PERK通路參與了缺氧心肌細胞的內質網應激及凋亡*
2012-09-07 03:31:40劉春蕾李蕊君何云云何昆侖王莉莉
中國病理生理雜志 2012年8期
劉春蕾, 李 鑫, 李蕊君, 何云云, 何昆侖△, 王莉莉
(1中國人民解放軍總醫院心內科,北京 100853;2南開大學醫學院,天津 300071;3軍事醫學科學院毒物藥物研究所,北京 100850)
PERK通路參與了缺氧心肌細胞的內質網應激及凋亡*
劉春蕾1,2, 李 鑫1, 李蕊君1, 何云云1,2, 何昆侖1△, 王莉莉3△
(1中國人民解放軍總醫院心內科,北京 100853;2南開大學醫學院,天津 300071;3軍事醫學科學院毒物藥物研究所,北京 100850)
目的:觀察缺氧對原代培養的Wistar乳鼠心肌細胞的損傷,探討內質網應激在缺氧心肌損傷發生發展過程中起的作用及PERK通路是否參與其信號轉導過程。方法:將原代培養的乳鼠心肌細胞隨機分為正常對照組和缺氧1 h、4 h、8 h、12 h、24 h組,通過測定細胞ATP含量反映細胞活力;高內涵分析細胞成像系統檢測多參數凋亡;采用免疫細胞化學和蛋白印跡方法檢測以內質網為靶點的分子伴侶(GRP78和鈣網蛋白)的表達,PERK通路(PERK和eIF2α)的磷酸化水平,以及其下游分子(ATF4和CHOP)在缺氧不同時點蛋白的表達變化特征。采用PERK通路激活型藥物salubrinal處理原代培養的Wistar乳鼠心肌細胞,觀察藥物是否對缺氧損傷的心肌細胞有保護作用。結果:缺氧引起心肌細胞凋亡,缺氧早期(約1 h)鈣網蛋白和GPR78的表達上調;缺氧中期(4 h)p-PERK、p-eIF2α和ATF4的表達上調;缺氧后期(12 h)CHOP的表達上調。Salubrinal對缺氧心肌有保護作用。結論:在培養的心肌細胞中,缺氧可激發內質網應激。在缺氧早期激活PERK通路保護機體對抗缺氧損傷,后期激活細胞凋亡通路。
心肌細胞;內質網應激;缺氧;未折疊蛋白反應;細胞凋亡
心肌缺血觸發心肌細胞發生持續性凋亡反應,使心肌細胞進行性丟失、膠原支架斷裂、心室擴張和心室重塑,是多種心臟不良事件發生的重要原因[1]。因此,深入闡明缺氧心肌細胞凋亡的分子機制可望為缺血缺氧相關的心臟疾病提供新的有效干預手段。
傳統的細胞凋亡的途徑又稱caspase依賴的細胞死亡途徑,主要包括死亡受體途徑和線粒體途徑,兩者均通過誘導caspase級聯反應激活caspase-3,導致細胞凋亡。近年的研究顯示,除了caspase依賴的細胞凋亡途徑外,內質網應激(endoplasmic reticulum stress,ERS)也與細胞凋亡密切相關[2]。ERS是由于細胞內外環境的改變引起ER內未折疊蛋白質聚集,鈣穩態失衡,從而導致的內質網功能紊亂的狀態。在ERS早期,未折疊蛋白反應(unfolded protein response,UPR)是重要特征,具有保護細胞抵抗外界環境對細胞的干擾作用;然而,一旦ERS持續而嚴重,則激活凋亡通路而誘發細胞凋亡[3]。研究表明,UPR可由3條獨立的信號通路所誘導,包括活化轉錄因子6(activating transcription factor 6,ATF6)、肌醇需酶(inositol-requiring enzyme 1,IRE1)和蛋白激酶R樣內質網激酶(protein kinase R-like ER kinase,PERK)[4]。目前,在心肌細胞中,缺氧誘發細胞ERS以及凋亡已經得到廣泛證實。Negoro等[5]和 Ron等[6]觀察到,缺氧確能誘導新生乳鼠心肌細胞產生UPR反應。