一株非谷氨酸依賴型聚谷氨酸產生菌的篩選和鑒定*
2012-09-12 13:22:04彭英云張濤繆銘沐萬孟江波
食品與發酵工業 2012年7期
彭英云,張濤,繆銘,沐萬孟,江波
1(江南大學食品科學與技術國家重點實驗室,江蘇無錫,214122)
2(鹽城工學院 化 學與生物工程學院,江蘇 鹽 城,224003)
一株非谷氨酸依賴型聚谷氨酸產生菌的篩選和鑒定*
彭英云1,2,張濤1,繆銘1,沐萬孟1,江波1
1(江南大學食品科學與技術國家重點實驗室,江蘇無錫,214122)
2(鹽城工學院 化 學與生物工程學院,江蘇 鹽 城,224003)
從土壤中分離到1株可不依賴谷氨酸發酵產γ-聚谷氨酸(γ-PGA)的菌株SK19.001,通過16S rDNA序列比對并結合菌株的形態、生理生化特征分析,鑒定此菌株為甲基營養芽孢桿菌(Bacillus methylotrophicus)。在以甘油和蛋白胨F403為碳源和氮源的培養基中,37℃振蕩培養24 h,γ-PGA產量可達到14.57 g/L。
γ-聚谷氨酸(γ-PGA),篩選,鑒定,甲基營養芽孢桿菌(Bacillus methylotrophicus)
γ-聚谷氨酸(Poly γ-Glutamate,γ-PGA)是一種多聚氨基酸類的環保型多功能生物可降解高分子材料,主要由D-谷氨酸和L-谷氨酸通過酰胺鍵聚合而成。作為一種高分子聚合物,γ-PGA具有一些獨特的物理、化學和生物學特性,如良好的水溶性,超強的吸附性,能徹底被生物降解,無毒無害,可食用等,可作為諸如保水劑、增稠劑、絮凝劑、重金屬吸附劑、藥物/肥料緩釋劑及藥物載體等的原料,在農業、食品、醫藥、化妝品、環保、合成纖維和涂膜等領域具有廣泛的應用前景。
γ-PGA多由微生物發酵法得到,主要菌種為地衣芽孢桿菌屬和枯草芽孢桿菌屬[1],也有報道從其它菌種如古生菌(archara)[2]、細菌(bacteria)[3]和真核生物(eucaryote)[4]發酵得到。根據細胞生長的營養要求是否需要谷氨酸,可以把γ-PGA產生菌分為兩大類:一種為谷氨酸依賴型[5],如Bacillus licheniformis ATCC9945[6],B.subtilis IFO3335[7],B.subtilis F-2-01[8]等,另一種為非谷氨酸依賴型,如B.subtilis TAM-4[9],B.licheniformis A35[10]。國內學術界對前者的研究較多,有的菌株γ-PGA的產量隨著培養基中谷氨酸含量的增加而增加[11],有的則需要有谷氨酸作為啟動因子才會有γ-PGA的產生[12]。而對于非谷氨酸依賴型γ-PGA產生菌報道較少,從工業化角度考慮,篩選非谷氨酸依賴型γ-PGA產生菌株,利用廉價原料生產此種生物可降解高分子材料是非常必要的。
1 材料和方法
1.1 材料
1.1.1 樣品來源
土壤。
1.1.2 培養基
斜面培養基(g/L):牛肉膏5,蛋白胨5,NaCl,瓊脂15,pH7.0~7.2,37℃。
種子培養基(g/L):葡萄糖20,酵母提取粉25,MgSO4·7H2O 0.5,K2HPO41,pH 7.0~7.2,37℃,200 r/min 12 h。
篩選培養基(g/L):種子培養基+瓊脂15。
發酵培養基(g/L):葡萄糖15,蛋白胨15,酵母提取粉25,MgSO4·7H2O 0.6,K2HPO41,MnSO41 mmol/L,pH 7.0~7.2,37℃,200 r/min,12 h。
1.2 方法
1.2.1 篩選方法
土壤樣品以無菌去離子水浸泡,稀釋液涂布于篩選培養基固體平板,37℃培養24 h,觀察菌落生長情況,挑選生長快,高黏度的菌落做進一步的篩選分離,將分離到的菌落接種到發酵培養基中,37℃,200 r/min培養24 h,測定發酵液中γ-PGA的含量。
1.2.2 分析方法
1.2.2.1 細胞生長的測定
將培養液稀釋20倍后用752分光光度計讀取660 nm處吸光值A(660)。
1.2.2.2 發酵液黏度的測定
采用SNB-2數字式黏度儀室溫下測定發酵液的黏度。
1.2.2.3 γ-PGA的分離純化
發酵液適當稀釋后以10000 r/min,離心25 min去除菌體,制得發酵上清液,加入2~4倍體積的無水乙醇沉淀聚谷氨酸,輕微攪拌后于4℃靜置過夜,將聚谷氨酸沉淀以少量無水乙醇洗滌,用少量去離子水溶解,再次離心去除不溶物,經透析除去鹽、糖等小分子,最后冷凍干燥,得到γ-聚谷氨酸。
1.2.2.4 γ-PGA的定性
通過水解液薄層層析法(TLC)和高效液相色譜法(HPLC)分析,將純化后的聚谷氨酸以6 mol/L HCl,110℃水解24 h,水解液以6 mol/L NaOH中和后以谷氨酸為對照分別用薄層層析法和高效液相色譜法分析其成分。
1.2.2.5 γ-PGA的測定
采用高效液相色譜法(HPLC)分析:將發酵液離心去除菌體后,取一定量以6 mol/L HCl,110℃水解過夜,分別用HPLC測定水解液和發酵液中游離谷氨酸的含量,兩者之差即為γ-PGA的產量。
1.2.2.6 菌種的鑒定與保藏
委托武漢大學中國典型培養物保藏中心(CCTCC)進行鑒定與保藏。
1.2.3 不同碳、氮源對γ-PGA產量的影響
在其它發酵條件相同的情況下,分別選取不同的碳源和氮源進行培養,測定發酵液中γ-PGA的含量,確定SK19.001對碳源和氮源的利用情況。
2 結果與分析
2.1 SK19.001菌株的鑒定
2.1.1 形態特征
通過篩選和分離,從土壤樣品中得到1株產γ-聚谷氨酸的高黏菌落,編號為SK19.001,保藏于中國典型培養物保藏中心(保藏號為CCTCC NO:M2011201)。SK19.001菌株在固體培養基上的生長形態有多樣性,當培養基潮濕時,菌落濕潤、不透明,表面光滑呈“木耳狀”,中心有高黏液滴(圖1);干燥時,菌落粗糙皺褶,中央凹陷,邊緣不規則。

