我國G2型人蛔蟲和豬蛔蟲的遺傳多樣性*
2012-08-21 06:52:04周春花彭衛(wèi)東
中國人獸共患病學報 2012年11期
周春花,彭衛(wèi)東
2.南昌大學生命科學與食品工程學院生物技術(shù)系,南昌 330031
人蛔蟲(Ascarislumbricoides)和與其同屬的豬蛔蟲(A.suum,)是具有重要社會經(jīng)濟意義的常見土源性線蟲,其分類學地位一直存有爭議,而近20年來分子分型的研究對這兩種蛔蟲的遺傳結(jié)構(gòu)、流行病學和種群演化等方面提供了豐富的信息[1]。比如,用ITS1對我國蛔蟲的基因分型,發(fā)現(xiàn)共有5種基因型(G1-G5),在人蛔蟲種群以G1型為主(約占70%);而豬蛔蟲種群中則以G3為主(約80%);各基因型的在不同地區(qū)同一宿主內(nèi)的分布頻率基本上相同,而在不同宿主之間的分布明顯不同[2]。線粒體DNA的2個基因片段COX1和NAD1的篩選也得到類似結(jié)果[3]。進一步用來自人體的G1型和來自豬體的G3型蛔蟲分別實驗感染C57BL/6小鼠和豬,結(jié)果表明,不同基因型的蛔蟲具有不同的宿主寄生特異性[4-6]。
G2型是人蛔蟲和豬蛔蟲共有的基因型,其在人蛔蟲種群中占25.5%,而在豬蛔蟲占15.2%[2]。如果說,G1和G3型在各宿主的分別偏好提示了人型蛔蟲和豬型蛔蟲的不同宿主特異性,那么G2型是否也存在某種差異性?有趣的是,初步實驗感染結(jié)果表明,來自人體的G2型蛔蟲不能在豬體內(nèi)發(fā)育為成蟲,而來自豬的G2型蛔蟲卻能夠在豬體內(nèi)發(fā)育為成蟲[6];鑒于ITS1標記不能進一步區(qū)分G2型蛔蟲,故有必要借助其它分子標記對其遺傳變異進行進一步研究。微衛(wèi)星在基因組中數(shù)量多、分布廣、多態(tài)性豐富、呈孟德爾共顯性遺傳等特點而被廣泛應用于動物種群間親緣關(guān)系、群體內(nèi)和群體間遺傳變異等的檢測。近年來使用微衛(wèi)星標記,發(fā)現(xiàn)了人蛔蟲和豬蛔蟲雜交個體的存在[7-8]以及豬蛔蟲一妻多夫現(xiàn)象[9];另外,譜系地理學研究提示人蛔蟲線粒體COX1的H9單倍體可能是豬蛔蟲遠古祖先[10]。這些都表明人蛔蟲和豬蛔蟲之間的關(guān)系并非是僅僅“同種或者異種”那么簡單。尤其是對人和豬共有的占有種群相當比例的同為G2型的蛔蟲,其遺傳變異情況尚不了解。因此,本研究通過20個常染色體微衛(wèi)星座位對中國G2型蛔蟲進行遺傳多樣性研究,以揭示G2型人蛔蟲和G2型豬蛔蟲遺傳學差異及其在流行病學方面的可能啟示。
