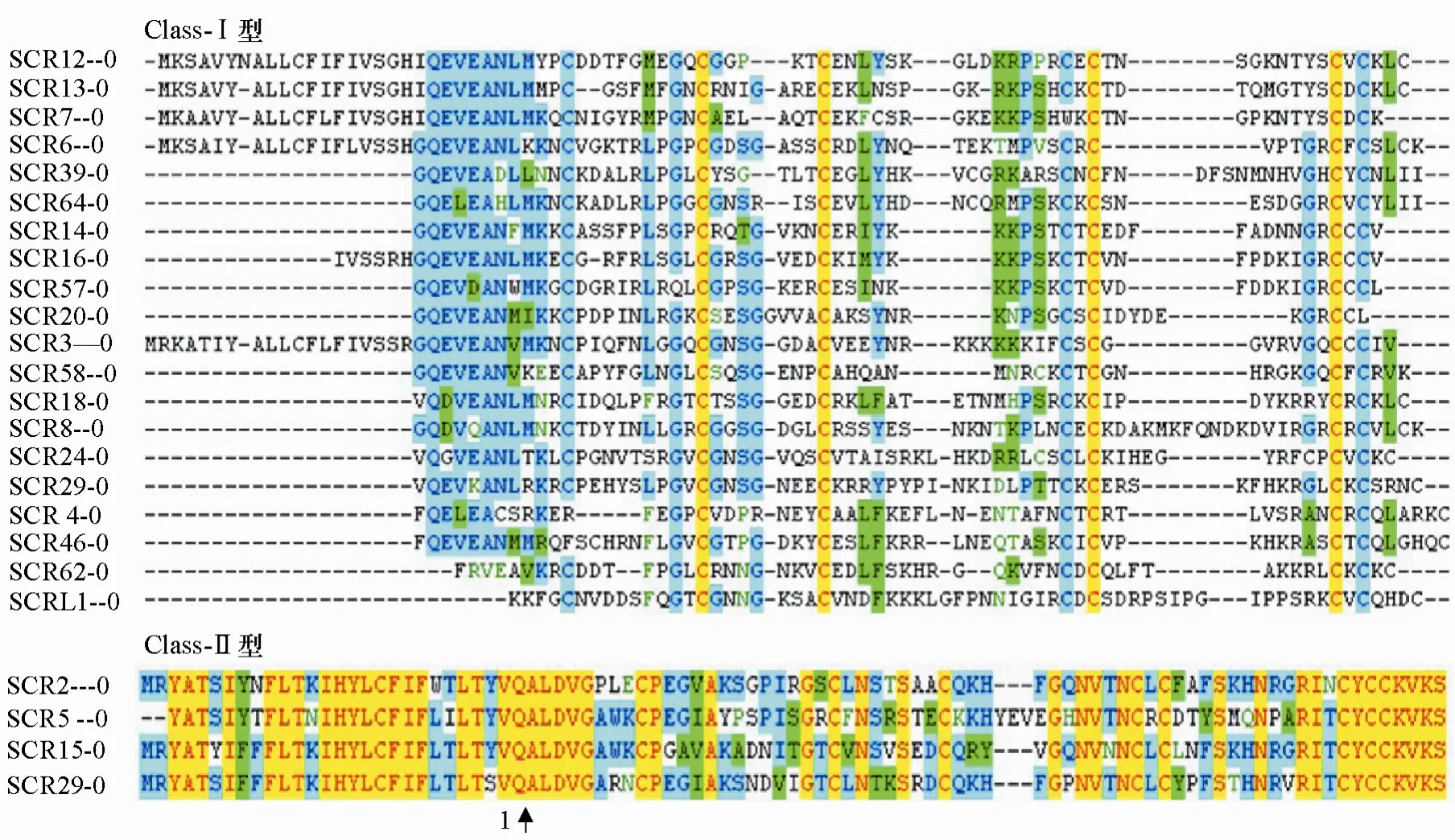
圖2 甘藍class-I型和class-Ⅱ型SCR氨基酸序列比對分析結果
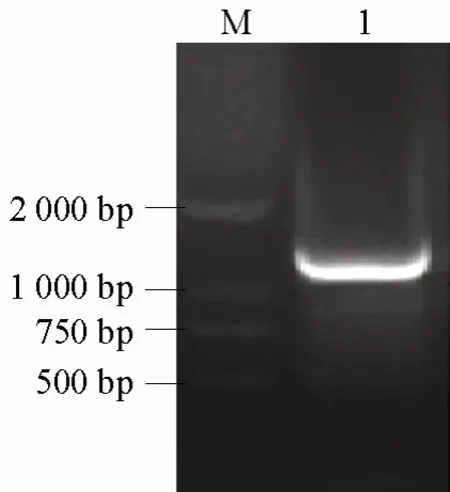
圖3 eSRK2基因PCR擴增結果
2.2 重組質粒 pGADT7-eSRK2、pGBKT7-SCR2、pGADT7-SRKJ 和 pGBKT7-THL1的構建及酶切鑒定
提取篩選的陽性克隆質粒DNA并進行雙酶切鑒定,電泳檢測得到與eSRK2、SCR2、SRKJ、THL1大小一致的片段(圖 6),表明載體構建成功。DNA測序結果分析表明,pGBKT7-eSRK2載體中包含的eSRK2片段序列、插入位點和讀框正確,且插入的eSRK2片段序列與目的序列完全一致。pGBKT7-SCR2載體中包含的SCR2片段序列、插入位點和讀碼框正確,且插入的SCR2片段序列與SCR2(SP11-2)序列完全一致。pGADT7-SRKJ和pGBKT7-THL1載體中包含的SRKJ和THL1片段序列、插入位點和讀碼框正確,且插入的SRKJ和THL1片段序列與SRKJ和THL1序列完全一致。

圖4 SCR2基因PCR擴增結果
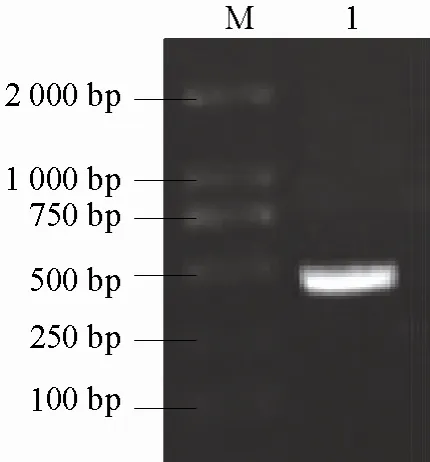
圖5 THL2基因PCR擴增結果M,Trans2K PlusⅡDNA marker;1,THL1。
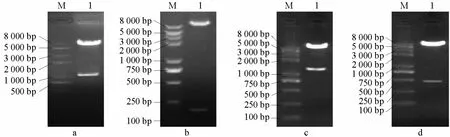
圖6 pGBKT7-eSRK2的EcoRⅠ和SmaⅠ雙酶切,pGBKT7-SCR2、pGADT7-SRKJ和pGBKT7-THL1的EcoRⅠ和BamHⅠ雙酶切圖譜
2.3 誘餌蛋白毒性及其自激活檢測分析和pGBKT7-SRK毒性檢測分析
將 pGBKT7-SCR2質粒、pGBKT7-THL1質粒、空載對照pGBKT7質粒用LiAc法轉化酵母雙雜交報告菌株Y2HGold中,而后接種于SD/-Trp、SD/-Trp/x-a-gal、SD/-Trp/x-α-gal/AbA平板培養基上,30 ℃培養3 d。pGBKT7-SCR2質粒在SD/-Trp(未附圖)和SD/-Trp/x-a-gal(圖7-c-2)平板上產生白色菌落;pGBKT7-THL1質粒在SD/-Trp(未附圖)和SD/-Trp/x-a-gal(圖7-d-2)平板上出現了生長狀態良好、菌斑與空載pGBKT7(圖7-c-1)大小相近、菌斑密度均無明顯差異的白色克隆,證明轉入的表達蛋白對酵母無毒害作用。經酵母菌落 PCR鑒定,證明誘餌載體pGBKT7-SCR2、pGBKT7-THL1已經成功轉化到酵母中。pGBKT7-SCR2和 pGBKT7-THL1在 SD/-Trp/x-α-gal/AbA平板上沒有菌落生長(未附圖)。陽性對照 pGBKT7-p53×pGADT7-T有明顯的藍色克隆子出現,而陰性對照pGBKT7-Lam×pGADT7-T無克隆子出現(圖7-b)。表明含有pGBKT7-SCR2和pGBKT7-THL1質粒的酵母細胞沒有激活報告基因AUR1-C和MEL1,沒有發生自身的轉錄激活作用。