也有研究發現,心肌細胞在缺氧條件下,ATF6通路蛋白表達上調[7],誘發ATF6和IRE1通路共同的下游分子X盒結合蛋白1(X-box-binding protein 1,XBP1)及凋亡蛋白的表達,并隨著缺氧時間延長,最終使細胞死亡[8]。由此可見,ATF6和IRE1通路參與了缺氧心肌細胞ERS及凋亡。相比較ATF6和IRE1通路而言,PERK抑制蛋白合成途徑,更容易通過增強核糖體真核生物起始因子2(eukaryotic initiation factor 2,eIF2α)磷酸化酶的活性或抑制其去磷酸化酶的活性,抑制細胞凋亡。然而,PERK通路是否也參與了心肌細胞缺氧過程尚不清楚。本實驗通過缺氧誘導原代培養乳鼠心肌細胞凋亡,采用免疫細胞化學和蛋白印跡的方法,觀察了缺氧不同時間對內質網應激及凋亡相關蛋白表達的影響,特別是PERK通路相關蛋白的變化特點,旨在揭示PERK信號通路與缺氧誘導心肌細胞損傷的關系。
材料和方法
1 材料
新生24 h內的Wistar大鼠,雌雄不拘(由軍事醫學科學院實驗動物中心提供),動物合格證號為SCXK-(軍)2007-004。小鼠抗大鼠GAPDH單克隆抗體、兔抗大鼠葡萄糖調節蛋白質78(glucoseregulated protein,GRP78)多克隆抗體購自Stressgen。兔抗大鼠鈣網蛋白(calreticulin,Cal)多克隆抗體、兔抗大鼠phospho-PERK多克隆抗體、兔抗大鼠PERK多克隆抗體、兔抗大鼠eIF2α多克隆抗體、兔抗大鼠phospho-eIF2α多克隆抗體和小鼠抗大鼠C/EBP同源蛋白質(C/EBP homologous protein,CHOP)多克隆抗體購自Cell Signaling Technology。兔抗大鼠活化轉錄因子4(activating transcription factor 4,ATF4)多克隆抗體購自Abcam。山羊抗小鼠IgG DylightTM594購自 Thermo scientific。ATP試劑盒購自 Promega。Salubrinal(Sal)和Hoechst 33258購自Invitrogen。高內涵分析系統 In cell Analyzer 2000為 GE產品; Western blotting成像與分析采用的Alphamager 5500凝膠成像系統為Alpha產品。
2 方法
2.1 心肌細胞原代培養 迅速取出新生Wistar大鼠心臟,剪碎,用終濃度為0.125%胰蛋白酶和0.1%膠原酶在37℃水浴中消化20 min,用含10%FBS的DMEM培養基中和蛋白酶,將細胞懸液離心并重懸細胞接種至75 cm2培養瓶或培養板中,37℃、5%CO2孵箱培養24 h后半量換液;之后每48 h換液1次。4 d后,缺氧處理的細胞置于密閉的恒溫缺氧倉內,通入95%N2/5%CO2的混合氣體,O2含量少于0.5%。
2.2 細胞活力測定 純化的乳鼠心肌細胞按1× 104cells/well接種到96孔板37℃、5%CO2孵箱培養4 d,之后置缺氧箱中分別缺氧處理細胞1 h、4 h、8 h、12 h、24 h,每時點設3個重復孔。同時設置藥物干預組,細胞缺氧處理前 24 h使用不同濃度的PERK通路激活型藥物salubrinal孵育心肌細胞30 min,按照細胞活力檢測試劑盒說明檢測細胞活性。采用酶標儀檢測562 nm波長下的熒光值。