1 材料與方法
1.1 材料 本研究所用的69個G2型蛔蟲樣本均來自Peng等自我國6省(自治區(qū))采集的標本[2],其中48條來自人體,21條來自豬體內(nèi)。
1.2 主要試劑和儀器 蛔蟲組織DNA提取試劑盒(WizardSVGenomicDNAPurificationSystem)購自Promega公司,熒光引物由上海英駿生物技術(shù)有限公司合成,DNA Taq酶(PremixTaqVersion 2.0)購自寶生物工程(大連)有限公司。熱循環(huán)儀為德國Eppendorf公司產(chǎn)品。STR掃描由上海生工生物技術(shù)有限公司完成。
1.3 方法
1.3.1 基因組DNA的提取 從卡洛氏液中取出蛔蟲樣本,剪下一小段(約5 mm)用濾紙吸去固定液并浸泡于雙蒸水中回水24 h以上,其間更換雙蒸水至少6次,吸干管中的液體,于50℃烘干15 min。參照試劑盒的操作步驟提取基因組DNA。
1.3.2 微衛(wèi)星擴增 采用20對常染色體微衛(wèi)星引物進行擴增。反應體系為20μL,上下游引物各0.2μmol/L,1μL DNA模板,加水至20μL。反應條件為94℃預變性5 min,先進行5個循環(huán)的高溫PCR:94℃變性45 s,退火30 s,72℃延伸1 min,再進行30個循環(huán)的低溫PCR:94℃變性45 s,退火30 s,72℃延伸1 min,最后72℃延伸7 min。引物及退火溫度詳見表1。
1.3.3 等位基因分型 PCR擴增產(chǎn)物先用1.5%瓊脂糖電泳檢測是否有擴增產(chǎn)物,以及擴增產(chǎn)物是否在所需的長度范圍內(nèi)。再送往上海生工生物技術(shù)有限公司進行STR掃描,由該公司利用測序儀進行毛細管電泳,配合Genemapper軟件分析確定微衛(wèi)星片斷長度。