圖1 SK19.001的菌落形態
2.1.2 生理生化特征(表1)

表1 菌株SK19.001生理生化特性
2.1.3 遺傳學特征
經過16S rRNA基因序列分析并將序列上傳Genbank做BLAST比較(基因登錄號為JQ723479),利用Mega4.1軟件構建進化樹(圖2),其中同源性最高的為菌株Bacillus methylotrophicus P07(gb|JN700153.1),同源性為100%,結合生理生化特征,將菌株SK19.001鑒定為甲基營養芽孢桿菌(Bacillus methylotrophicus)。
2.2 發酵水解產物的分析
按照1.2.2.4的方法對發酵和發酵水解產物進行TLC分析,以谷氨酸為對照,結果見圖3。從圖3可以看出,發酵產物水解后主要成分為谷氨酸,進一步通過高效液相色譜分析發現,水解產物出峰單一且出峰時間與谷氨酸一致(圖4和圖5),由此可以斷定發酵產物為聚谷氨酸。
2.3 對碳源的利用
分別選取葡萄糖、甘油、谷氨酸鈉、可溶性淀粉、α-乳糖、麥芽糖、蔗糖、檸檬酸、果糖為碳源,濃度各為30 g/L,其余培養基成分為(g/L):酵母提取粉50、MgSO4·7H2O 0.6、K2HPO41、MnSO41 mmol/L,pH 7.0~7.2,37℃,200 r/min,24 h。結果見表2。

