蕓薹根腫菌(Plasmodiophora brassicae)單孢分離接種及生理小種的鑒定
2012-09-11 07:22:56沈向群耿新翠
植物保護 2012年3期
李 茜, 沈向群, 耿新翠, 李 林
(沈陽農業大學園藝學院,沈陽 110866)
近百年來十字花科作物的根腫病一直在不斷蔓延,成為該類作物的主要威脅[1]。蕓薹屬油用品種、大白菜、羽衣甘藍、結球甘藍、蘿卜、芥菜等均可被十字花科蔬菜根腫病菌感染[2]。分類學家已經根據分子數據將根腫病菌重新分類到新分類單元,它既不是一種真菌,也不是黏菌[3],而是一個具有兩種不同鞭毛變形蟲的原生生物多樣化群組[4]。
雖然自然界根腫病菌的種群是生理性多樣化分化型混合的狀態,但是種中也觀察到根毛侵染,皮層侵染僅發生在十字花科植物的感病植株上[4]。研究發現根腫菌生活史的大部分時期都是單倍體,發生核配后立即進行減數分裂形成次生游動孢子囊,最后形成休眠孢子[5]。可以猜測到單一的休眠孢子的后代帶有同樣的基因型。要確定這是否是同宗或異宗配合,根腫病菌單孢的研究十分必要。為了對這些病原菌的遺傳進行研究和篩選出專化抗源,獲得單孢子菌株也是非常重要的。
不同生態環境的根腫菌可以形成不同的致病性,根腫病菌也組成了一些不同的致病型,除了用病原菌根感染宿主組織和愈傷組織進行培養外,根腫病菌不能進行人工培養,所以試驗一直都較難進行[6]。因為這個原因,根腫病菌研究比其他病原菌準確性差,特別是生理小種分化型的研究,在世界上僅僅只有幾個菌株分離成功的報道。韓國是完全使用營養液培養[7],不能為根腫病菌提供理想的增殖環境;日本則是將孢子接種在無菌土壤中[8],接種成功率較低;加拿大使用瓊脂法[9],對不同接菌條件下單孢菌株進行比較,對最適合發病的土壤傳播并沒有進行試驗對比。在國內沒有純粹單孢分離生理小種的研究報道。
本文改進了單一孢子分離及接種的方法,通過進行大量接種試驗,提高了單孢感染的成功率,并對不同小種分離后致病性的不同進行了鑒定。
1 材料與方法
1.1 單孢分離材料
1.1.1 供試感病品種
大白菜主栽品種‘91-12’來自遼寧凌海,是沈陽市主栽品種,能夠被試驗用菌高度感染。
1.1.2 根腫病菌來源
根腫病菌生理小種3個,分別取自沈陽新民市、四川彭州市和四川西昌縣,通過Williams鑒別寄主鑒定,分別確定為生理小種4、生理小種7、生理小種10優勢種群。其中生理小種4為平原地區病菌,生理小種7、生理小種10為高山地區病菌。
1.2 單孢分離與接種
從被感染的大白菜根上取下根瘤組織,在培養皿中加入無菌蒸餾水搗碎。孢子懸浮液通過8層紗布過濾后離心,3000r/min離心9min,棄上清液,沉淀用蒸餾水懸浮后再4000r/min離心12min,2次重復。棄上清液,用蒸餾水懸浮沉淀物,將懸浮液的濃度調至2×103/mL 4℃保存備用[10]。單孢接菌稀釋液不可放置過夜,否則細胞會因為半透膜滲透作用,吸水漲破。單孢濃度控制在每0.5μL的液滴中含有1個單孢,孢子懸浮于滅菌的載玻片上并用蓋玻片蓋上。液滴在顯微鏡下重新使用微管吸入,反復檢查至少3次,以確保只含有一個孢子。