2.3 多參數凋亡檢測試劑盒檢測細胞凋亡[9]每孔加入10 000個細胞,在37℃、95%N2、5%CO2的缺氧箱中孵育36 h,加入5%線粒體膜電位和DNA熒光素染料MitoTracker/Hoechst溶液,室溫染色10 min,PBS清洗后,細胞進行透化處理,鬼筆環肽(Alexa Fluor 488 phalloidin solution)室溫暗處理30 min。剩余的溶液用PBS洗3次。應用特異性的DNA熒光素染料 Hoechst 33258,線粒體膜電位染料 Mito-Tracker,細胞骨架染料Alexa Fluor 488進行染色后,用高內涵分析細胞成像系統在波長350/461 nm、495/519 nm和579/599 nm下進行檢測觀察。檢測內容包括細胞形態、細胞核面積、DNA含量、核強度、肌動蛋白含量、線粒體膜電位等指標。正常細胞核為圓形或橢圓形,熒光著色淺,凋亡細胞核固縮,呈強熒光反應;與正常細胞相比,凋亡細胞骨架微絲蛋白含量降低,線粒體膜電位下降等現象
2.4 Western blotting法檢測心肌細胞蛋白表達 純化的乳鼠心肌細胞按1×106cells/well接種到6孔板,在37℃、5%CO2孵箱培養4 d后,在95%N2、5%CO2的缺氧箱中孵育不同時間。用1%SDS裂解液(mmol/L:NaCl 100,EDTA 0.1,DTT 10,HEPES 50)處理心肌細胞,離心取上清,用BCA法進行蛋白定量。于100℃水浴加熱10 min,12 000×g、4℃離心10 min。經10%SDS-PAGE電泳分離后,將目標條帶電轉至PVDF膜,其中PERK和p-PERK用25 V半干式轉移30 min,其余采用15 V半干式轉移15 min;之后,采用5%脫脂奶粉室溫封閉電轉后的PVDF膜1 h,Ⅰ抗分別為GRP78(1∶1 000)、calreticulin(1∶200)、PERK(1∶1 000)、p-PERK(1∶1 000)、phospho-eIF2α(1∶1 000)、eIF2α(1∶1 000)、ATF4(1∶500)、GAPDH(1∶200)、β-actin(1∶500),4℃孵育過夜,TBS/T洗3次,加入Ⅱ抗(1∶5 000)室溫孵育1 h,TBS/T洗3次;之后加SuperECL Plus超敏發光液避光孵育 3 min,最后采用 AlphaImager 5500凝膠成像系統進行成像拍攝。用 FluorChem 5500進行蛋白量灰度分析。
2.5 免疫細胞化學方法檢測CHOP蛋白表達 純化的乳鼠心肌細胞按1×104cells/well接種到黑色底透96孔板,在37℃、5%CO2孵箱培養4d后,置于37℃、95%N2和5%CO2缺氧箱中孵育不同時間。之后,細胞用4%多聚甲醛固定15 min。封閉液室溫封閉1 h;1∶1 600稀釋Ⅰ抗CHOP,4℃孵育過夜; PBS洗3次,隨后用含Hoechest 33258(3 μmol/L)的熒光Ⅱ抗(1∶500)37℃孵育2 h。洗后,用In Cell Analyzer 1000高內涵分析細胞成像系統檢測,采用In Cell Analyzer Workstation進行熒光強度分析。
3 統計學處理
數據以均數±標準差(珋x±s)表示,采用SPSS 13.0統計軟件,組間比較采用單因素方差分析(One way-ANOVA),以P<0.05為差異有統計學意義。
結果
1 缺氧對大鼠心肌細胞ATP含量的影響
細胞ATP含量測定表明,與正常對照組相比,缺氧1 h、4 h、8 h、12 h、24 h后,細胞ATP含量顯著下降(P<0.05),隨著缺氧時間的延長,細胞ATP含量呈減低趨勢,但缺氧4 h較缺氧1 h有一升高,見圖1。