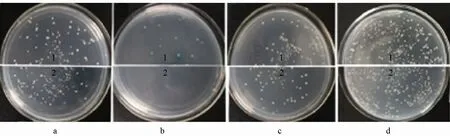
圖7 重組質粒pGBKT7-SCR2自激活作用和毒性檢測及誘餌質粒pGADT7-eSRK2的毒性檢測結果
將pGADT7空載、pGADT7-SRKJ和pGADT7-eSRK2轉化Y187,觀察到它們都能在SD/-Leu平板上良好的生長。pGADT7-SRKJ(圖7-d-1)和pGADT7-eSRK2(圖7-a-2)在SD/-Leu平板上生長狀態良好。而且其對照 pGADT7(圖 7-a-1)斑點大小及生長時間與 pGADT7-eSRK2和pGADT7-SRKJ一致,說明表達蛋白對酵母菌無毒害作用。經酵母菌落 PCR鑒定,證明誘餌載體pGADT7-eSRK2和pGADT7-SRKJ已經成功轉化到酵母中。
2.4 SCR2與 eSRK2、eSRK28與 SCR2、SRKJ 與 THL1相互作用檢測
將 3 個組合 Y187〔pGADT7-SRKJ〕×Y2HGold〔pGBKT7-THL1〕、Y2HGold〔pGBKT7- SCR2〕×Y187〔pGADT7-eSRK2〕、Y2HGold〔pGBKT7-SCR28〕×Y187〔pGADT7-eSRK28〕和 Y2HGold〔pGBKT7-SCR2〕×Y187〔pGADT7-eSRK28〕都轉到DDO平板上,均能出現生長狀態良好的菌斑,表明雙雜交重組表達質粒成功轉入二倍體接合型酵母菌中。同時用 3個組合中片段相應的引物組合對 DDO平板上的菌落進行酵母菌液 PCR檢測,結果發現同一單菌落有明顯的SCR2和eSRK2、SRKJ和THL1目的片段;Y2HGold〔pGBKT7-SCR2〕×Y187〔pGADT7-eSRK28〕未檢測出目的片段,進一步驗證了重組質粒 pGBKT7-SCR2×pGADT7-eSRK2、pGADT7-SRKJ×pGBKT7-THL1成功轉入二倍體接合型酵母菌中。組合 Y187〔pGADT7-SRKJ〕×Y2HGold〔pGBKT7-THL1〕和Y2HGold〔pGBKT7-SCR2〕×Y187〔pGADT7-eSRK2〕都能在DDO/x/A平板上長出明顯的藍色菌落(表2),表明這2個組合激活了報告基因AUR1-C和MEL1;Y2HGold〔pGBKT7-SCR2〕×Y187〔pGADT7-eSRK28〕在DDO/x/A平板上未長出藍色菌落,初步判斷沒有相互作用。為更進一步檢測相互作用的可能性,排除假陽性和假陰性的出現,將 DDO/x/A平板上的藍色克隆劃線至TDO/x/A和QDO/x/A平板上,4 d后發現涂有Y2HGold〔pGBKT7-SCR2〕×Y187〔pGADT7-eSRK2〕的TDO/x/A在平板上呈藍色,而QDO/x/A平板上酵母菌株沒有生長的跡象,表明報告基因HIS3也能被激活,而報告基因ADE未激活(圖8)。