大白菜種子55℃溫湯浸種30min,蒸餾水沖洗后進行在顯微鏡下觀察,確保種皮表面沒有黏著根腫菌,并用無菌蒸餾水沖洗3次。將種子放在濕潤的濾紙上于25℃下培養1~2d。當開始長出根毛時將其單獨放入培養皿中,在顯微鏡下使用微管輕輕將載玻片上的單孢粘到根毛上,之后幼苗保持在25℃16h光照,20℃8h黑暗條件下,在生長箱中培養21d[11]。
1.3 單孢菌繼代擴繁
21d后將幼苗移入裝有滅菌土(pH6~6.5)的無菌苗缽中,在20~25℃的溫室中單獨培養40~50d,在流動的水中進行沖洗并確認病癥。將疑似發病的菌根放回原缽中,在菌根中間重新播入滅菌后的白菜種子,繼續培養,再培養40~50d后確認病癥。重復進行操作,直到能觀察到明顯發病為止。
單孢接種后,確認病癥的個體雖然被病原菌侵入,但還沒有形成可以滿足肉眼觀察的膨大程度,使用單孢子接種后的寄主幼苗根再接種,在最適合該菌的土壤環境下增殖。
用棕色玻璃瓶遮光臨時保存擴繁后的菌根,營養液浸根放入瓶中,用棉花封口,25℃保存3~5d。根瘤腐爛后,將其倒入到裝有無菌土的穴盤中央,并將種子播種到病根土上。
1.4 單孢后代的小種鑒定
1.4.1 Williams鑒別寄主[12]
使用Williams鑒別寄主鑒定:寄主分別為2個結球甘藍品種JQ(‘Jersey Queen’)、BS(‘Badger Shipper’),2個蕪菁甘藍品種 LT(‘Laurentian’)、WB(‘Wilhelmsburger’),4個寄主均由韓國坂田種苗公司提供。按照病菌在4個寄主上致病性的分化來確定生理小種,使用其原缽土+原缽菌根方法進行接種,避免交叉感染。
1.4.2 接種體的制備及接種方法[13]
每份擴繁后的菌株取鮮重15g進行勻漿,配制菌液100mL,利用血球計數板計算并將菌液孢子濃度稀釋為12×107個/mL,將菌液加入到300g無菌土中,攪拌均勻,密封48h以上,此時病菌孢子濃度為4×107個/g。裝入32缽穴盤中,穴盤底部封嚴,防止病菌從底部小孔互相感染。缽中心插一深3cm、直徑2cm的圓穴,置入病菌土,每穴在病菌土上播4粒種子,使每份菌土保苗(1種鑒別寄主)20株以上。采用十字花式分離栽種,可以有效避免病菌交叉感染。苗床溫度播種后控制在25℃,土壤濕度為90%,pH 控制在6~7[14]。
1.4.3 病情調查
接種后45d開始調查,根據國際通用病情分級標準[15]:0級,根系發育正常,根部沒有腫瘤;1級,1%~30%的根著生少許腫瘤,相互不連接;2級,31%~60%的根著生腫瘤,有大的腫塊,相互連接;3級,61%~100%的根著生腫瘤,紡錘形瘤著生在主根使其膨大,膨大延伸到下胚軸,植株發育矮小,生長停止。

表1 十字花科根腫病生理小種Williams鑒別系統[11]1)

2 結果與分析
2.1 單孢分離
950株幼苗接種單休眠孢子并存活,因為密度極低,在外觀上病癥極小。兩年接種擴繁后,篩選出觀察到形成菌根瘤的植物,顯微鏡檢測后成功感染單孢為27株(表2)。

表2 在2日齡大白菜根毛上單孢接種根腫病菌87d后發病情況
單孢接種后,孢子增殖是呈指數倍數增加的:開始時由于菌量很小,所以只能在主根上輕微發病。主根上出現小突起(為接種單孢擴繁后的細胞群);隨著接種時間的延長、擴繁代數的增加,孢子總數增加,植物根部病變的范圍擴大,量變增大,植株感病后因受病菌的刺激造成細胞增生,作物根部形成大小不等的腫瘤(表3)。感染的成功率低可能由于存在技術固有局限性或休眠孢子接種后存活率低。
