不同基因型普通白菜對鉻脅迫的生理響應及鉻吸收差異
2012-08-08 01:23:26呂金印李鷹翔孫惠莉
中國蔬菜 2012年24期
呂金印 齊 君 王 帥 李鷹翔 孫惠莉
(1西北農林科技大學生命科學學院,陜西楊凌 712100;2西北農林科技大學理學院,陜西楊凌712100)
隨著我國工業生產的快速發展,含鉻(Cr3+)廢水廢渣的排出逐年增加,導致土壤、水體和生物受到不同程度的污染。作物受Cr3+污染后,生長發育滯后,產量降低,甚至死亡(Bini et al.,2008;王愛云 等,2011)。Cr3+進入食物鏈,對人體健康造成危害。因此,篩選Cr3+低積累作物品種顯得較為重要和迫切。普通白菜〔Brassica campestrisL.ssp.chinensis(L.)Makino var.communisTsen et Lee〕是我國廣泛種植的大宗葉菜之一,對土壤環境中Zn、Cu、Pb有一定的富集能力,且品種間存在顯著差異(任安芝和高玉葆,2000,胡學玉 等,2002;于光金 等2009)。鉻對綠豆、黃瓜和油菜生長的影響已有報道(郭峰,2006;陳慶華,2009;沈奇 等,2009),但鉻處理下普通白菜對重金屬的吸收及生理變化鮮有報道。本試驗選用 6個普通白菜品種,測定鉻處理下不同基因型普通白菜對鉻的吸收積累及生理變化,探討普通白菜在鉻處理下的生理響應,為普通白菜品種選育及無害化栽培提供依據。
1 材料與方法
1.1 試驗材料
供試普通白菜品種為矮抗青、綠秀、五月慢、七寶青、雞毛菜和黃心菜。其中矮抗青、五月慢、雞毛菜、黃心菜種子購自陜西楊凌西北農林科技大學農城種業科技中心,綠秀種子購自青島國際種苗有限公司,七寶青種子購自南京蘇豪種業有限公司。
1.2 試驗方法
試驗于2011年4月在西北農林科技大學生命科學學院網室內進行。取大田耕層土壤,經室溫風干后過5 mm篩。土壤有機質含量1.14%,速效氮85.94 mg·kg-1,速效磷40.01 mg·kg-1,速效鉀81.34 mg·kg-1,總鉻28.03 mg·kg-1,pH=8.3。以溶液態施入分析純CrCl3,制成全鉻含量為 90、150、200、250 mg·kg-1的污染土壤,其中最高濃度是參照國家土壤環境質量標準(GB15618-1995)中的二級標準。以不施鉻為對照。室溫放置20 d后裝盆(上口徑20.6 cm,下口徑10.4 cm,高13.9 cm),每盆裝2.3 kg干土。將6個普通白菜品種分別播于盆中,每盆15粒,20 d后間苗至8株。每品種3次重復。培養期間,根據普通白菜生長需要正常水肥管理。生長40 d后收集全部植株,先用自來水沖凈表面污物,將地上部和地下部分開,用于測定各項指標。
1.3 測定項目