Y187〔pGADT7-SRKJ〕×Y2HGold〔pGBKT7-THL1〕和 Y2HGold〔pGBKT7-SCR28〕×Y187〔pGADT7-eSRK28〕在 TDO/x/A和 QDO/x/A平板上都呈藍色,表明報告基因HIS3和ADE被激活。Y2HGold〔pGBKT7-SCR2〕×Y187〔pGADT7-eSRK28〕的融合株在三缺平板和四缺平板上都沒有生長,證明了不同單倍型之間沒有相互作用。
表2 酵母中SCR2與eSRK2、eSRK28與SCR2、SRKJ與THL1相互作用的檢測結果

續 表
與楊紅等(2011)對于class-Ⅰ型的研究結果進行對比分析(表 3),更加系統的說明了自交不親和Ⅰ型和Ⅱ型不同的相互作用關系,即酵母中報告基因表達的數目和類型的差異反映了class-Ⅱ內部的SCR-SRK和class-Ⅰ型的SCR-SRK的互作強度,Ⅰ型高于Ⅱ型;THL與SRK之間存在較高的互作強度。

圖8 酵母融合株在QDO/x/A和TDO/x/A平板上相互作用檢測結果
表3 酵母雙雜交中class-Ⅰ型和class-Ⅱ型不同的相互作用關系
3 結論與討論
酵母的基因組中有一種特殊的上游激活序列(upstream activating sequence,UAS),在轉錄水平調控基因AUR1-C、HIS3、ADE2、lacZ和MEL1表達,它與激活蛋白結合大大增加了下游基因的轉錄速度。GAL4為酵母UAS的結合蛋白,它的正常結合可以激活UAS下游基因的轉錄。GAL4有兩個結構域,即N端的DNA結合結構域(DNA binding domain,BD)和C端的轉錄激活結構域(transcriptional activating domain,AD)。GAL4蛋白的兩個結合域分開時不能激活UAS下游基因的轉錄,只有將這兩部分通過適當的途徑連在一起后才能恢復激活轉錄的活性。如果BD連上1個誘餌蛋白(bait),AD上接1個獵物蛋白(prey),當誘餌蛋白和獵物蛋白發生互作,則BD和AD因距離靠近或結合在一起形成的完整的GAL4蛋白,激活下游報道基因的表達。在酵母菌 Y2HGold中受 3個不同的啟動子作用,通過不同的操縱基因調節 4個完整的報告基因AUR1-C、HIS3、ADE2和MEL1,酵母菌Y187中有2個報告基因MEL1和lacZ;誘餌蛋白和獵物蛋白的互作強度與整個基因之間的相互作用強度存在必然的聯系。根據酵母雙雜交系統可以提出假設:由于誘餌蛋白和獵物蛋白的相互作用強度差異,使下游啟動子對報告基因啟動的數目不同,再由酵母雙雜交試驗得出結果,從而探討class-Ⅰ型和class-Ⅱ型以及與THL1自交不親和程度關系。結果顯示,class-Ⅱ型中能夠激活的報告基因要比 class-Ⅰ型要少,證明了 3個啟動子存在不同的啟動強度,激活不同數目的報告基因。
