生育三烯酚生物合成與生理功能研究進展
2012-11-10 06:16:46范麗萍靳雅欣趙福永長江大學生命科學學院湖北荊州434025
長江大學學報(自科版) 2012年11期
關鍵詞:植物
范麗萍,靳雅欣,趙福永 (長江大學生命科學學院,湖北 荊州 434025)
生育三烯酚生物合成與生理功能研究進展
范麗萍,靳雅欣,趙福永 (長江大學生命科學學院,湖北 荊州 434025)
生育三烯酚是天然維生素E的重要組成物質,包含α、β、γ、δ-生育三烯酚4種同系物,僅在植物中有合成。生育三烯酚因具有多不飽和側鏈不僅是一種優良的抗氧化劑,而且還具有抗癌、防治心腦血管疾病等重要生理功能。近幾年來,對生育三烯酚的生物合成及生理作用進行了廣泛地研究,就這些研究成果進行了一個階段性歸納總結,并就生育三烯酚的開發與利用等問題進行了討論。
維生素E;生育三烯酚;生物合成;生理功能
維生素E(Vitamin E,VE)是1922年由Evans和Bishop最先發現的能夠顯著提高老鼠繁殖力的一種“致育因子”,后被命名為“生育酚”。天然VE是一種脂溶性抗氧化劑,包含生育酚(tocopherol)和生育三烯酚(tocotrienol)2大類,每類均由α、β、γ、δ4種同系物組成[1]。天然VE只能由光合自養植物合成,廣泛存在于植物的綠色組織和含油種子中[2-4]。VE特有的理化性質使其具有多種生理功能,可增強動物繁殖能力、提高機體免疫能力、防治心腦血管疾病、糖尿病和癌癥等,作為抗氧化劑和營養補充劑廣泛應用于醫藥、保健品、食品、化妝品和飼料等行業[5]。同時,植物自身合成的VE在維持其膜系統的穩定性、光合電子傳遞、胞間信號轉導與轉錄調控等方面也承擔著重要角色,高水平表達可顯著提高其抗逆能力[6-9]。鑒于天然VE的重要生理功能和廣闊的開發應用前景,20世紀中葉就對生育酚的生物合成、代謝及其生理作用機制等方面開展了廣泛的研究,尤其是對生物活性最高的α-生育酚研究得最為深入[10]。直到20世紀80年代末,生育三烯酚的降低膽固醇和抗癌特性的發現才逐漸引起研究者的重視。近幾年來,生育三烯酚的非抗氧化性生理功能得到了廣泛地發掘和研究,為此,筆者就這些研究成果進行了一個階段性的歸納和總結。
1 生育三烯酚的生物合成與來源
天然VE是通過一系列的酶促反應在葉綠體內膜合成的[11],合成途徑受多個關鍵酶基因(VTE1、VTE2、VTE3、VTE4、VTE5)調控(圖1)[12]。生育三烯酚和生育酚的合成均是通過非甲羥戊酸途徑和酪氨酸降解途徑來獲得合成前體物。在單子葉植物中,尿黑酸牻牛兒基牻牛兒基轉移酶(HGGT)牻牛兒基牻牛兒基二磷酸(GGDP)和尿黑酸(HGA)為底物合成生育三烯酚前體物2-甲基-6-牻牛兒基牻牛兒基苯醌(MGGBQ),進而進入生育三烯酚合成途徑;而在雙子葉植物中,尿黑酸植基轉移酶(HPT)可以植基二磷酸(PDP)和尿黑酸為底物催化合成生育酚前體物2-甲基-6-植基苯醌(MPBQ)。研究表明,盡管單子葉來源的HGGT和雙子葉來源的HPT在氨基酸序列上存在40~50%的相似性,但是它們對底物的親和力卻有顯著差別,HGGT與GGDP的親和力是其與PDP的5倍,而HPT對PDP的親和力則是其與GGDP的50~80倍[12]。
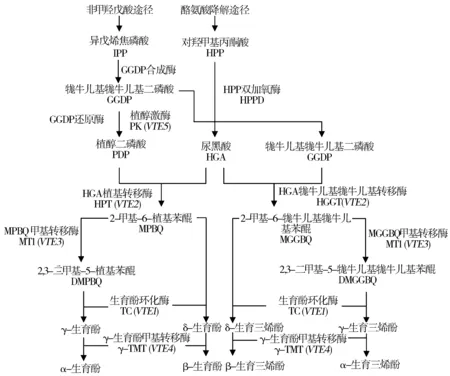
圖1 植物維生素E生物合成途徑