1.3.1 土壤基礎養分測定 栽種前采用棋盤法收集土壤樣品,土壤 pH測定采用電位法(水∶土=2.5∶1);有機質含量測定采用重鉻酸鉀滴定法;速效氮含量測定采用0.01 mol·L-1NaCl浸提、酚二磺酸比色法;速效磷含量測定采用0.5 mol·L-1NaHCO3浸提、鉬銻抗比色法;速效鉀含量測定采用1.0 mol·L-1NH4OAc浸提、火焰光度法;總鉻含量測定采用HNO3-HClO4聯合消化、原子吸收分光光度計(柳鈴 等,2010)。
1.3.2 生物量測定 分別取地上部和地下部植株,105 ℃殺青 10 min,80 ℃烘干至恒質量,測定地上部和地下部干質量。
1.3.3 鉻含量測定 分別取地上部和地下部植株,自來水浸泡10 h,除去根系表面Cr3+。離子水沖洗,吸干水分,105 ℃殺青15 min,80 ℃烘干至恒質量,磨碎混勻,加入 HNO3-HCLO4(4V∶1V)混合酸,220 ℃沙浴消解至透明色,用火焰原子吸收分光光度計(Perkin Elmer AA180-80)測定鉻含量(孫守琴 等,2009;Liu et al.,2009)。
1.3.4 葉綠素、丙二醛、可溶性蛋白含量測定 用剪刀采集葉片鮮樣,每個樣品 0.5 g。葉綠素含量測定采用80%丙酮浸提法;丙二醛含量測定采用TBA比色法;可溶性蛋白含量測定采用考馬斯亮藍G-250法(高俊鳳,2006)。
1.3.5 過氧化氫酶(CAT)、超氧化物歧化酶(SOD)、過氧化物酶(POD)活性測定 酶液提取:分別稱取葉片0.5 g,加入磷酸緩沖液(pH=7.5),冰浴研磨,4 ℃下離心(4000 r·min-1)15 min,上清液即為粗酶提取液。CAT活性采用紫外吸收法測定;SOD活性采用氮藍四唑(NBT)比色法測定;POD活性采用愈創木酚比色法測定(高俊鳳,2006;孫群和胡景江,2006)。
各項指標測定均為3次重復,結果以平均值±標準差表示,采用SPSS 17.0軟件進行統計分析。
2 結果與分析
2.1 鉻處理對不同基因型普通白菜生物量的影響
由表1可知,隨著鉻濃度增加,6個普通白菜品種生物量均呈下降趨勢。低濃度Cr3+(90 mg·kg-1)處理下,6個普通白菜品種的生物量與對照相比下降均不顯著。高濃度Cr3+(250 mg·kg-1)處理下,黃心菜、矮抗青、五月慢、綠秀及七寶青的生物量與對照相比顯著下降,其中黃心菜、七寶青下降尤為明顯,分別比對照下降了58.0%和48.8%;雞毛菜下降幅度較小,比對照下降了34.0%。說明鉻脅迫會降低普通白菜的生物量,對雞毛菜的影響較小,而黃心菜、矮抗青、五月慢、綠秀及七寶青的生物量均顯著降低。

表1 不同濃度鉻處理對不同基因型普通白菜生物量的影響
2.2 鉻處理對不同基因型普通白菜鉻吸收量的影響
由表2可知,低濃度Cr3+(90 mg·kg-1)處理下,矮抗青、雞毛菜、五月慢、七寶青地上部鉻含量與對照相比下降,而黃心菜和綠秀略有增加;6個普通白菜品種地下部鉻含量均顯著增加。高濃度Cr3+(250 mg·kg-1)處理下,6個普通白菜品種地上部和地下部鉻含量大體上均顯著增加,其中地下部鉻含量增幅較大。說明高濃度Cr3+處理對普通白菜的根部傷害較大。