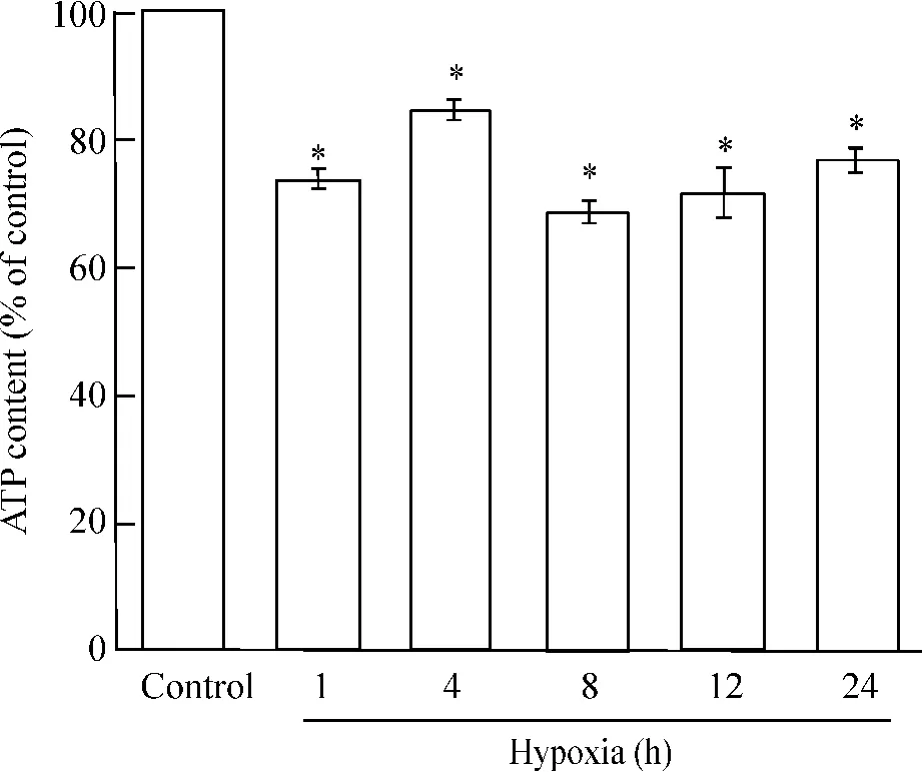
Figure 1. The effect of hypoxia on the viability of primary neonatal rat cardiomyocytes at different time points.x珋±s.n=3.*P<0.05 vs control(non-hypoxia treatment) group.圖1 缺氧不同時點對原代培養新生大鼠心肌細胞ATP含量的影響
2 凋亡心肌細胞的鑒定
多參數凋亡檢測試劑盒檢測細胞凋亡的結果顯示:缺氧后,細胞出現核固縮、骨架微絲蛋白含量下降、線粒體膜電位下降等凋亡細胞的特征,見圖2。從這3個參數的數據分析顯示缺氧16 h和24 h造成明顯的細胞凋亡,核熒光強度分別較對照組高26.3%和26.6%;細胞骨架微絲含量分別較對照組低5%和11%;線粒體膜電位分別較對照組低17%和27%(P<0.05)。
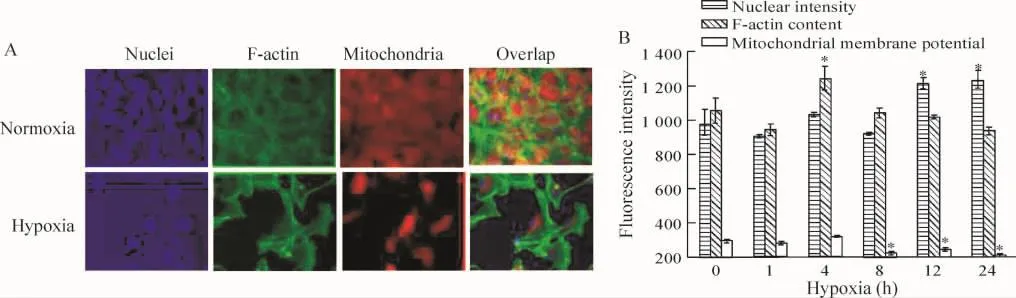
Figure 2. The effect of hypoxia on the apoptosis of the cardiomyocytes detected by high-content analysis.A:the images with nuclear staining,F-actin staining and mitochondrial membrane potential staining under normoxia or hypoxia for 24 h(×400);B: the nuclear intensity,F-actin content and mitochondrial membrane potential were statistical analyzed.珋x±s.n=3.*P<0.05 vs control group(0 h).圖2 多參數檢測缺氧不同時間后心肌細胞的凋亡情況
3 Sal對缺氧誘導心肌細胞ATP含量的影響
細胞ATP含量測定發現,提前30 min給予心肌細胞salubrinal 10、20、40 μmol/L,再缺氧培養24 h后,與單純缺氧組相比,salubrinal預給后增加細胞活力,細胞ATP含量顯著上升,見圖3。
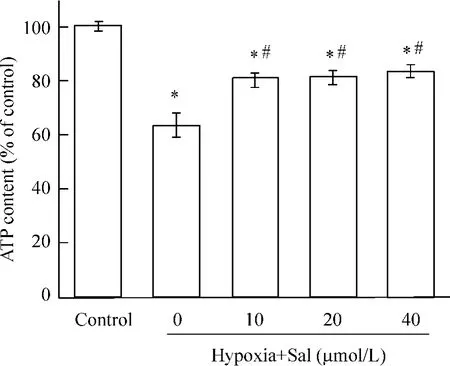
Figure 3. The protective effect of salubrinal(Sal)on hypoxic cardiomyocytes±s.n=3.#P<0.05 vs hypoxia+ Sal(0 μmol/L)group;*P<0.05 vs control group.圖3 不同濃度salubrinal對缺氧心肌細胞的保護作用
4 缺氧對心肌細胞內質網應激相關因子的影響
4.1 缺氧對心肌細胞內質網應激標志分子(鈣網蛋白和GRP78)表達的影響 與正常對照相比,缺氧1 h,心肌細胞內質網應激激活蛋白GRP78、鈣網蛋白顯著增加,其中鈣網蛋白較對照組高72%(P<0.05),見圖 4A,GRP78較對照組高 65%(P<0.05),見圖4B;但隨著缺氧時間的延長,鈣網蛋白在缺氧4、8和12 h與對照組表達量相當,缺氧24 h顯著低于對照組;而GRP78在缺氧4、8 h與對照組表達量相當,缺氧12 h、24 h顯著低于對照組。上述結果表明,心肌細胞內質網應激分子鈣網蛋白和GRP78表達均呈現早期升高,后期下降的趨勢。