天然VE廣泛地存在于油菜、玉米和大豆等各種油料作物的種子中。研究發現,油菜籽中不論是VE的總含量還是α-生育酚的含量均要高于玉米和大豆,平均含量分別達到了109、42.61mg/100g[3]。核桃仁中的VE含量也比較高,高效液相色譜分析表明,其總含量在8.51~35.28mg/100g之間,α-生育酚含量偏低,僅為0.27~1.53mg/100g[2]。說明未經精煉的植物油是一種人類VE補充的良好膳食來源。
生育三烯酚往往在單子葉植物種子及其植物油中含量較高,如大麥(910mg/kg)、棕櫚油(738mg/kg)、米糠(465mg/kg)、燕麥(210mg/kg)、小麥胚(189mg/kg)、椰果(25mg/kg)等,且各種植物中各種生育三烯酚同系物的成份和相對含量也不一致,棕櫚油中只含有α、γ、δ-生育三烯酚3種,且γ-生育三烯酚含量最高,達(439mg/kg),而米糠中僅含α、γ-生育三烯酚2種[13]。
2 生育三烯酚的生理功能
Matringe等[9]研究了生育三烯酚在植物中的生理作用。結果發現,生育三烯酚在轉HPPD-PDH煙草的幼葉中有積累,并且在低溫強光脅迫下可以將光系統Ⅱ的光合抑制和質體膜的過氧化反應維持在一個較低水平,說明生育三烯酚在植物體內可以作為質體膜過氧化反應的有效抗氧化劑。與生育酚不同,生育三烯酚因在側鏈上具有3個不飽和雙鍵,有高于生育酚50倍的抗氧化力,對皮膚細胞膜的浸透力也比生育酚要高15倍。4種生育三烯酚同系物中生理活性最高的是δ-生育三烯酚[12]。而生育酚當中,生理活性最高的是α-生育酚,其活性分別是β、γ、δ-生育酚的2倍、10倍、33倍,其生理功能也是最為清楚的[10]。體外條件下,相同濃度的生育三烯酚、α-生育酚和γ-生育酚在質體氧化過程中清除氧自由基的能力相當。但是體內氧自由基的清除主要是α-生育酚,而非γ-生育酚,主要是α-生育酚在組織中的含量是γ-生育酚的10倍左右[14]。人類和高等哺乳動物從食物中攝取的維生素E在小腸內被吸收,然后經過淋巴系統進入體內循環,最后以乳糜微滴形式被轉運致肝臟[15]。維生素E是由細胞色素P450進行代謝并最終由尿液排除體外[16]。
2.1 α-生育三烯酚的功能
α-生育三烯酚在色滿環R1、R2、R3 3個位置均含有甲基,是在γ-生育三烯酚基礎上合成的產物,在大麥中含量最高,達670mg/kg,其最重要的生理功能在于神經保護作用。
中樞神經系統腦組織中富含花生四烯酸等多種多不飽和脂肪酸,花生四烯酸的氧化不穩定性會引起大腦神經細胞的病變。棕櫚油中的α-生育三烯酚能夠通過酶代謝途徑和非酶代謝途徑減緩花生四烯酸的氧化進程[17]。谷氨酸鹽可激活pp60(c-Src)和12-脂氧合酶(12-lipoxygenase,12-Lox)的表達,是導致神經細胞死亡的重要因素。藥理學和遺傳學研究表明,12-Lox的酪氨酸在c-Src存在的條件下會快速磷酸化,這種磷酸化對c-Src的抑制因子及α-生育三烯酚非常敏感,納摩爾濃度的α-生育三烯酚能起到很好的抑制作用。α-生育三烯酚也可直接結合12-Lox及磷酸化的12-Lox而影響其催化活性,說明神經細胞胞質中的c-Src和12-Lox是α-生育三烯酚的作用靶標,α-生育三烯酚可以通過其抗氧化作用和非抗氧化功能起到保護神經的作用[18]。此外,α-生育三烯酚還具有阻止血小板血栓形成和血小板凝集的作用[19]。
2.2 β-生育三烯酚的功能