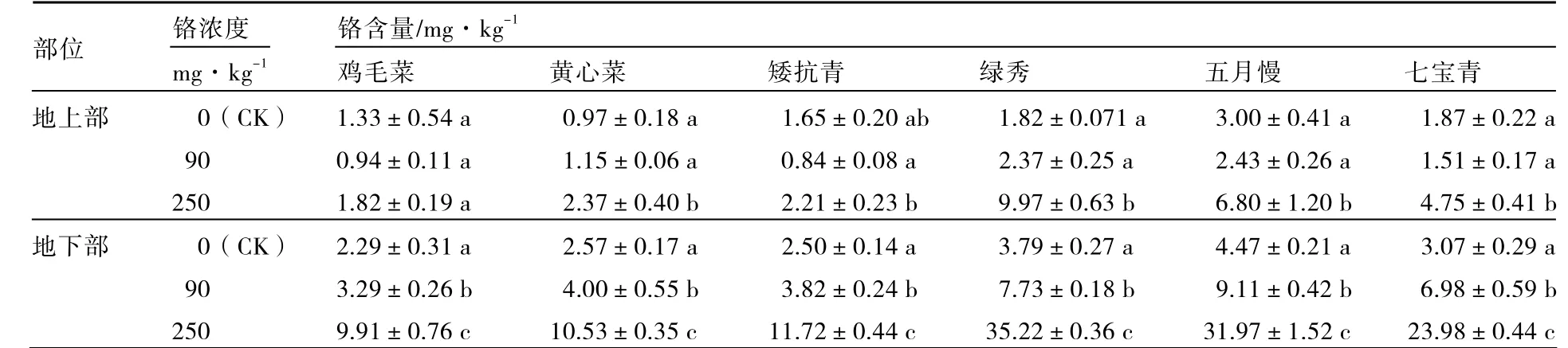
表2 不同濃度鉻處理對不同基因型普通白菜地上部、地下部鉻含量的影響
由圖1可以看出,6個基因型普通白菜的鉻轉移率均隨著鉻濃度的增加而降低。高濃度Cr3+(250 mg·kg-1)處理下,雞毛菜和矮抗青的鉻轉移率分別為0.18和0.19,低于綠秀(0.28)。表明在鉻處理下,雞毛菜和矮抗青根部滯留鉻較多,向地上部轉運較少;而綠秀向地上部轉運相對較多。
2.3 鉻處理對不同基因型普通白菜葉綠素含量的影響
由圖 2可以看出,隨著鉻濃度的增加,普通白菜葉片葉綠素含量呈降低趨勢。低濃度 Cr3+(90 mg·kg-1)處理下,雞毛菜、五月慢和綠秀的葉綠素含量與對照相比增加。高濃度Cr3+(250 mg·kg-1)處理下,黃心菜和七寶青的葉綠素含量分別比對照下降了 38.5%和41.9%。這可能是鉻處理下光合作用減弱,葉綠體結構和功能受到影響,導致葉綠素分解。
2.4 鉻處理對不同基因型普通白菜丙二醛含量的影響
丙二醛可作為植物在逆境下細胞膜脂過氧化程度的指標。由圖3可知,隨著鉻處理濃度的增加,6個基因型普通白菜葉片丙二醛含量呈增加趨勢。高濃度 Cr3+(250 mg·kg-1)處理下,雞毛菜、黃心菜、矮抗青的丙二醛含量分別是對照的1.86、1.59、1.51倍,而五月慢、綠秀、七寶青分別是對照的2.15、2.25、2.58倍。表明雞毛菜、黃心菜、矮抗青的耐鉻脅迫能力較強。
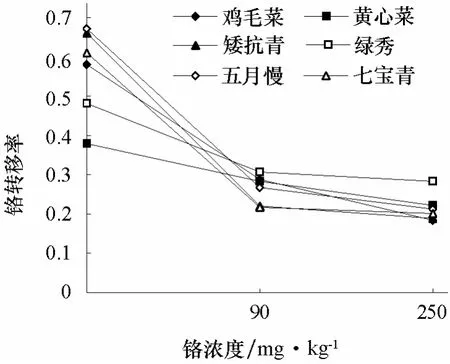
圖1 不同濃度鉻處理對不同基因型普通白菜鉻轉移率的影響
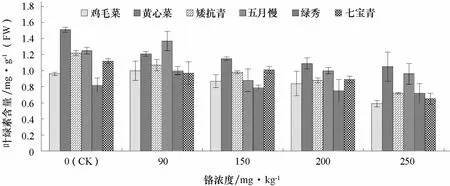
圖2 不同濃度鉻處理對不同基因型普通白菜葉片葉綠素含量的影響
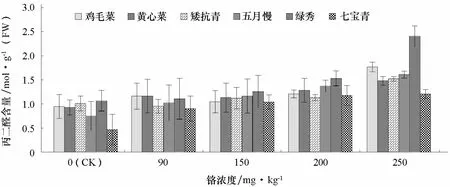
圖3 不同濃度鉻處理對不同基因型普通白菜葉片丙二醛含量的影響
2.5 鉻處理對不同基因型普通白菜可溶性蛋白含量的影響
隨著鉻濃度的增加,普通白菜葉片可溶性蛋白含量呈先增后降的趨勢(圖 4)。低濃度 Cr3+(90 mg·kg-1)處理下,葉片可溶性蛋白含量增加,而高濃度(250 mg·kg-1)Cr3+處理則下降。中濃度Cr3+(150 mg·kg-1)處理下,雞毛菜和黃心菜的可溶性蛋白含量下降明顯,分別比對照降低了20.5%和36.3%。高濃度Cr3+(250 mg·kg-1)處理下,黃心菜和七寶青的可溶性蛋白含量降幅明顯,分別比對照下降了28.6%和25.5%,而矮抗青和雞毛菜分別比對照下降了16.9%和23.0%。
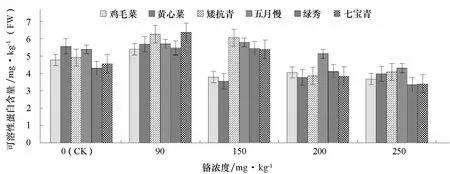