甘藍中存在自交不親和機制,根據S位點的基因結構不同可以將其分為兩類,class-Ⅰ型和class-Ⅱ型,然而它們產生差異的分子機制尚不清楚。有研究表明,SCR與 SRK胞外結構域之間的親和力很高(Shimosato et al.,2007),SLG僅通過與SRK胞外S結構域聚集而起輔助受體的作用(Cui et al.,2000;Takasaki et al.,2000;Silva et al.,2001)來進一步提高這種親和力,因此SRK與SCR之間的識別速度和結合強度與自交不親和性的強弱類型直接成正相關,當SRK與SCR之間識別快而強時,表現為Ⅰ類自交不親和性的諸多亞類型;當SRK與SCR之間識別慢而弱時,表現為Ⅱ類自交不親和性的諸多亞類型。Bower等(1996)利用酵母雙雜交技術從甘藍柱頭cDNA文庫篩選出SRK下游因子類硫氧還蛋白1/2(THL1/THL2)。Ivanov等(2010)研究證明了SRK是自交不親和信號傳導的核心因子,負責將細胞外信號傳遞到細胞內。Kachroo等(2002)和Ivanov等(2010)提出自交不親和的信號通路是:在甘藍開花時期THL1/THL2與SRK激酶結構域相結合,從而抑制其激酶活性,當花粉落到柱頭上后,相同單倍型SCR與SRK胞外結構域結合,致使 SRK構象變化,使其釋放原本結合在激酶結構域的THL1/THL2,激活SRK激酶域活性,進而在細胞內引發次級級聯反應,繼續傳遞來自細胞外的信號,并最終導致自交不親和反應的發生。這樣自交不親和反應中的SCR、THL與SRK之間就存在一個交替互作的關系,也為本試驗利用酵母雙雜交技術探討兩類SRK之間的SCR-SRK-THL的交替相互作用的關系奠定了基礎。
S單倍型之間的S基因的高度多態性是蕓薹屬自交不親和性以多種類型存在的分子原因(Castric & Vekemans,2007;Edh et al.,2008)。Edh等(2009)將 class-Ⅱ型的S單倍型與class-Ⅰ型的S單倍型的SP11、SLG和SRK的核苷酸序列進行比較分析,發現class-Ⅱ型的序列更加保守;對class-Ⅱ型的S-60單倍型的研究發現,class-Ⅱ型的S-60單倍型與class-Ⅰ型截然不同,而且在S-60中SCR、SRK和SLG的位置與class-Ⅰ型的S-910完全不相同。經氨基酸序列比較分析,發現 class-Ⅱ型的氨基酸多樣化程度明顯要比 class-Ⅰ型小得多。class-Ⅰ型和class-Ⅱ型的SCR、SLG和SRK序列上的差異導致了class-Ⅰ型和class-Ⅱ型的不同,這些不同有助于有效地抑制兩者和不同的S單倍型之間的重組。