圖2 SK19.001菌株的系統發育進化樹

圖3 發酵水解產物層析圖
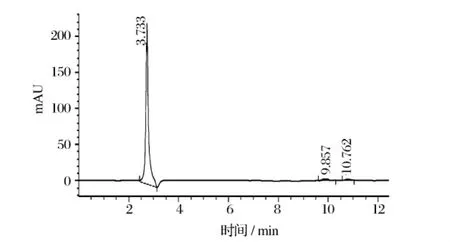
圖4 谷氨酸標樣的HPLC圖譜

圖5 發酵水解產物的HPLC圖譜

表2 碳源對SK19.001產γ-PGA的影響
從表2中可以看出,與其它聚谷氨酸產生菌不同的是,甲基營養芽孢桿菌SK19.001可利用的碳源非常廣泛,對選取的每種碳源都可利用并產生PGA,說明它是1株可不依賴谷氨酸發酵產聚谷氨酸的菌株,在已經報道的文獻中,除了B.subtilis TAM-4[9],B.licheniformis A35[10]以外,其余產聚谷氨酸的菌株大都需要谷氨酸的存在,如Jung等人[11]對1株Bacillus sp.RKY3的研究表明,培養基如不添加谷氨酸則沒有γ-PGA的生成。
2.4 對氮源的利用
分別選取魚粉蛋白胨、胰蛋白胨、酪蛋白、酵母提取粉、蛋白胨(動物)、蛋白胨F403、玉米漿粉、大豆蛋白胨、NH4Cl、(NH4)2SO4、NH4NO3、脲素作為氮源,含氮量為5 g/L,其余培養基為(g/L):甘油30,MgSO4·7H2O 0.6,K2HPO41,MnSO41 mmol/L,pH 7.0~7.2,37℃,200 r/min,24 h。結果見表3。
從表3可以看出,甲基營養芽孢桿菌SK19.001主要利用有機氮源產聚谷氨酸,無機氮源幾乎不利用,有別于B.subtilis TAM-4[9]、B.subtilis PGA-O-7[13]可利用無機氮源產γ-PGA;在有機氮源中,酵母提取粉、玉米漿粉、蛋白胨(動物)和蛋白胨F403產γ-PGA較高,而不同來源的蛋白胨對SK19.001產γ-PGA的影響較大,表現為對魚粉蛋白胨幾乎不利用;而對動物蛋白胨和蛋白胨F403利用率較高。此外,對γ-PGA黏度的測定表明,發酵液黏度與產量有一定的相關性,即有γ-PGA產生的發酵液都具有一定的黏度,但并不是黏度越高產量越高,有時可能因為發酵液氣泡的影響而使得黏度測定值偏高。