圖4 不同濃度鉻處理對不同基因型普通白菜葉片可溶性蛋白含量的影響
2.6 鉻處理對不同基因型普通白菜抗氧化酶活性的影響
SOD、CAT、POD是植物體內清除活性氧重要的細胞保護酶類,其活性高低可以反映植物對逆境脅迫的適應能力(鄒清成 等,2011)。由圖5可以看出,6個基因型普通白菜葉片POD、SOD、CAT活性隨著鉻濃度的增加呈先升后降的趨勢。高濃度Cr3+(250 mg·kg-1)處理下,綠秀、七寶青、五月慢POD活性分別比對照增加了 17.4%、9.2%、18.9%,SOD活性分別比對照增加了14.4%、12.6%、0.69%;而雞毛菜、黃心菜、矮抗青POD活性分別比對照增加了31.7%、29.3%、33.2%,SOD活性分別比對照增加了28.1%、18.2%、24.1%。中濃度Cr3+(200 mg·kg-1)處理下,雞毛菜、黃心菜、矮抗青、五月慢的CAT活性達到最高值,而綠秀、七寶青在150 mg·kg-1處理時達到最高值并開始下降。說明鉻脅迫下雞毛菜、黃心菜、矮抗青具有相對較高抗氧化酶活性,清除活性氧能力較強。
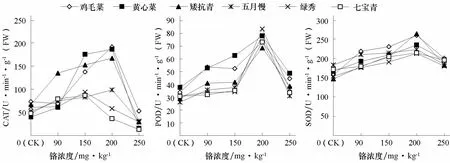
圖5 不同濃度鉻處理對不同基因型普通白菜葉片CAT、POD、SOD活性的影響
3 結論與討論
鉻對植物生長發育產生影響(Han et al.,2004),導致植物生理代謝紊亂,抑制正常生長。植物莖葉干物質可作為篩選對重金屬耐性的指標之一。本試驗中,低濃度鉻處理普通白菜的生物量與對照相比沒有顯著影響,但高濃度的鉻會明顯降低普通白菜的生物量。這與王愛云等(2011)對芥菜型油菜在鉻脅迫下的研究結果相符。可能高濃度鉻抑制植物根細胞分化,阻礙水分吸收,從而導致植株矮小,葉片泛黃,葉面積減少,生物量降低(Dixit et al.,2002;Suseela et al.,2002;Shanker et al.,2004)。隨著鉻濃度的增加,黃心菜和七寶青生物量下降顯著,而雞毛菜、矮抗青則下降較少,可以推測雞毛菜和矮抗青具有較強的鉻耐受力。
轉移率是反映植株富集重金屬能力的指標之一,轉移率低表明植株地上部分的富集量少。本試驗中,雞毛菜和矮抗青在高濃度鉻脅迫下的鉻轉移率低于其他基因型普通白菜,表明雞毛菜和矮抗青的地上部鉻積累量相對較少。
丙二醛作為膜脂過氧化的產物(周希琴和吉前華,2005),其在機體內的積累會對細胞產生毒害作用,可作為膜脂過氧化程度的指標之一。隨著鉻濃度的增加,6個基因型普通白菜葉片丙二醛含量都逐漸增加。鉻脅迫可促進植株體內活性氧的產生,啟動膜脂過氧化,導致丙二醛積累,引起活性氧代謝平衡失調而最終破壞生物膜結構。本試驗中,高濃度Cr3+(250 mg·kg-1)處理下,雞毛菜、黃心菜和矮抗青的丙二醛含量增幅小于五月慢、綠秀和七寶青。表明雞毛菜、黃心菜和矮抗青具有相對較高的抗氧化能力。