Kristina等(2009)分析S結構域的進化起源時,根據甘藍的優勢地位,利用系統進化樹分析SRK激酶結構域得到一個緊密的關系,并將其分成了class-Ⅰa型、class-Ⅰb型和class-Ⅱ型,這個結論重新定義了class-Ⅰ型和class-Ⅱ型的關系。在關于class-Ⅰ型的研究中class-Ⅰb型的單體型占有的主導優勢要低一些,它處在class-Ⅰa單體型和class-Ⅱ單體型之間。經過不斷的試驗和總結,Billiard等(2007)定義了一個優勢等級,提出這樣一個理論模型:認為在柱頭共顯性和在花粉中的優勢序位是class-Ⅰa>class-Ⅰb>class-Ⅱ。自交不親和越強就意味著它能夠更有力的阻止自身的花粉與柱頭受精、預防近親繁殖和促進遠系雜交,保持遺傳多樣性,筆者利用酵母雙雜交試驗證明了兩類相互作用的程度不同,驗證了上面提出的理論優勢序位,證實了相互作用的程度class-Ⅰ型>class-Ⅱ型。
SRK作為柱頭S基因,與SCR識別的胞外S結構域編碼區變異性較大(Takayama et al.,2001),兩者之間的這種較大的變異性,在進化上賦予了S單倍型的多態性;而負責同下游功能蛋白作用的C-端激酶結構域的編碼區變異性卻較小,說明在進化上賦予了其下游功能蛋白所決定的是一個目前尚未完全知曉的共同信號傳導過程(Samuel et al.,2008)。張賀翠等(2011)擴增了不同亞種的ARC1基因,利用酵母雙雜交證明都能與SRK激酶域發生互作,也證明了SRK的C-端激酶結構域是一個高度保守的區域。本試驗中利用酵母雙雜交技術證明了組合SRKJ與THL1在四缺平板(QDO/x/A)上能夠生長,存在相互作用,并激活了全部的 4個報告基因,由于C-端激酶結構域的編碼區變異性較小甚至不存在什么變化,所以SRK2激酶域也能夠和THL1相互作用,激活了全部的4個報告基因。另外,Luo等(2011)以甘藍材料D3、B3、E1、A1、H1、230、240、243、N1、G1的柱頭和花粉cDNA為模板,擴增了甘藍的SRK胞外域序列(eSRK3),利用酵母雙雜交技術證明了D3材料中的eSRK與SCR蛋白之間以及E1材料中的eSRK與SCR蛋白之間能夠相互結合,但E1材料中的eSRK與D3材料中的SCR蛋白之間以及D3材料中的eSRK與E1材料中的SCR蛋白之間不相互結合,為深入研究eSRK-SCR作用位點建立了一個技術平臺,也證明了class-Ⅰ型S-3中的SRK和SCR存在相互作用。楊紅等(2011)和本試驗利用酵母雙雜交技術證明了S-28中的SCR與eSRK存在相互作用,且其相互作用區域位于SCR28(GenBank AB190356)的 1876~2052 bp、eSRK28(GenBank AB190356)的 418~1276 bp 區域。同時,也指出了SRK跨膜域的存在與否對SCR與SRK相互作用沒有影響;SCR-SRK復合體的S單倍型特異性導致了SI反應的特異性。這充分說明利用酵母雙雜交技術可以證明class-Ⅰ型自交不親和復等位基因的SCR和eSRK存在相互作用,而class-Ⅱ型S等位基因是否也存在同樣的相互作用,以及class-Ⅰ型和class-Ⅱ型的相互作用程度關系,卻并不了解。