β-生育三烯酚在色滿環R1、R3 2個位置含有甲基,是在δ-生育三烯酚基礎上合成的產物,在小麥中含量較高,達33~43mg/kg[20]。目前對其生理功能的認識還很膚淺,僅發現其抑制小牛動脈內皮細胞增殖與微管形成活性高于γ-生育三烯酚和α-生育三烯酚[21]。
2.3 γ-生育三烯酚的功能
γ-生育三烯酚在色滿環R2、R3 2個位置含有甲基,是生育三烯酚合成途徑中第一個合成的產物,在棕櫚油中含量較高,其主要生理功能表現在抗癌和抑制細胞衰老。
徐偉麗等[22]研究發現,γ-生育三烯酚對結腸癌細胞株HT-29具有很強的細胞毒作用,可導致癌細胞中DNA斷裂損傷,且與濃度呈一定相關性。孫文廣等[23]也證明,γ-生育三烯酚人胃癌SGC-7901細胞生長具有抑制作用,具有劑量和時間效應,推測可能是γ-生育三烯酚導致癌細胞中DNA斷裂損傷而誘導的細胞凋亡有關。Makpol等[24]選取了不同年齡(21歲、40歲、68歲)人的成纖維原細胞用H2O2處理產生氧化脅迫,對處理前、后的細胞再用一定濃度(40μmol/L或80μmol/L)的γ-生育三烯酚進行再處理。結果表明,γ-生育三烯酚可以抑制人成纖維原細胞的衰老,主要是通過抗氧化作用顯著地阻止端粒長度變短和端粒酶活性降低。相對年幼細胞而言,衰老的人成纖維原細胞中CCND1、RB1、MMP1和IL6基因的表達水平提高。將其與γ-生育三烯酚共培養24h后,衰老的人成纖維原細胞中CCND1、RB1的表達水平會顯著降低,且處于G0/G1期的細胞群體減少,而處于G2/M期的細胞群體增加。不論在衰老細胞還是年輕細胞,γ-生育三烯酚均會引起與衰老相關基因表達水平的變化,表明γ-生育三烯酚可以通過調節細胞周期和與衰老相關基因的表達來延緩細胞的衰老[25-26]。
一定劑量的γ-生育三烯酚與其他化學抗癌藥物聯用可以對乳腺癌細胞的生長與活力起到很好的抑制作用[27-28]。抑制素是3-hydroxy-3-methylglutaryl-coenzyme A(HMGCoA)還原酶的優良抑制劑,是一種良好的抗癌化學藥物,但是具有較大毒性。厄洛替尼和吉非替尼也是一種抗癌良藥,它可抑制HER/ErbB受體亞型的活性,但是癌細胞中不同EGF受體之間可以形成異源二聚體,只針對其中1個亞型也起不到良好的殺滅作用。低劑量的γ-生育三烯酚與Celecoxib聯用可以通過依賴和不依賴COX-2兩種機制具有協同抗癌作用,前者主要是抑制了PGE2的表達,而后者主要是減少了ErbB2-4受體表達量[29]。
2.4 δ-生育三烯的功能
δ-生育三烯僅在色滿環R3位置含有甲基,其主要生理功能在于抗癌和抑制腫瘤細胞生長作用。
李大鳴等[30]研究發現,一定濃度(gt;30μmol/L)的δ-生育三烯酚對結腸癌SW620細胞的生長具有明顯的抑制作用,推測可能是δ-生育三烯酚上調了癌細胞中軸蛋白(Axin)的表達而實現抑制作用的。δ-三烯生育酚還能通過調控Bax、Bcl-2等細胞凋亡相關基因的表達抑制鼻咽癌5-8F細胞的增殖并誘導其凋亡[31]。δ-生育三烯還具有輻射防護作用,其機制可能在于δ-生育三烯可以激活胞外信號傳導相關激酶的磷酸化而阻止DNA損傷的產生。δ-生育三烯可以促進rapamycin靶標蛋白的表達以及其下游效應因子4EBP-1的磷酸化,這些均與負責細胞存活與生長的核糖體蛋白S6和翻譯調控因子eIF4E轉錄激活有關[32]。