4.2 缺氧對PERK和eIF2α磷酸化水平及ATF4蛋白表達的影響 Western blotting測定結果顯示,與正常對照組相比,從缺氧 1 h起,心肌細胞 p-PERK/PERK開始顯著升高,至缺氧4 h,p-PERK/ PERK達到最大,較對照組高73%,隨后逐漸下降,見圖5A,缺氧8 h仍保持較高水平,但到12 h后則低于對照水平;缺氧心肌細胞eIF2α的磷酸化水平也是從缺氧1 h起開始顯著升高,至缺氧4 h,peIF2α/eIF2α達到最大,較對照組高87%(P<0.05),見圖5B,隨后雖逐漸下降,但直至缺氧24 h仍然保持較高表達水平;對照組ATF4不表達,缺氧后,細胞中ATF4蛋白表達量升高,在4 h達到最高峰,隨后顯著下降(P<0.05),見圖5C,但至缺氧24 h仍有表達。
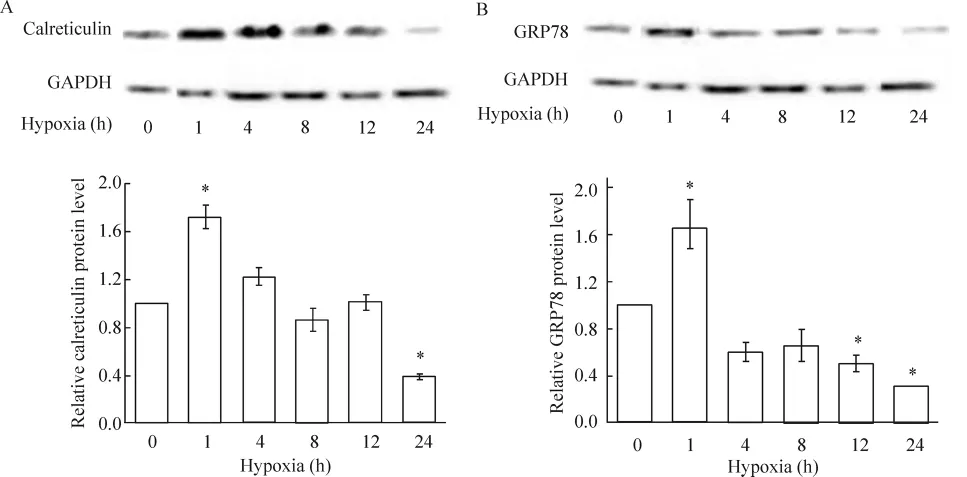
Figure 4. Effect of hypoxia on calreticulin(A)and GRP78(B)protein expression.珋x±s.n=3.*P<0.05 vs control(0 h)group.圖4 缺氧不同時間對鈣網蛋白和GRP78蛋白表達的影響
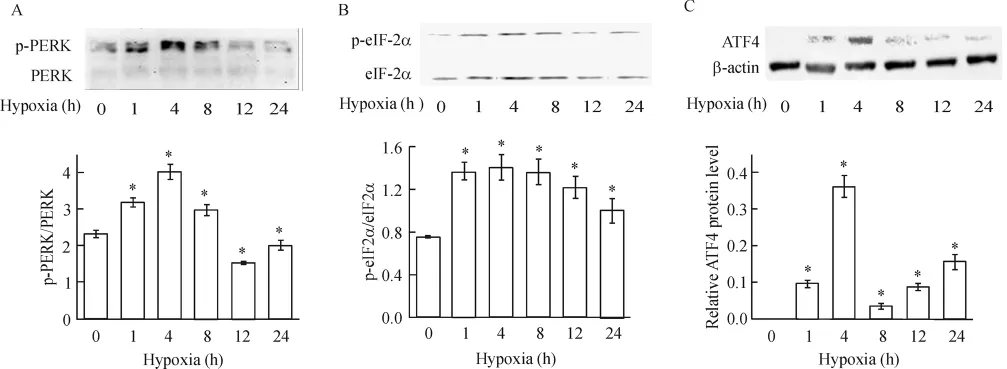
Figure 5. Effect of hypoxia on p-PERK/PERK(A),p-eIF2α/eIF2α(B)and ATF4(C)protein expression.珋x±s.n=3.*P<0.05 vs control(0 h)group.圖5 缺氧不同時間對p-PERK/PERK、p-eIF2α/eIF2α及ATF4蛋白表達的影響
4.3 缺氧對內質網應激特異性凋亡分子(CHOP)表達的影響 由免疫細胞化學結果可以看出,內質網應激特異性凋亡分子CHOP在非缺氧處理的對照組和短期缺氧組心肌細胞上不表達,但在缺氧12 h和 24 h,CHOP出現表達。可見,缺氧可激活PERK通路下游的凋亡活性分子CHOP,缺氧12 h心肌細胞凋亡相關蛋白達到較高水平(P<0.05),見圖6。
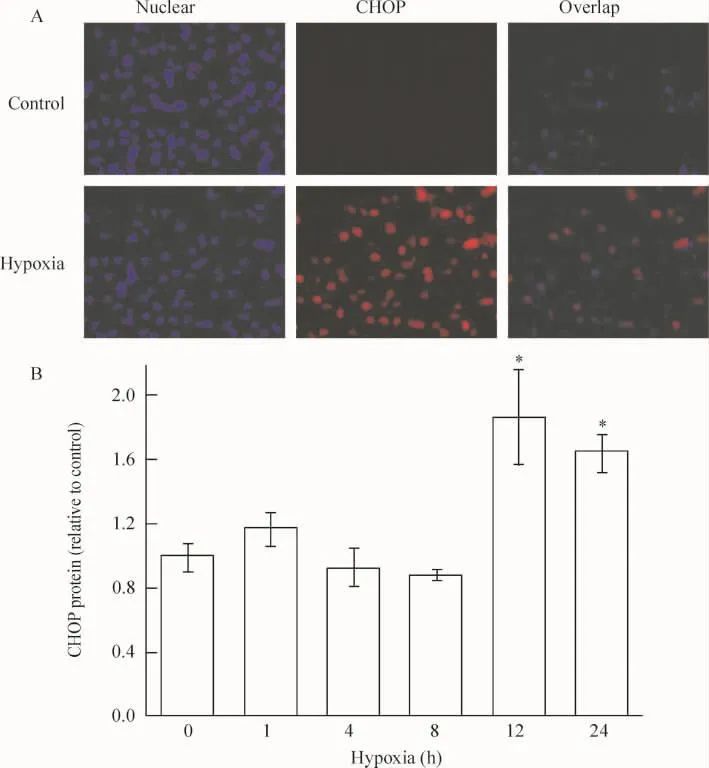
Figure 6. Effect of hypoxia on CHOP protein expression.A:the representative images of high-content analysis(×200);B:the fluorescence intensity analysis of CHOP.珋x±s.n=3.*P<0.05 vs control(0 h)group.圖6 免疫細胞化學檢測缺氧對CHOP蛋白表達水平的影響
討論
組織缺氧是指因氧氣供應不足或用氧障礙,而導致組織代謝、功能和形態結構發生異常變化的病理過程[9-10]。心肌缺氧可導致以細胞壞死和凋亡為特征的心肌細胞損傷。近年研究表明,ERS相關的凋亡途徑與缺氧誘發細胞凋亡密切相關[11-12]。UPR保護性機制主要由2條相對獨立的內質網跨膜分子介導信號通路來調控:一條是IRE1和ATF6途徑,即IRE1和ATF6通過誘導內質網分子伴侶使未折疊或錯誤折疊蛋白恢復正確構像,終止ERS和細胞凋亡;另一條是PERK途徑,即PERK在翻譯水平抑制蛋白合成,減少未折疊蛋白,最終促進ERS恢復,減少細胞凋亡。有實驗表明ATF6和IRE1通路參與了缺氧誘發的心肌細胞ERS及凋亡。本實驗在體外培養的乳鼠心肌細胞上發現,PERK通路在缺氧心肌細胞損傷中起到重要的作用,缺氧不僅能夠在內質網應激早期激活UPR的PERK通路,對心肌細胞起到保護作用,當內質網應激時間延長,還能夠激活其下游的凋亡信號分子,激活凋亡通路,引發心肌細胞凋亡。