4號和10號小種進行到第3代擴繁時,除個別植株外,大部分植株主根根瘤長到3級后就停止生長,不再膨大。表明由一個單孢接種的菌根,其病原菌的致病性與田間根腫表現出顯著差異,單孢可能由于純化后病菌活力下降導致沒有充分增殖,降低了感病能力[16]。

表3 單孢擴繁的發病情況
7號小種擴繁后3株植株外觀沒有任何感病癥狀,離心鏡檢后卻發現含有病菌。且其接種的成功率明顯比其他兩個小種低,可以看出它相對于其他兩個小種為弱勢種群,不易感染寄主。
2.2 單孢后代生理小種鑒定
接菌濃度、寄主種群數相同,但是其感染鑒別寄主的能力不同(表4)。
試驗所選擇的根腫菌生理小種4、生理小種7、生理小種10均為中國平原地區及高山地區主要的致病種群,每一種在當地都給十字花科蔬菜的種植帶來了極大的經濟損失。但是從分離出來的單孢菌株的致病性來看又有所不同。
平原地區生理小種4在東北本地傳染力極強,而其單孢分離出來的菌株也均能顯著感染鑒別寄主,包括其中分離出來的諸如生理小種2、生理小種11等單孢菌株若對該鑒別寄主沒有抗性,則一定能夠深度感染鑒別寄主,感染率多集中在70%~80%,感染力較強。
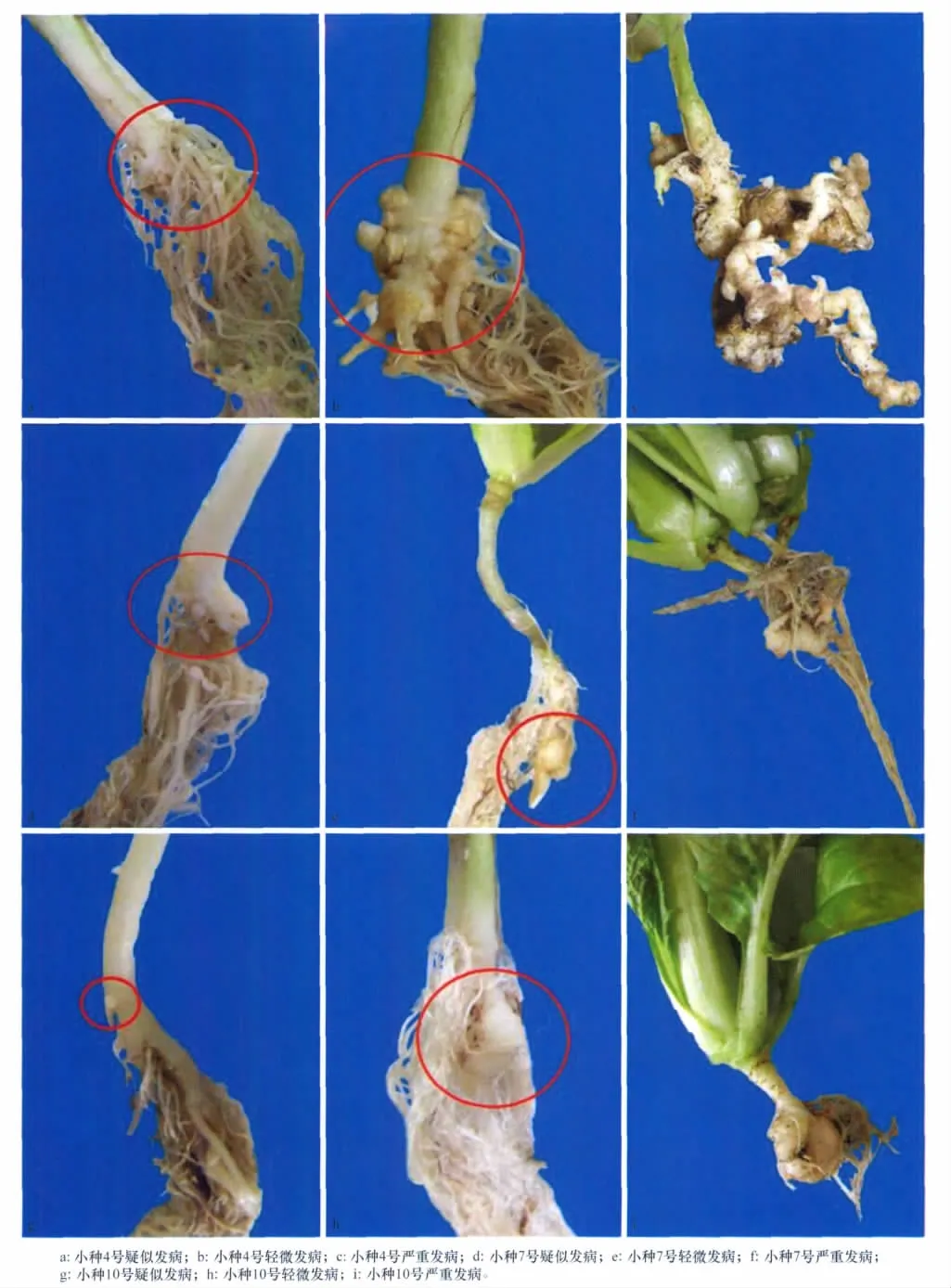
圖1 由單一的休眠孢子接種大白菜病情指數分級標準
高山地區生理小種10分離出來的10號小種,雖然為同一種群生理小種分離出來的單孢菌株,但其致病力也不同,編號9對鑒別寄主的感染力較弱,僅為其余8株小種單株感染力的一半左右;而分離出的其他小種的單孢小種12單株和小種14單株感染鑒別寄主的能力較弱,僅為30%~40%,可以看出單孢分離出來的原混生含量較少的附生菌株,致病性較差,不能大量感染鑒別寄主。
高山地區生理小種7雖然混生狀態為較強的致病菌,但是單孢分離后致病性非常弱,可見其如果要致病需要小種間協同作用,才能夠使植物大面積發病。若只有一種小種則致病力較弱。

表4 采用Williams鑒別寄主對經過繼代擴繁發病明顯的27株進行小種鑒定的結果1)
由以上鑒定結果可以發現,平原地區根腫菌和高山地區的根腫菌顯著不同。
平原地區典型生理小種4在東北本地傳染力極強,其單孢分離出來的菌株也均能顯著感染鑒別寄主,且擴繁能力較強,能夠使鑒別寄主重度感病,苗期感染3級情況較多,嚴重影響鑒別寄主的發育,重則導致絕收,對十字花科蔬菜主產區生產造成嚴重威脅[18]。因此平原地區應主要選育抗生理小種4的抗病品種。
高山地區典型生理小種7為弱勢小種,3株植株外觀沒有任何感病癥狀,離心鏡檢后卻發現含有病原菌孢子,而有明顯癥狀的植株進行Williams鑒定后沒有分離出7號小種。雖然混生狀態為較強的致病菌,但是單孢分離后致病性非常弱,需要小種間協同致病,才能夠使植物大面積發病。
高山地區另一典型生理小種10也分離出3個小種。雖然為同一種群生理小種10分離出來的10號小種單孢菌株,但其致病力卻不同。編號9對鑒別寄主的感染力僅為其余的10號小種單株的一半左右;12號小種單株和14號小種單株感染鑒別寄主的能力較弱,說明單孢分離出來的原混生含量較少的附生菌株,致病性較差,不能大量感染鑒別寄主。生理小種10對‘Laurentian’無致病性,可以考慮在高山發病地區種植蕪菁甘藍。