本試驗選擇了一個新的思路,將SCR-SRK-THL之間存在的相互作用做了一個縱向的研究,以試驗中出現的激活不同報告基因的個數來提出假設,以試驗結果推斷 class-Ⅰ型和 class-Ⅱ型的相互作用程度關系,利用酵母雙雜交技術初步確定 PGBKT7-SCR2×pGADT7-eSRK2之間存在相互作用,說明class-Ⅱ型自交不親和復等位基因激活了編碼AUR1-C、MEL1和HIS3報告基因,卻未激活編碼ADE2報告基因。由于激酶域的高度相似性,認為class-Ⅱ型SRK2激酶域與THL1之間的相互作用也可以激活 4個報告基因。對照 pGBKT7-SCR2×pGADT7-eSRK28的融合株在TDO/x/A平板上不能生長,說明不同單倍型中的SCR與eSRK并不存在相互作用。再結合楊紅等(2011)對于class-I型的相互作用的研究結果,以class-Ⅰ型和class-Ⅱ型的相互作用激活3個起始信號傳導元件,而它們之間的識別程度差異表現為不同數量、種類的報告基因,使其在不同的氨基酸缺陷平板上生長,分析 class-Ⅰ型和 class-Ⅱ型與 THL1的相互作用關系。結果表明Ⅰ型的SCR-eSRK激活了4個報告基因;Ⅱ型的SCR-eSRK由于啟動子的啟動強度較Ⅰ型弱,激活了3個報告基因;由于S位點激酶域的高度保守性,兩類SRK(激酶域)的變異性較小,所以SRK(激酶域)-THL1都能激活4個報告基因,利用酵母雙雜交試驗檢測了甘藍SCR、THL和SRK之間交替相互作用的關系。這一結果也證明了酵母雙雜交系統對于class-Ⅱ型自交不親和復等位基因的相互作用檢測同樣適用,這樣也從一個側面進一步完善了酵母雙雜交系統。
class-Ⅰ型的SCR-eSRK和SRK激酶域與THL1之間的相互作用激活了4個報告基因,即編碼AUR1-C、MEL1、ADE2、HIS3的基因,class-Ⅱ型的SCR-eSRK之間的相互作用由于誘餌載體與獵物載體蛋白的結合能力不同,使得激活3個啟動子的能力不同,激活了3個報告基因,即編碼AUR1-C、MEL1、HIS3的基因,ADE2的基因未激活。class-Ⅱ型 SRK2激酶域與 THL1之間的相互作用也可以激活4個報告基因。class-Ⅰ型之間和class-Ⅱ型之間不存在相互作用。初步認為,class-Ⅰ型的自交不親和性強于class-Ⅱ型;利用酵母雙雜交技術同樣可以鑒定class-Ⅱ型之間存在的相互作用,Ⅰ型強于Ⅱ型,甘藍 SCR-SRK-THL存在一個 SCR-SRK、SRK-THL交替相互作用的關系。
劉東,朱利泉,王小佳.2003.甘藍自交不親和信號傳導中SRK結合蛋白基因THL1的克隆與序列分析.園藝學報,30(1):56-58.