1.3.4 數(shù)據(jù)分析 通過 CERVUS V2.0[11]計算各個位點的等位基因數(shù)、等位基因頻率、觀測雜合度(observed heterozygosities,H O)、期望雜合度(expected heterozygosities,H E)和多態(tài)信息含量(polymorphism information content,PIC)。利用軟件GENETIX V4.05[12]計算人蛔蟲和豬蛔蟲間的遺傳分化程度FST和基因流Nm。應用ARLEQUIN V3.11[13]進行分子變異方差分析(analysis of molecular variance,AMOVA),估測遺傳變異在種群間和種群內(nèi)的分配情況。
2 結(jié) 果
2.1 G2型人蛔蟲和G2型豬蛔蟲的等位基因及頻率差異 G2型人蛔蟲共檢出378個等位基因,范圍在3~31,平均每對引物可檢測18.9個等位基因;G2型豬蛔蟲共檢出229個等位基因,范圍在5~19,平均每對引物可檢測11.45個等位基因。G2型人蛔蟲的等位基因頻率在0~0.821之間;G2型豬蛔蟲的等位基因頻率在0~0.833之間。20個微衛(wèi)星座位共檢測到433個等位基因,變化范圍為5~38,平均每個位點等位基因數(shù)21.65個。在G2型人蛔蟲中位點19的多態(tài)性最豐富,有31個等位基因,在G2型豬蛔蟲中位點18的多態(tài)性最豐富,有19個等位基因;最少的在G2型人蛔蟲中有3個等位基因,在G2型豬蛔蟲中有5個等位基因,均為位點15。從各微衛(wèi)星基因座位的等位基因頻率來看,各個等位基因頻率分布不均勻,每個基因座位都有一種相對優(yōu)勢等位基因存在。20個基因位點中有14個位點在2個蛔蟲種群中的優(yōu)勢等位基因一致,6個基因位點(位點1、6、10、12、16和17)在2個蛔蟲種群中的優(yōu)勢等位基因不一致。部分等位基因在不同蛔蟲種群中所占的比例存在較大的差異。兩個種群在除位點13G2型豬蛔蟲無特有等位基因,位點15G2型人蛔蟲無特有等位基因外,其它位點中均存在各自的特有等位基因,其中G2型人蛔蟲有204個特有等位基因,G2型豬蛔蟲有55個特有等位基因,見表2。
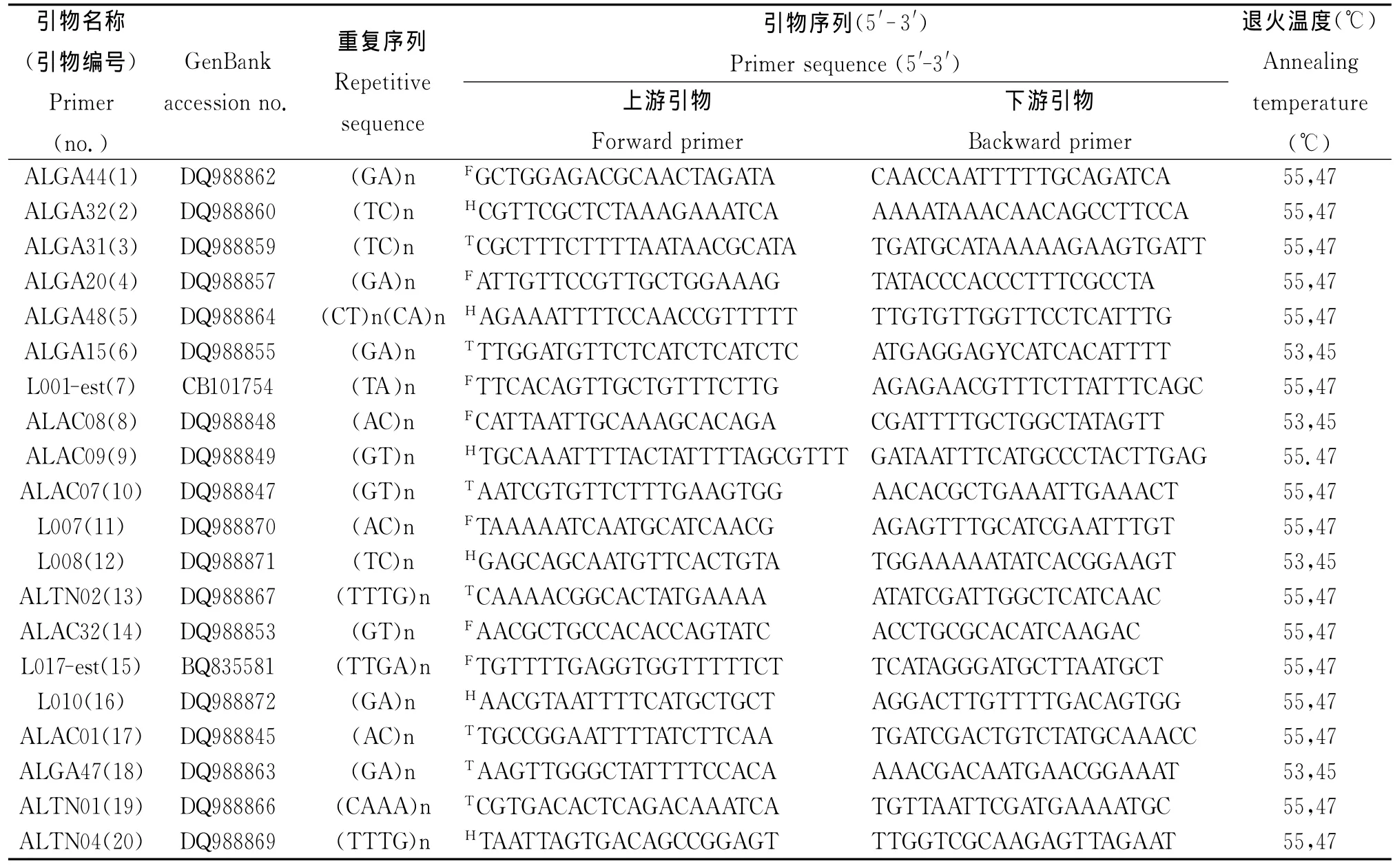
表1 20對微衛(wèi)星引物及退火溫度Tab.1 Twenty pairs of microsatellite primers and annealing temperature
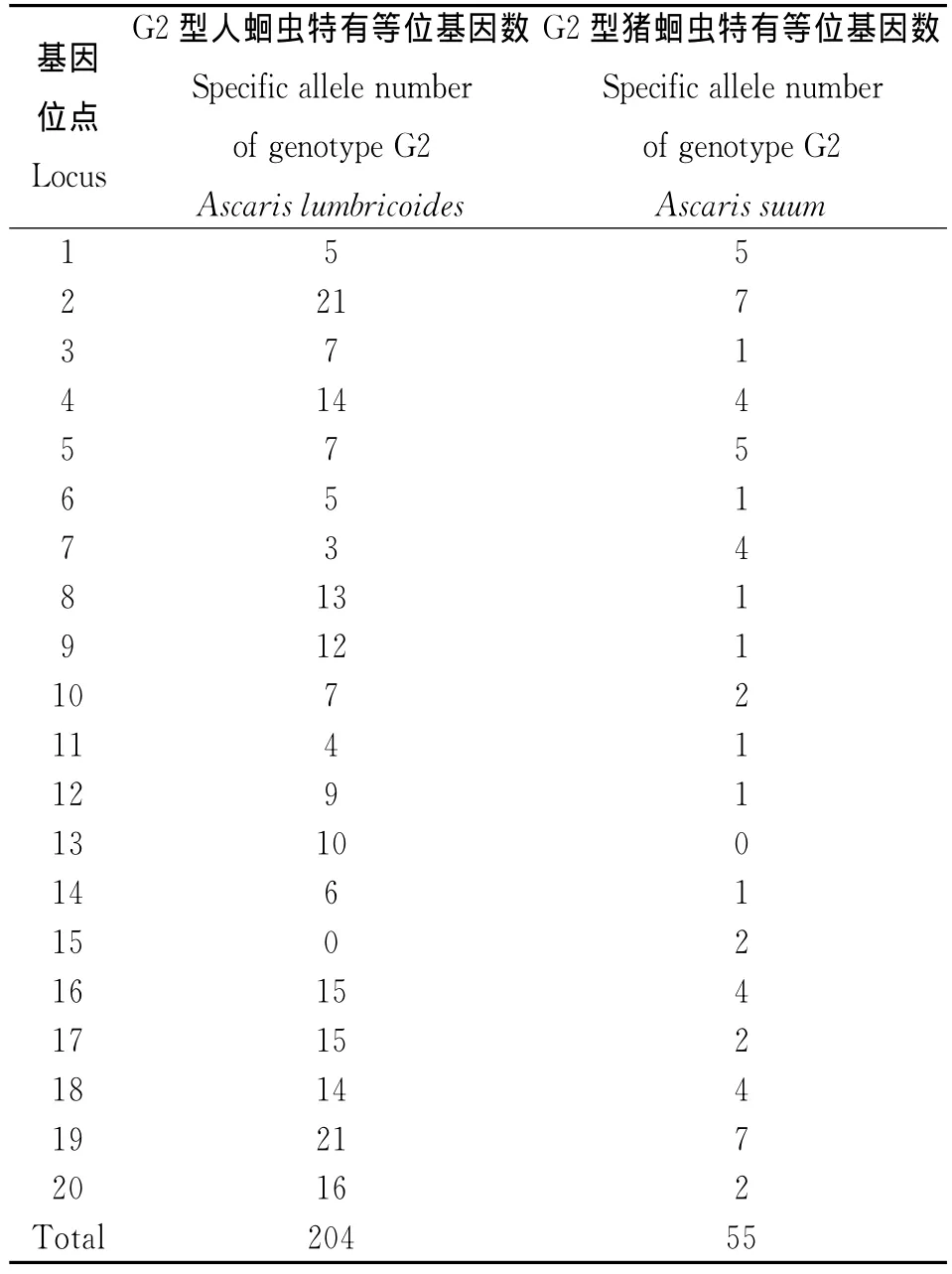