3 討論
蕓薹根腫菌(Plasmodiophora brassicae)生活史的大部分時期都屬單倍體[19],發生核配后立即進行減數分裂形成次生游動孢子囊,最后形成休眠孢子[20]。各小種根腫病菌孢子經倒置顯微鏡觀察,休眠孢子形狀多為圓形或近圓形,偶見不規則的橢圓形、三角形。懸浮液經觀察有1、2、4核的休眠孢子的頻率分別為89.3%、10.3%和0.3%[21]。雙核休眠孢子平均直徑(4.9μm)均大于單核休眠孢子直徑(3.5μm)。大型4核休眠孢子(直徑≥5μm)有異常不規則形狀,活性差,接種成功率極低。單核孢子接種成活率最高。
接種兩個月后,950株接種單休眠孢子的植株存活了46株,其中27株形成了菌根,成功感染率為核58.7%,明顯高于以前的研究結果[22]。單孢接種有兩個技術問題,一個是單孢子獲得:經過顯微鏡多次檢測,確保其中只含有一個孢子,增加了單孢接種的準確度[7]。二是成功接種到寄主幼苗上使用無菌土,在適合植物生長的環境下隔離培養,有利于植物發病,病菌實現穩定的增殖[8]。本研究感染非常成功可能是由于兩個原因:接種選定活性較強的單核休眠孢子,并使用微管接種使游動孢子有更多感染機會;接種后暗培養2d,使病菌附著并初步進入根毛中,使單株直立便于放入單獨容器中進行培養,25℃暗條件下經過3~5d腐爛的根瘤作為病菌來源有利于休眠孢子活化[14]。
從上述小種構成看,原始種群4中分離出2、4、8、11號4個小種,4號單株占63.6%為優勢種群,原始種群經過Williams鑒定顯示為4號。一個原始種群4,進行單孢分離后一般產生不同的其他種群,新產生的4號小種及其派生的2、8、11單孢菌株的致病性差異可能是由于不同致病型之間的相互作用。一個致病原始小種不同的孢子接種后可能會產生其他致病型,混合后可能會導致在相同的鑒定小種號存在多種致病型。10號優勢種群中分離出10、12、14號,10號單株占81.8%為優勢種群,原始種群經過Williams鑒定顯示為10號。
原始種群7單孢分離后,編號1、2單株致病性較低可能是由于其內部不同小種構成了致病型的混合物,反映了致病性菌株和非致病性菌株之間的協同作用。非致病性菌株也可能誘發抗病的宿主植物染病,從而導致植物發病。編號3、4、5雖然外表沒有感病跡象,但是它也在宿主體內增殖,宿主對它有抗性,可能是致病型和宿主基因型之間復雜的相互作用,但以何種形式相互作用,抑制發病的物質基礎是什么,仍待深入研究。
本文通過分析27株單孢菌株:非本身小種以外的9個菌株,是通過再接種也不易大量增殖的病原菌株,和被接種的3個小種是有區別的。它們雖然單獨看起來是弱勢種群,但隨著作物品種的多樣化可能會轉為新的病原型,并成為新的遺傳性感染源,或通過跟別的病原型重復感染等的相互作用,有可能引起病害蔓延。
對不同地區生理小種進行大量單孢接種,可以提高接種成功率,且較易分離出罕見生理小種,并為探明根腫菌各種不同生理小種之間是否存在致病性變異和建立生理小種鑒別體系、篩選出專化抗性品種提供依據。
[1] 徐瑞英,徐瑞芹.十字花科根腫病的發生與防治[J].吉林蔬菜,2006(2):41-42.