本實驗將原代培養的乳鼠心肌細胞置于密閉的恒溫缺氧倉內,通入95%N2/5%CO2混合氣體,氧氣含量少于0.5%,模擬體內心肌細胞缺氧環境。以ATP含量作為細胞活力的檢測指標,結果顯示,隨著缺氧時間的延長,細胞活力明顯降低,但在缺氧早期,大約4 h,細胞活力有所逆轉。多參數檢測細胞凋亡的實驗同樣發現,在缺氧時間延長至12 h,無論從細胞的核亮度,還是線粒體膜電位及肌動蛋白含量的改變上來看,心肌細胞均表現出明顯的凋亡特征,但在缺氧早期表現出輕微逆轉。這說明在內質網應激早期,有一種保護性作用的存在對抗由缺氧引起的心肌細胞損傷。本實驗設計不同缺氧時間體現了內質網應激對心肌細胞造成損傷的不同。內質網分子伴侶GRP78作為內質網穩態的感受器,在監測內質網中未折疊蛋白質的聚集和內質網應激的激活過程中發揮重要作用。鈣網蛋白為ER中主要的鈣結合伴侶蛋白,是ER貯存的重要應激蛋白和ERS的重要標志。本實驗中,缺氧1 h,就能上調GRP78和鈣網蛋白的表達,缺氧4、8 h與正常組表達量相當,缺氧12、24 h顯著低于正常組,提示缺氧1 h后導致了內質網應激反應發生。有文獻多次報道稱,在缺氧模型中,GRP78和鈣網蛋白作為內質網應激的誘導因子,在應激前期被激活表達[13-14]。上述實驗現象與文獻報道一致,表明本實驗的缺氧模型是成功的。
eIF2α是ERS中PERK通路直接的調控因子,PERK通過催化eIF2α磷酸化,使GDP-eIF2α不能轉化為GTP-eIF2α,不能形成GTP-eIF2α-MettRNA/40S復合體,結果Met-tRNA的翻譯被終止,蛋白合成下降,未折疊蛋白減少等,最終促進ERS恢復,減少細胞凋亡。Boyce等[15]報道,p-eIF2α去磷酸化抑制劑salubrinal通過激活UPR具有對抗ERS相關的PC-12細胞凋亡的作用。本實驗室前期工作也證實在原代培養的 Wistar大鼠心肌細胞上,salubrinal具有對抗ERS特異性誘導劑衣霉素(tunicamycin)引起的心肌細胞損傷的作用,并且證實salubrinal的保護作用與eIF2α的磷酸化水平相關[16]。因此,salubrinal對細胞的保護作用可以間接證明PERK通路參與了ERS相關的細胞凋亡。本實驗在缺氧誘導的原代培養的乳鼠心肌細胞上,發現提前給予salubrinal組ATP含量較單純缺氧組有所回升,即抑制p-eIF2α的去磷酸化,對缺氧心肌細胞損傷有保護作用。以上結果提示,缺氧激活了UPR中的PERK通路。
PERK位于內質網內,作為未折疊蛋白反應中的重要因子,在發生內質網應激時與游離的GRP78結合而使自身蛋白暴露,發生磷酸化與二聚化。活化的PERK蛋白進一步使內質網外側的eIF2α發生磷酸化修飾,一方面通過eIF2α磷酸化抑制蛋白合成,另一方面激活下游的ATF4,ATF4合成后轉位至細胞核內,作為轉錄因子能夠上調內質網分子伴侶蛋白并參與氨基酸轉運蛋白質的轉錄表達,同樣有助于恢復內質網穩態[17-18]。本實驗在原代培養的心肌細胞缺氧模型上檢測缺氧是否激活了PERK通路,結果發現:在缺氧4 h時,PERK和eIF2α磷酸化水平與對照組相比,都顯著升高。ATF4作為peIF2α的下游分子,在缺氧后4 h表達上調,明顯高于對照組。