表2 G2型人蛔蟲和豬蛔蟲各位點特有等位基因數(shù)Tab.2 Number of unique alleles in Ascaris from humans and pigs with genotype G2
2.2 G2型人蛔蟲和豬蛔蟲的遺傳多樣性參數(shù)G2型人蛔蟲和豬蛔蟲的觀測雜合度H O,期望雜合度H E,多態(tài)信息含量PIC,詳見表3。結(jié)果表明,G2型人蛔蟲在各位點的觀測雜合度H O、期望雜合度H E和多態(tài)信息含量PIC分別為0.262~0.909、0.324~0.951和0.312~0.937,均值分別為0.565、0.802和0.779;G2型豬蛔蟲在各位點的觀測雜合度H O、期望雜合度H E和多態(tài)信息含量PIC分別為0.143~0.950、0.307~0.937和0.291~0.908,均值分別為0.533、0.777和0.737。
2.3 蛔蟲種群遺傳分化和遺傳結(jié)構(gòu) 本實驗對兩個蛔蟲種群的F統(tǒng)計量計算為:0.3218(F IT),0.3031(FIS),0.0268(FST)。兩種群間的基因流Nm值為9.08。分子變異方差分析(AMOVA)發(fā)現(xiàn),僅有2.6%的遺傳變異來自于種群間,其余68.6%的變異均來自于個體間,個體間遺傳變異大于種群間的遺傳變異(表4)。
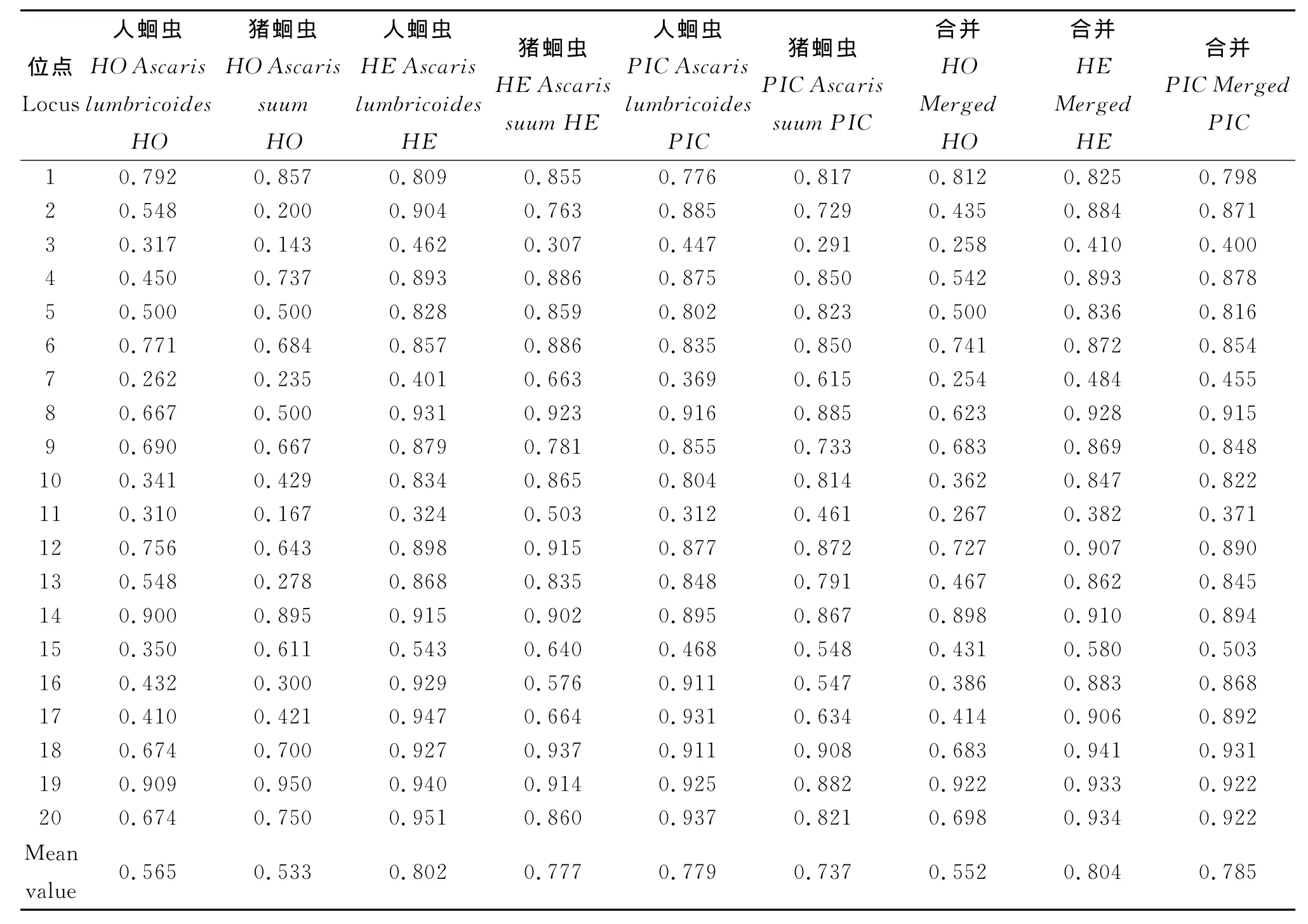
表3 G2型人蛔蟲和豬蛔蟲的觀測雜合度(HO)、期望雜合度(H E)和多態(tài)信息含量(PIC)Tab.3 Observed heterozygosity(HO),expected heterozygosity(H E),and polymorphic information content(PIC)in Ascaris from humans and pigs with genotype G2
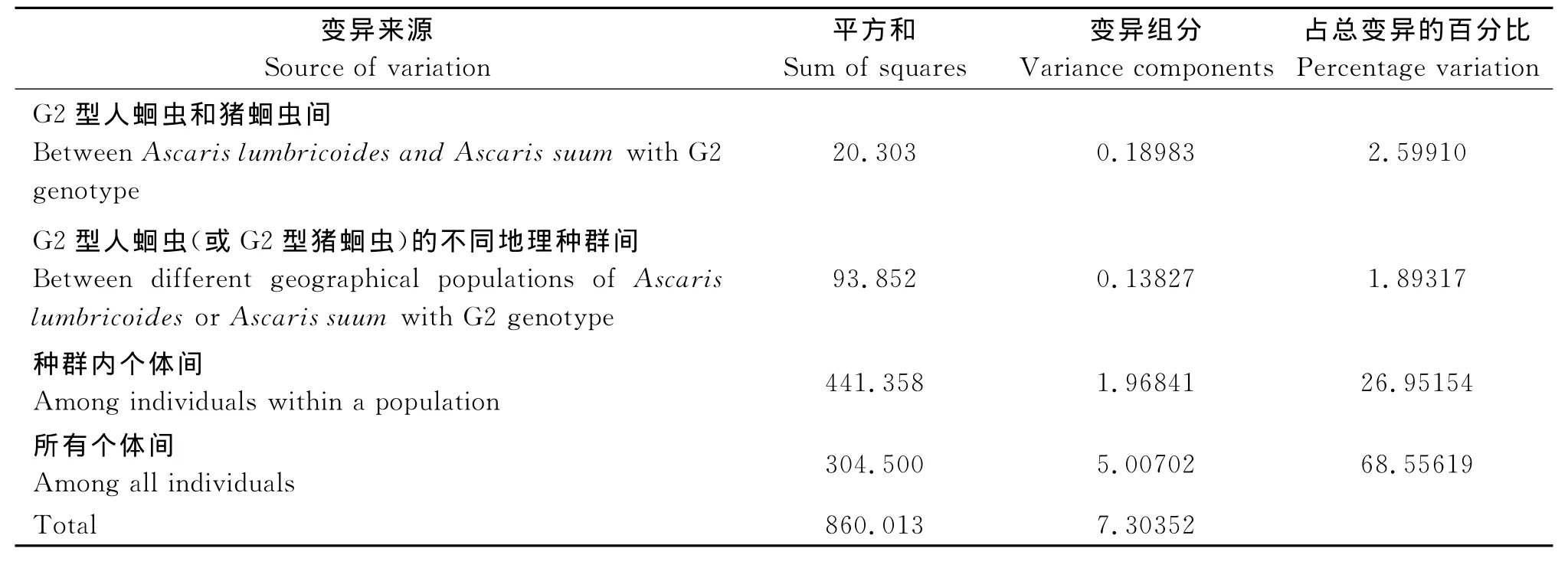
表4 G2型人蛔蟲和豬蛔蟲種群的分子變異方差分析Tab.4 Analysis of molecular variation of two Ascaris populations
3 討 論