[2] Crete R.Distribution and importance of cruciferae growing areas and clubroot in 1974[J].Clubroot Newsl,1975,13:3.
[3] Nikolaev S I,Berney C,Fahrni J F,et al.The twilight of Heliozoa and rise of Rhizaria,an emerging supergroup of amoeboid eukaryotes[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101:8066-8071.
[4] Dixon G R.The biology of Plasmodiophora brassicae Wor.a review of recent advances[J].Acta Hortic,2006,706:271-282.
[5] Narisawax K,Kageyama K.Efficient root infection with single resting spores of Plasmodiophora brassicae[J].Mycological Research,1996,100(7):855-858.
[6] Elke Diederichsen,Martin Frauen,Linders E G A,et al.Status and perspectives of clubroot resistance breeding in crucifer crops[J].J Plant Growth Regul,2009,28:265-281.
[7] Se Jeong Jangs,Seung Hwan Heo,Chang Soon Jang,et al.Races and Dominant population of Chinese cabbage clubroot pathogen,Plasmodiophora brassicae in Korea[J].Research in Plant Disease,2007,13(1):45-49.
[8] Koji Kageyama,Yoriko Kamimura,Mitsuro Hyakumachi.A simple inoculation method with a single resting spore of Plasmodiophora brassicae[J].日植病報,1996,61:415-418.
[9] Xue S,Cao T.Isolation and variation in virulence of singlespore isolates of Plasmodiophora brassicae from Canada[J].Plant Disease,2008,92(3):456-462.
[10]楊佩文,李家瑞,楊勤忠,等.十字花科蔬菜根腫病菌休眠孢子的分離與檢測[J].云南農業大學學報,2002,17(3):301-303.
[11]F?hling M,Graf H,Siemens J.Characterization of a singlespore isolate population of Plasmodiophora brassicae resulting from a single club[J].Journal of Phytopathology,2004,152(7):438-444.
[12]Williams P H.A system for the determination of races of Plasmodiophora brassicae that infect cabbage and rutabaga[J].Phytopathology,1966,56:624-626.
[13]沈向群,聶凱,吳瓊,等.大白菜根腫病主要生理小種種群分化鑒定初報[J].中國蔬菜,2009(8):59-62.
[14]肖崇剛,郭向華.十字花科蔬菜根腫病菌的生物學特性研究[J].菌物系統,2002,21(4):597-603.
[15]孫保亞,沈向群,郭海峰,等.十字花科植物根腫病及抗病育種研究進展[J].中國蔬菜,2005(4):34-37.
[16]吉川宏昭.アブラナ科野菜の根こぶ病抵抗性育種に關する研究[J].野菜、茶業試驗場研究報告(野菜、花卉),1993,A7:61-68.
[17]唐文華.北京十字花科蔬菜根腫病的發生和鑒定[J].植物保護,1990,16(1):17-18.
[18]章元壽.植物病理生理學[M].南京:江蘇科學技術出版社,1993.
[19]呂理樂.十字花科蔬菜根腫菌[J].云南農業大學學報,2002,17(2):134-136.
[20]Larnmerink J.Physiological specialization of Plasmodiophora brassicae Wor.New Zealand[J].Agriculture Research,1964,7:37-41.
[21]Castlebury L A,Maddox J V,Glawe D A.A technique for theextraction and purification of viable Plasmodiophora brassicae resting spores from host root tissue[J].Mycologia,1994.86,458-460.
[22]Rarisawa K,Kageyama K,Hashiba T.Efficient root infection with single resting spores of Plasmodiophora brassicae[J].Mycological Research,1996,100(7):855-858.