張賀翠,楊昆,朱利泉,楊永軍,薛麗琰,楊紅,常登龍,高啟國,任雪松,李成瓊,王小佳.2011.利用酵母雙雜交系統鑒定羽衣甘藍臂重復蛋白(ARC1)與結球甘藍S位點受體激酶(SRK)激酶結構域的相互作用.農業生物技術學報,19(6):988-995.
楊紅,朱利泉,張賀翠,楊永軍,薛麗琰,楊昆,余浩,彭一波,羅兵,吳志剛,黃丹,高啟國,王小佳.2011.利用酵母雙雜交系統鑒定甘藍SCR和SRK胞外域片段間的相互作用.中國農業科學,44(9):1953-1962.
Billiard S,Castric V,Vekemans X.2007.A general model to explore complex dominance patterns in plant sporophytic self-incompatibility systems.Genetics,175:1351-1369.
Bower M S,Matias D D,Fernandes-Carvalho E,Mazzurco M,Gu T,Rothstein S J,Goring D R.1996.Two members of the thioredoxin-h family interact with the kinase domain of a BrassicaSlocus receptor kinase.Plant Cell,8:1641-1650.
Cabrillaca D,Delormea V,Garinb J,Ruffio-Chablec V,Girantona J L,Dumasa C,Gaudea T,Cock J M.1999.The S15self-incompatibility haplotype inBrassica oleraceaincludes threeSgene family members expressed in stigmas.American Society of Plant Physiologists,11:971-986.
Castric V,Vekemans X.2007.Evolution under strong balancing selection:how many codons determine specificity at the female self-incompatibility geneSRKin Brassicaceae.BMC Evolutionary Biology,7:132-136.
Chen C H,Nasrallah J B.1990.A new class of S sequences defined by a pollen recessive self-incompatibility allele ofBrassica oleracea.Molecular and General Genetics MGG,11(2):241-248.
Cui Y H,Bi Y M,Brugiere N,Arnoldo M,Rothstein S J.2000.TheSlocus glycoprotein and theSreceptor kinase are sufficient for self-pollen rejection inBrassica.Proceedings of the National Academy of Sciences of the USA,97:3713-3717.
Edh K,Widén B,Ceplitis A.2008.Unequal segregation ofSRKalleles at theSlocus inBrassica cretica.Genetics Research,90(3):223-228.
Edh K,Widén B,Ceplitis A.2009.The evolution and diversification ofS-locus haplotypes in the Brassicaceae family.Genetics,181:977-984.
Hatakeyama K,Watanabe M,Takasaki T,Ojima K,Hinata K.1998a.Dominance relationships betweenS-alleles in self-incompatibleBrassica campestrisL.Heredity,80:241-247.
Hatakeyama K,Takasaki T,Watanabe M,Hinata K.1998b.Molecular characterization ofSlocus genes,SLGandSRK,in a pollen recessive self-incompatibility haplotype ofBrassica rapaL.Genetics,149:1587-1597.
Kachroo A,Nasrallah M E,Nasrallah J B.2002.Self-incompatibility in the Brassicaceae:receptor-ligand signaling and cell-to-cell communication.Plant Cell,14:227-238.
Kristina E,Bjorn W,Alf C.2009.Molecular population genetics of theSRKandSCRself-Incompatibility genes in the wild plant speciesBrassica cretica.Genetics,181(3):985-995.