GADD153/CHOP是內質網應激特異的凋亡蛋白,屬C/EBP轉錄因子家族成員。正常情況下,CHOP表達十分低,在內質網應激反應時,CHOP作為PERK通路的下游分子,在缺氧12 h,表達明顯增加。在哺乳動物中,調節蛋白合成是一個十分復雜的過程,我們不能夠排除其它小分子蛋白來調節蛋白合成的可能。但是我們可以推測,在缺氧早期,缺氧激活UPR中的PERK通路,并且通過PERK的磷酸化使eIF2α磷酸化,并且誘導ATF4表達上調,起到部分抑制未折疊蛋白合成、保護細胞的作用。但在缺氧后期,CHOP作為PERK通路的下游分子,誘發細胞心肌凋亡,是導致心肌細胞缺氧損傷的一個重要原因。阻斷PERK通路,減少CHOP的表達,對于減少細胞凋亡的發生將起到決定性作用,有關這方面的討論有待進一步研究。
綜上所述,本實驗在原代培養的大鼠心肌細胞中發現,UPR反應中PERK通路蛋白及其下游的凋亡信號在缺氧過程中被激活,并在缺氧心肌細胞中依次表達。即ERS相關的PERK通路參與了缺氧誘發的心肌細胞內質網應激及凋亡。針對PERK通路與缺氧誘導心肌細胞損傷關系的研究,將有望為心肌缺血性疾病的防治提供新靶點。
[1]Wencker D,Chandra M,Nguyen K,et al.A mechanistic role for cardiac myocyte apoptosis in heart failure[J].J Clin Invest,2003,111(10):1497-1504.
[2]Szegezdi E,Duffy A,O'Mahoney ME,et al.ER stress contributes to ischemia-induced cardiomyocyte apoptosis[J].Biochem Biophys Res Commun,2006,349(4): 1406-1411.
[3]Faitova J,Krekac D,Hrstka R,et al.Endoplasmic reticulum stress and apoptosis[J].Cell Mol Biol Lett,2006,11(4):488-505.
[4]Davenport EL,Morgan GJ,Davies FE.Untangling the unfolded protein response[J].Cell Cycle,2008,7(7): 865-869.
[5]Negoro S,Kunisada K,Tone E,et al.Activation of JAK/ STAT pathway transduces cytoprotective signal in rat acute myocardial infarction[J].Cardiovasc Res,2000,47(4): 797-805.
[6]Ron D,Walter P.Signal integration in the endoplasmic reticulum unfolded protein response[J].Nat Rev Mol Cell Biol,2007,8(7):519-529.
[7]Doroudgar S,Thuerauf DJ,Marcinko MC,et al.Ischemia activates the ATF6 branch of the endoplasmic reticulum stress response[J].J Biol Chem,2009,284(43):29735-29745.