一般認為,重金屬脅迫影響植物體內蛋白質代謝(柳玲 等,2010)。本試驗中,高濃度Cr3+處理下,黃心菜和七寶青可溶性蛋白含量下降幅度大,而矮抗青和雞毛菜可溶性蛋白含量下降幅度則相對較小,表明鉻脅迫對矮抗青和雞毛菜的影響相對較小。
葉綠素含量的高低在一定程度上反映光合作用水平。高濃度鉻處理下,黃心菜和七寶青的葉綠素含量下降明顯,可能是重金屬抑制葉綠體色素生物合成及光合電子傳遞,導致葉綠素含量減少(何冰 等,2009)。
植物對逆境的生理響應與體內的抗氧化防御系統密切相關(陳茂銓和劉術新,2011)。有研究指出,鎘脅迫下不同基因型的番茄幼苗抗氧化酶的生理活性存在差異(張微 等,2010)。黃永杰等(2009)通過水培試驗研究不同濃度銅脅迫下水花生生長特性和體內生理特性的變化規律,結果表明低濃度銅處理下植物細胞內的SOD、POD和CAT活性均有所提高,高濃度銅處理下則有所下降。本試驗中,隨著Cr3+濃度的增加,6個基因型普通白菜POD、SOD、CAT活性均先升后降。低濃度 Cr3+處理下,SOD活性呈上升趨勢,可能是由于鉻刺激了植物體本身存在的SOD或者誘導合成新的SOD,而POD為清除過多的H2O2與SOD起協同保護作用(Vannini et al.,2006),以延緩活性氧的積累和降低鉻脅迫造成的傷害。POD活性升高可能是鉻脅迫導致 H2O2等POD底物增加。高濃度Cr3+(250 mg·kg-1)處理下,綠秀、七寶青、五月慢POD和SOD 活性增幅低于雞毛菜、黃心菜和矮抗青,表明雞毛菜、黃心菜、矮抗青脂質過氧化程度低,具有相對較高的抗氧化能力。
蔬菜對重金屬的吸收積累存在基因型差異的普遍性,有選擇的種植重金屬低積累品種,對蔬菜無害化栽培具有重要的意義。本試驗選用的 6個普通白菜品種中,雞毛菜、矮抗青在高濃度鉻處理下,生物量下降幅度低于五月慢、七寶青、黃心菜及綠秀;鉻轉移率低,地上部鉻積累相對較少;丙二醛含量相對較低;抗氧化酶 POD、SOD、CAT活性較強,具有較好的抗氧化能力,屬于鉻低積累品種,適宜在生產中推廣。
陳茂銓,劉術新.2011.Spd浸種對Cd脅迫下油菜幼苗的抗氧化系統的影響.農業環境科學學報,30(10):1953-1957.
陳慶華.2009.鉻脅迫對黃瓜種子萌發及某些生理生化指標的影響.湖北農業科學,48(6):1402-1405.
高俊鳳.2006.植物生理實驗指導.北京:高等教育出版社.
郭峰.2006.重金屬Hg、Pb、Cr單一污染對綠豆生理生化指標的影響.山西農業大學學報,26(3):382-387.
何冰,何計興,何新華,李峰,韋泓明,蔡春波.2009.鉛脅迫對楊梅生理特性的影響.農業環境科學學報,28(6):1263-1268.
胡學玉,李學垣,謝振翅.2002.不同青菜品種吸鋅能力差異及與根系分泌物的關系.植物營養與肥料,8(2):234-238.