Kusaba M,Nishio T,Satta Y,Hinata K,Ockendon D.1997.Striking sequence similarity in interation and intra-specific comparisons of classⅠSLGalleles fromBrassica oleraceaandBrassica campestris:implications for the evolution and recognition mechanism.Plant Biology,94:7673-7678.
Kusaba M,Nishio T.1999.Comparative analysis of S-haplotypes with very similarSLGalleles inBrassica RapaandBrassica oleracea.The Plant Journal,17:83-91.
Ivanov R,Fobis L,Gaude T.2010.When no means no:guide to Brassicaceae self-incompatibility.Trends in Plant Science,15:387-394.
Luo Bing,Xue Li-Yan,Zhu Li-Quan,Zhang He-Cui,Peng Yi-Bo,Chen Song,Yang Hong,Yang Kun,Li Cheng-Qiong,Wang Xiao-Jia.2011.Detection of Interactions between SCR and SRK inBrassica oleraceaL.by Yeast Two-Hybrid System.Acta Agronomica Sinica,37(4):579-586.
Nasrallah J B,Nishio T,Nasrallah M E.1991.The self-incompatibility genes of Brassica:expression and use in genetic ablation of floralt Tissues.Annu Rev Plant Physiol Plant Mol Biol,42:393-422.
Nishio T,Kusaba M,Watanabe M,Hinata K.1996.Registration ofSalleles inBrassica campestrisL.by the restriction fragment size ofSLGs.Theoretical and Applied Genetics,92:388-394.
Samuel M A,Mudgil Y W,Salt J N,Delmas F,Ramachandran S,Chilelli A,Goring D R.2008.Interactions between theS-domain receptor kinases and AtPUB-ARM E3 ubiquitin ligases suggest a conserved signaling pathway in Arabidopsis.Plant Physiology,147:2084-2095.Scutt C P,Croy R R D.1992.An S5 self-incompatibility allele-specific cDNA sequence fromBrassica oleraceashows high homology to the SLR2 gene.Molecular Genetics and Genomics,232:240-246.
Shiba H,Iwano M,Entani T,Ishimoto K,Shimamoto H,Che F-S,Satta Y,Ito A,Takada Y,Watanabe M,Isogai A,Takayama S.2002.The dominance of alleles controlling self-incompatibility inBrassicapollen is regulated at the RNA level.Plant Cell,14:491-504.
Shimosato H,Yokota N,Shiba H,Iwano M,Entani T,Che F S,Watanabe M,Isogai A,Takayama S.2007.Characterization of the SP11/SCR high-affinity binding site involved in self/nonself recognition inBrassicaself-incompatibility.Plant Cell,19(1):107-117.
Silva N F,Stone S L,Christie L N,Sulaman W,Nazarian K A,Burnett L A,Arnoldo M A,Rothstein S J,Goring D R.2001.Expression of theSreceptor kinase in self-compatibleBrassica napuscv.Westar leads to the allele-specific rejection of self-incompatibleBrassica napuspollen.Molecular Genetics and Genomics,265:552-559.
Stein J C,Howlett B,Boyes D C,Nasrallah M E,Nasrallah J B.1991.Molecular cloning of a putative receptor protein kinase gene encoded at the self-incompatibility locus ofBrassica oleracea.Proceedings of the National Academy of Sciences of the USA,88:8816-8820.
Takayama S,Himosato H,Shiba H,Funato M,Che F,Watanabe M,Iwano M,Isogai A,2001.Direct ligand-receptor complex interactions controlBrassicaself-incompatibility.Nature.413:534-538.
Tantikanjana T,Nasrallah M E,Stein J C,Chen C H,Nasrallah J B.1993.An alternative transcript of theSlocus glycoprotein gene in a classⅡ pollen-recessive self-incompatibility haplotype ofBrassica oleraceaencodes a membrane-anchored protein.Plant Cell,5:657-666.Takasaki T,Hatakeyama K,Suzuki G,Watanabe M,Isogai A,Hinata K.2000.TheSreceptor kinase determines self-incompatibility in Brassica stigma.Nature,403:913-918.