3.1 蛔蟲微衛(wèi)星標記和等位基因 本研究結(jié)果表明所用的20個微衛(wèi)星位點具有較豐富的遺傳多態(tài)性。從各位點的等位基因頻率來看,其頻率分布不均,每個位點都有一種或幾個優(yōu)勢等位基因(即頻率最高的等位基因)存在。在20個基因位點中,有14個位點在2個蛔蟲種群中的優(yōu)勢等位基因一致,6個不一致。此外,位點中均存在各自的特有等位基因(表2)。種群中的優(yōu)勢等位基因往往是該物種中最保守和最原始的,而其余的等位基因則是優(yōu)勢等位基因在進化過程中因DNA突變等機制而形成。G2型的人蛔蟲和豬蛔蟲優(yōu)勢等位基因的不同以及特有等位基因的差異,可能和它們在宿主選擇上的差異[4-6]有關(guān)。
3.2 G2型人蛔蟲和豬蛔蟲種群遺傳多樣性 在微衛(wèi)星標記技術(shù)中,期望雜合度H E和多態(tài)信息含量PIC是用于估測種群遺傳多樣性的重要指標。G2型人、豬蛔蟲的平均H E分別為0.802和0.777(表3),說明各自都存在較大的遺傳變異。PIC是表示微衛(wèi)星位點變異程度高低的一個指標,本文中G2型人、豬蛔蟲種群在20個微衛(wèi)星位點都有較高的多態(tài)性。在人蛔蟲種群有4個中度、16個高度多態(tài)基因位點;而豬蛔蟲種群有2個中度、18個高度多態(tài)基因位點(表3)。同一微衛(wèi)星座位在不同蛔蟲宿主種群間以及同種宿主種群不同微衛(wèi)星座位間的多態(tài)信息含量都存在差異。前者反映了G2型人、豬蛔蟲間遺傳多樣性之間的差異,這種差異可能與宿主有關(guān);而G2型人蛔蟲或G2型豬蛔蟲中不同的微衛(wèi)星座位之間的多態(tài)信息的差異,可能和長期的自然和人為的選擇過程中(如驅(qū)蟲藥的使用),不同的微衛(wèi)星座位所受的選擇壓力不同有關(guān)。從G2型人蛔蟲和G2型豬蛔蟲在各位點的觀測雜合度H O、期望雜合度H E和多態(tài)信息含量PIC來看,G2型人蛔蟲的遺傳多樣性和G2型豬蛔蟲的相似,均有較高的遺傳多樣性,而G2型人蛔蟲的遺傳多樣性稍高于G2型豬蛔蟲。
3.3 G2型人蛔蟲和豬蛔蟲間的種群遺傳分化和基因流分析 種群間的遺傳分化系數(shù)FST是反映各種群遺傳分化的重要指標。本研究得出G2型人蛔蟲和G2型豬蛔蟲間的FST值為0.0268,根據(jù)Wright對FST值4個分級來判斷[14],說明G2型的兩種蛔蟲間遺傳分化水平較低,這不僅與現(xiàn)有種群內(nèi)高的遺傳多樣性有關(guān),還可能與種群間的基因流有關(guān)。