免疫系統功能的變化導致的多種與年齡相關的疾病的發生與體內一氧化氮的增加密切相關。δ-三烯生育酚可抑制由脂多糖誘導的與衰老和炎癥相關基因(IL-1b、IL-1a、IL-6、TNF-a、IL-12、iNOS、VCAM1、ICAM1、COX2、IL-1RA、TRAF1、CD40)的表達,從而抑制一氧化氮的產生[33-34]。生育三烯酚可以通過下調內表皮細胞VEGF受體的表達而減少VEGF誘導的血管的形成,進而抑制腫瘤細胞的生長[35]。生育三烯酚在納摩爾濃度具有保護神經的作用,在毫摩爾濃度具有抗癌作用。Husain等[36]研究發現,將小鼠100mg/kg劑量口飼δ-生育三烯酚,連續飼喂6周后,胰腺癌組織中的δ-生育三烯酚含量可達(41.8±3.5)nmol/g,可以抑制80%的癌細胞生長。而胰腺中的含量是癌組織中的10倍,沒有表現出組織病理變異和毒性。
4 展望
提高植物維生素E含量不僅可以擴大人類和高等哺乳動物天然維生素E的攝取來源,而且還可以提高植物自身的抗逆性,加之維生素E表現出來的重要生理功能,因此如何提高植物中的維生素E含量近來已被植物育種家們提上議事日程。自然界中很多植物本身的維生素E含量就較高,因此可以以這些材料為親本進行雜交育種,從而選育出高維生素E含量的品種;但是,有些研究[2-4、37-39]表明,維生素E含量的遺傳受環境因素影響很大,這給采用常規育種途徑培育高維生素E含量的品種帶來了一定的困難。然而,目前對于維生素E的生物合成途徑已比較清楚,多個關鍵酶合成基因也已被克隆,因此,采用基因工程手段來提高植物中的維生素E含量也不失為一條理想途徑。然而,近十年來采用基因工程手段來提高花生、玉米、油菜等油料作物種子中的維生素E含量的報道屢見不鮮[40-45],但是真正實現商業化生產的品種卻沒有,可見優良的基因資源仍是制約這一技術的瓶頸。
盡管生育三烯酚體外抗氧化能力是生育酚的50多倍,且具有預防動脈硬化、風濕性關節炎、心臟疾患、腦梗塞、癌癥、糖尿病性視網膜癥、血管新生病、延緩細胞衰老等重要生理作用,但是人類和高等哺乳動物肝臟中的生育酚結合蛋白(tocopherol-binding protein,TBP)僅對α-生育酚具有高度的專一性,使α-生育酚總是被優先吸收和利用,而生育三烯酚很難被吸收而發揮功效,因此如何促進生育三烯酚的吸收和利用也是一個值得研究的問題。生育三烯酚被認為是開發抗癌疫苗的良好佐劑,與其他抗癌藥物聯用表現出了良好的抗癌效果[46-48],但是對其分子機制目前還沒有深入的認識,廣泛開展這方面的研究也是深入利用生育三烯酚的一個重要方面。
[1]Kamal-Eldin A,Appelqvist LA.The chemistry and antioxidant properties of tocopherols and tocotrienols [J].Lipids,1996,31:671-701.
[2]孫 翠.核 桃(JuglansregiaL.)堅果仁維生素 E含量分析及jrMPBQ基因克隆 [D].青島:山東農業大學.2010.
[3]李祿慧,徐妙云,張 蘭,等.不同作物中維生素E含量的測定和比較 [J].中國農學通報,2011,27(26):124-128.
[4]Corzo-Valladaresa P A,Fernández-Martínezb J M,Velascob L.Tocochromanol content and composition inJatrophacurcasseeds [J].Industrial Crops and Products,2012,36:304-307.