[8]Thuerauf DJ,Marcinko M,Gude N,et al.Activation of the unfolded protein response in infarcted mouse heart and hypoxic cultured cardiac myocytes[J].Circ Res,2006, 99(3):275-282.
[9]Gasparetto M,Gentry T,Sebti S,et al.Identification of compounds that enhance the anti-lymphoma activity of rituximab using flow cytometric high-content screening[J].J Immunol Methods,2004,292(1-2):59-71.
[10]Natarajan R,Salloum FN,Fisher BJ,et al.Prolyl hydroxylase inhibition attenuates post-ischemic cardiac injury via induction of endoplasmic reticulum stress genes[J].Vascul Pharmacol,2009,51(2-3):110-118.
[11]Osada N,Kosuge Y,Ishige K,et al.Characterization of neuronal and astroglial responses to ER stress in the hippocampal CA1 area in mice following transient forebrain ischemia[J].Neurochem Int,2010,57(1):1-7.
[12]楊 鵬,楊成明,曾春雨,等.心肌梗死后心力衰竭小鼠心肌組織內質網應激相關凋亡途徑的研究[J].中國病理生理雜志,2010,26(6):1069-1074.
[13]Minamino T,Kitakaze M.ER stress in cardiovascular disease[J].J Mol Cell Cardiol,2010,48(6):1105-1110.
[14]Flores-Diaz M,Higuita JC,Florin I,et al.A cellular UDP-glucose deficiency causes overexpression of glucose/oxygen-regulated proteins independent of the endoplasmic reticulum stress elements[J].J Biol Chem,2004,279(21):21724-21731.
[15]Boyce M,Bryant KF,Jousse C,et al.A selective inhibitor of eIF2α dephosphorylation protects cells from ER stress[J].Science,2005,307(5711):935-939.
[16]胡國梁,何昆侖,范 利,等.Salubrinal保護心肌細胞內質網應激凋亡的實驗研究[J].軍醫進修學院學報,2010,31(5):483-485.
[17]Terai K,Hiramoto Y,Masaki M,et al.AMP-activated protein kinase protects cardiomyocytes against hypoxic injury through attenuation of endoplasmic reticulum stress[J].Mol Cell Biol,2005,25(21):9554-9575.
[18]Cybulsky AV,Takano T,Papillon J,et al.Role of the endoplasmic reticulum unfolded protein response in glomerular epithelial cell injury[J].J Biol Chem,2005,280 (26):24396-24403.
PERK signal pathway is involved in hypoxia-induced endoplasmic reticulum stress and apoptosis in cultured cardiac myocytes
LIU Chun-lei1,2,LI Xin1,LI Rui-jun1,HE Yun-yun1,2,HE Kun-lun1,WANG Li- li3
(1Department of Cardiology,General Hospital of Chinese PLA,Beijing 100853,China;2Medical School of Nankai University,Tianjin 300071,China;3Institute of Poisons and Drugs,Academy of Military Medical Sciences,Beijing 100850,China.E-mail:hekunlun2002@yahoo.com;wangll63@126.com)
AIM:To investigate the role of endoplasmic reticulum(ER)stress in the process of hypoxia-induced neonatal rat myocardial injury through PERK signal pathway.METHODS:Neonatal rat cardiac myocytes were randomly divided into control group and hypoxia 1 h,4 h,8 h,12 h and 24 h groups.Cell viability was evaluated by determining the intracellular content of ATP.Apoptosis was measured by high-content analysis(HCA)cell imaging system.The protein levels of GRP78,calreticulin,p-PERK,p-eIF2α,ATF4 and CHOP were detected by Western blotting at different time points.The primary cultured neonatal rat cardiac myocytes were treated with an agonist of PERK pathway salubrinal and the cell apoptosis was observed under hypoxia.RESULTS:In the early phase,hypoxia induced an increase in the expression of calreticulin and GPR78.In the middle phase of hypoxia,the levels of p-PERK,p-eIF2α and ATF4 were increased.In the later phase of hypoxia,increased CHOP level was also observed.Salubrinal effectively protected the cardiac myocytes from hypoxic injury.CONCLUSION:Hypoxia activates ER stress in cardiac myocytes and also activates PERK signal pathway.PERK signaling protects cardiac myocytes from hypoxic damage in the early stage and triggers apoptosis of the cells in the later phase.
Cardiomyocytes;Endoplasmic reticulum stress;Hypoxia;Unfolded protein response;Apoptosis
R363.2
A
10.3969/j.issn.1000-4718.2012.08.010
1000-4718(2012)08-1392-07
2012-03-02
2012-06-25
國家科技部國際科技合作重大專項課題(No.2006DFB32210)
△通訊作者Tel:010-66939685;何昆侖E-mail:hekunlun2002@yahoo.com;王莉莉E-mail:wangll63@126.com