基因流是種群遺傳結(jié)構(gòu)均質(zhì)化的主要因素之一,具有高水平基因流的物種往往比具有有限基因流的物種遺傳分化小,當Nm>4時,種群間的基因交流比較充分,均質(zhì)化作用足以抵制遺傳漂變的作用,可以防止種群間遺傳分化的產(chǎn)生;當Nm<1,遺傳漂變?yōu)榉N群遺傳結(jié)構(gòu)變化主要因素[15]。本研究G2型人蛔蟲和豬蛔蟲間的Nm為9.08,說明在蛔蟲進化過程中,遺傳漂變未成為群體遺傳結(jié)構(gòu)變化的主要因素,G2型人、豬蛔蟲基因交流機會較多,這和從線粒體基因研究得出的結(jié)果一致[10],而G2型人、豬蛔蟲雜交個體的客觀存在,即是這兩種蛔蟲之間較高水平基因流的證明[8]。
[1]Peng W,Criscione CD.Ascariasis in people and pigs:new inferences from DNA analysis of worm populations[J].Infect Genet Evol,2012,12(2):227-235.DOI:10.1016/j.meegid.2012.01.012
[2]Peng W,Yuan K,Zhou X,et al.Molecular epidemiological survey ofAscarisgenotypes in China based on single strand conformation polymorphism analysis of ribosomal DNA[J].Electrophoresis,2003, 24 (14): 2308-2315. DOI: 10.1002/elps.200305455
[3]Peng W,Yuan K,Hu M,et al.Mutation scanning-coupled analysis of haplotypic variability in mitochondrial DNA regions reveals low gene flow between human and porcineAscarisin endemic regions of China[J].Electrophoresis,2005,26(22):4317-4326.DOI:10.1002/elps.200500276