表3 氮源對SK19.001產γ-PGA的影響
3 結果與討論
通過篩選和鑒定,從土壤中得到1株可不依賴谷氨酸發酵產γ-PGA的甲基營養芽孢桿菌(Bacillus methylotrophicus),這是除枯草芽孢桿菌、地衣芽孢桿菌和淀粉液化芽孢桿菌[14]外的1株新的產γ-PGA的芽孢桿菌種屬。通過對碳源和氮源的篩選,SK19.001對碳源的利用范圍非常廣泛,而對氮源則有一定的選擇性。
國內外對谷氨酸依賴型菌株研究較多,但從生產原料角度考慮,不需谷氨酸作為發酵底物的菌株具有成本上的優勢,更具有工業化應用前景。目前國內對此類菌株的研究還較少,產量也不高(Bacillus subtilis PGA-O-7:2.8 mg/mL[13];Bacillus licheniformis PGAN:11 mg/mL[15];Bacillus amyloliquefaciens LL3:4.36 g/L[14]),SK19.001在碳氮源簡單篩選的情況下可產出γ-PGA 14.57 g/L,因此如進一步優化發酵條件,研究代謝過程中各種因素對γ-PGA積累的影響,γ-PGA的產出水平還有較大的提高空間。
[1]Joerg M Buescher.Microbial Biosynthesis of Polyglutamic Acid Biopolymer and Applications in the Biopharmaceutical,Biomedical and Food Industries[J].Critical Reviews in Biotechnol,2007,27(1):1-19.
[2]Hezayen F F,Rehmb H A,Eberhardt R,et al.Polymer production by two newly isolated extremely halophilic archaea:application of a novel corrosion-resistant bioreactor[J].Applied Microbiology Biotechnology,2000,54(3):319-325.
[3]Ashiuehi M,Soda K,Misono H A.Poly-γ-glutamate synthetic system of Bacillus subtilis IFO3336:gene cloning and biochemical analysisofpoly-γ-glutamateproduced by Escherichia coli clone cells[J].Biochemical Biophysical Research Communications,1999,263(1):6-12.
[4]Kocianova S,Vuong C,Yao,Y F,et al.Key role of polygamma-DL-glutamic acid in immune evasion and virulence of Staphylococcus epidermidis[J].Journal of Clinical Investigation,2005,115(3):688-694.
[5]Shih I L and Van Y T.The production of poly-(γ-glutamic acid)from microorganisms and its various applications[J].Bioresource Technology,2001,79(3):207-225.
[6]Troy F A.Chemistry and biosynthesis of the poly(γ-D-glutamyl)capsule in Bacillus licheniformis and properties of the membrane-mediated biosynthetic reaction[J].Journal of Biological Chemistry,1973,248(1):305-316.
[7]Kunioka M.Biosynthesis of poly(γ-glutamic acid)from L-glutamine,citric acid and ammonium sulfate in Bacillus subtilis IFO 3335[J].Applied Microbiology Biotechnology,1995,44(3):501-506.
[8]Kunioka M,Goto A.Biosynthesis of poly(γ-glutamic acid)from L-glutamic acid,citric acid,and ammoniumsulfate in Bacillus subtilis IFO 3335[J].Applied Microbiology Biotechnology,1994,40(6):867-872.
[9]Ito Y,Tanaka T,Ohmachi T,et al.Glutamicacid independent production of poly(γ-glutamicacid)by Bacillus subtilis TAM-4[J].Bioscience Biotechnology and Biochemistry,1996,60(8):1239-1242.
[10]Cheng C,Asada Y,Aaida T.Production of γ-polyglutamic acid by Bacillus subtilis A35 under denitrifyingconditions[J].Agricultural Biological and Chemistry,1989,53(9):2369-2375.
[11]Jung D Y,Jung S,Yun J S.Influence of Cultural Medium Compoment on the Production of Poly(γ-glutamic acid)by Bacillus sp.RKY3[J].Biotecchnology and Bioprocess Engineering,2005,10:289-295.
[12]Ko Y H,Gross R A.Effects of Glucose and Glycerol on γ-Poly(glutamic acid)formation by Bacillus licheniformis ATCC 9945a[J].Biotechnology and Bioengineering,1997,57(4):430-437.
[13]施慶珊,李誠斌,王春華,等.一株不需谷氨酸的產聚γ-谷氨酸菌株的篩選與鑒定[J].微生物學通報,2007,34(2):307-311.
[14]Cao M F,Geng W T,Liu L,et al.Glutamic acid independent production of poly-γ-glutamic acid by Bacillus amyloliquefaciens LL3 and cloning of pgsBCA genes[J].Bioresource Technology,2011,102(5):4251-4257.
[15]疏秀林,施慶珊,馮靜,等.一株非谷氨酸依賴型聚γ-谷氨酸高產菌株的鑒定與誘變育種[J].微生物學通報,2009,36(5):705-710
ABSTRACTA strain SK19.001 which could produce poly γ-glutamate(γ-PGA)independent of glutamic acid in culture was isolated.It was identified as Bacillus methylotrophicus based on the sequence analysis of 16S rDNA and a series of morphological and biochemical characteristics.The yield of γ-PGA was 14.57 g/L when the strain was grown in glycerol and peptone F403-containing medium at 37℃for 24 h with shaking.
Key wordspoly γ-glutamate(γ-PGA),screening,identification,Bacillus methylotrophicus
Screening and Identification of a Glutamate-independent Poly γ-glutamate Produced Strain
Peng Ying-yun1,2,Zhang Tao1,Miao Ming1,Mu Wan-meng1,Jiang Bo1
1(State Key Laboratory of Food Science and Technology,Jiangnan University,Wuxi 214122,China)
2(School of Chemical and Biological Engineering,Yancheng Institute of Technology,Yancheng 224003,China)
在讀博士(江波教授為通訊作者)。
*國家自然科學基金項目(20976073,31000764);江蘇省國際合作項目(BZ2011026)。
2012-03-20,改回日期:2012-05-22