黃永杰,楊紅飛,楊集輝,王曉蘭,周守標.2009.銅脅迫對水花生生長及活性氧代謝的影響.生態學雜志,28(6):1112-1116.
柳玲,呂金印,張微.2010.不同濃度Cr6+處理下芹菜的鉻累積量及生理特性.核農學報,24(3):639-644.
任安芝,高玉葆.2000.鉛、鎘、鉻單一和復合污染對青菜種子萌發的生物學效應.生態學雜志,19(1):19-22.
沈奇,秦信蓉,張敏琴,郭貴敏,杜才富.2009.鉻脅迫對油菜種子萌發及幼苗生長的毒性效應.貴州農業科學,37(10):25-26.
孫群,胡景江.2006.植物生理研究技術.楊凌:西北農林科技大學出版社:168-170,176-178.
孫守琴,何明,曹同,程頌,宋洪濤.2009.Pb、Ni脅迫對大羽蘚抗氧化酶系統的影響.應用生態學報,20(4):937-942.
王愛云,鐘國鋒,徐剛標,劉志祥,申響保.2011.鉻脅迫對芥菜型油菜生理特性和鉻富集的影響.環境科學,32(6):1717-1725.
于光金,成杰民,姜琪,徐淑新.2009.不同土壤施銅對小白菜生物效應及吸收能力的研究.資源環境與發展,(4):22-25.
張微,呂金印,柳玲.2010.不同基因型番茄幼苗對鎘脅迫的生理響應及鎘吸收差異.農業環境科學學報,29(6):1065-1071.
周希琴,吉前華.2005.鉻脅迫下不同品種玉米種子和幼苗的反應及其與鉻積累的關系.生態學雜志,24(9):1048-1052.
鄒清成,朱開元,劉慧春,周江華,馬廣瑩.2011.外源茉莉酸甲酯對非生物脅迫下蝴蝶蘭幼苗葉綠素熒光和抗氧化指標的影響.植物生理學報,47(9):913-917.
Bini C,Maleci L,Romanin A.2008.The chromium issue in soils of the leather tannery district in Italy.J Geochem Explor,96:194-202.
Dixit V,Pandey V,Shyam R.2002.Chromiumions inactivate electron transport and enhance superoxide generationin vivoin pea(Pisum sativumL. cv. Azad)root mitochondria.Plant,Cell & Environment,25(5):687-693.
Han F X,Maruthi S B B,Monts D L,Su Y.2003.Phytoavailability and toxicity of trivalent and hexavalent chromium toBrassica juncea.New Phytol,162:489-499.
Liu W T,Zhou Q X,Sun Y B,Liu R.2009.Identification of Chinese cabbage genotypes with low cadmium accumulation for food safety.Environ Pollut,157:1961-1967.
Shanker A K,Djanaguiraman M,Sudhagar R,Chandrashekar C N,Pathmanabhan G.2004.Differential antioxidative response of ascorbate glutathione pathway enzymes and metabolites to chromium speciation stress in green gram〔Vigna radiata(L.)R.Wilczek.cv CO 4〕roots.Plant Science,166(4):1035-1043.
Suseela M R,Sinha S,Singh S,Saxena R.2002.Accumulation of chromium and scanning electron microscopic studies inScirpus lacustrisL.treated with metal and tannery effluent.Bulletin of Environmental Contamination and Toxicology,68(4):540-548.
Vannini C,Iriti M,Bracale M,Locatelli F,Faoro F,Croce P,Pirona R,di Maro A,Coraggio I,Genga A.2006.The ectopic expression of the riceOsmyb4gene in Arabidopsis increases tolerance to abiotic,environmental and biotic stresses.Physiological and Molecular Plant Pathology,69(1-3):26-42.
Wu F Y,Leng H M,Wu S C,Ye Z H,Wang M H.2009.Variation in arsenic,lead and zinc tolerance and accumulation in six populations ofPteris vittataL.from China.Enviromental Pollution,157:2394-2404.