[4]Peng W,Yuan K,Hu M,et al.Experimental infections of pigs and mice with selected genotypes ofAscaris[J].Parasitology,2006,133(Pt 5):651-657.DOI:10.1017/S0031182006000643
[5]Peng W,Yuan K,Peng G,et al.Ascaris:Development of selected genotypes in mice[J].Exp Parasitol,2012,131(1):69-74.DOI:10.1016/j.exppara.2012.03.006
[6]Peng W,Yuan K,Hu M,et al.Recent insights into the epidemiology and genetics ofAscarisin China using molecular tools[J].Parasitology,2007,134(Pt 3):325-330.DOI:10.1017/S0031182006001521
[7]Criscione CD,Anderson JD,Sudimack D,et al.Disentangling hybridization and host colonization in parasitic roundworms of humans and pigs[J].Proc Boil Sci,2007,274(1626):2669-2677.DOI:10.1098/rspb.2007.0877
[8]Zhou C,Li M,Yuan K,et al.PigAscaris:An important source of human ascariasis in China[J].Infect Genet Evol,2012,12(6):1172-1177.DOI:10.1016/j.meegid.2012.04.016
[9]Zhou C,Yuan K,Tang X,et al.Molecular genetic evidence for polyandry inAscarissuum[J].Parasitol Res,2011,108(3):703-708.DOI:10.1007/s00436-010-2116-3
[10]Zhou C,Li M,Yuan K,et al.Phylogeography ofAscarislumbricoidesandA.suumfrom China[J].Parasitol Res,2011,109(2):329-338.DOI:10.1007/s00436-011-2260-4
[11]Marshall TC,Slate J,Kruuk LE,et al.Statistical confidence for likelihood-based paternity inference in natural populations[J].Mol Ecol,1998,7(5):639-655.DOI:10.1046/j.1365-294x.1998.00374.x
[12]Belkhir K,Borsa P,Chikhi L,et al.GENETIX,a WindowsTMbased software for population genetic analyses[OL].Laboratoire Génome,Populations,Interactions CNRSUMR 5000,Universitéde MontpellierⅡ,Montpellier,F(xiàn)rance,2000.http://www.univmontp2.fr^genetix/genetix.htm
[13]Excoffier L,Laval G,Schneider S.Arlequin(version 3.0):an integrated software package for population genetics data analysis[J].Evol Bioinform Online,2007,1:47-50.
[14]Wright S.Evolution and the genetics of population,Vol 4.Variability within and among natural populations[M].Chicago:University of Chicago Press,1978.
[15]Whitlock MC,McCauley DE.Indirect measures of gene flow and migration:[J].Heredity (Edinb.),1999,82(Pt 2):117-125.