[5]Brigelius-Flohe R,Traber M G.Vitamin E:function and metabolism [J].FASEB J,1999,13:1145-1155.
[6]Havaux M,Eymery F,Porfirova S,et al.vitamin E protects against photoinhibition and photooxidative stress inArabidopsisthaliana[J].Plant Cell,2005,17:3451-3469.
[7]Kanwischer M,Porfirova S,Bergmüller E,et al.Alterations in tocopherol cyclase activity in transgenic and mutant plants ofArabidopsisaffect tocopherol content,tocopherol composition,and oxidative stress [J].Plant Physiol,2005,137:713-723.
[8]郭 娟,劉小麗,李曉峰,等.擬南芥VTE1過量表達可以增加維生素E含量和提高煙草植株耐鹽性[J].應用與環境生物學報,2006,12(4):468-471.
[9]Matringe M,Ksas B,Rey P,et al.Tocotrienols,the unsaturated forms of vitamin E,can function as antioxidants and lipid protectors in tobacco leaves [J].Plant Physiol,2008,147:764-778..
[10]Engin K N.Alpha-tocopherol:looking beyond an antioxidant [J].Molecular Vision,2009,15:855-860.
[11]Soll J,Douce R,Schultz G.Site of biosynthesis of α-tocopherol in spinach chloroplasts [J].FEBS Lett,1980,112:243-246.
[12]Rebeiz C A,Benning C,Bohnert H J,et al.Advances in Photosynthesis and Respiration(Volume 31):The Chloroplast:Basics and Applications[M].Dordrecht:Springer,2010.
[13]http://www.tocotrienol.org/index.php?option=com_contentamp;view=articleamp;id=49amp;itemid=59.
[14]Wolf G.How an increased intake of alpha-tocopherol can suppress the bioavailability of gamma-tocopherol [J].Nutr Rev,2006,64:295-299.
[15]Fairus S,Nor R M,Cheng H M,et al.Alpha-tocotrienol is the most abundant tocotrienol isomer circulated in plasma and lipoproteins after postprandial tocotrienol-rich vitamin E supplementation [J].Nutrition Journal,2012,11:5.
[16]Brigelius-Flohé R.Vitamin E and drug metabolism [J].Biochem Biophys Res Commun,2003,305:737-740.
[17]Sen C K,Rink C,Khanna S.Palm oil-derived natural vitamin E α-tocotrienol in brain health and disease [J].J Am Coll Nutr,2010,29(3 Suppl):314S-323S.
[18]Khanna S,Parinandi N L,Kotha S R,et al.Nanomolar vitamin E α-tocotrienol inhibits lutamate-induced activation of phospholipase A2 and causes neuroprotection [J].J Neurochem,2010,112:1249-1260.
[19]Qureshi A A,Karpen C W,Qureshi N,et al.Tocotrienols-induced inhibition of platelet thrombus formation and platelet aggregation in stenosed canine coronary arteries [J].Lipids in Health and Disease,2011,10:58.
[20]Panfili G,Fratianni A,Irano M.Normal phase high-performance liquid chromatography method for the determination of tocopherols and tocotrienols in cereals [J].J Agric Food Chem,2003,51:3940-3944.
[21]Miyazawa T,Tsuzuki T,Nakagawa K,et al.Antiangiogenic potency of vitamin E [J].Ann N Y Acad Sci,2004,1031:401-404.
[22]徐偉麗,陳炳卿,王 琪,等.γ-生育三烯酚對人結腸癌HT-29細胞中DNA 損傷作用的研究 [J].疾病控制雜志,2007,11(6):603-606.
[23]孫文廣,劉慧坤,喬 羽,等.γ-生育三烯酚對人胃癌SGC-7901細胞的抑制作用及機制 [J].現代腫瘤醫學,2012,20(2):230-233.
[24]Makpol S,Abidin A Z,Sairin K,et al.γ-Tocotrienol prevents oxidative stress-induced telomere shortening in human fibroblasts derived from different aged individuals [J].Oxidative Medicine and Cellular Longevity,2010,3:35-43.
[25]Makpol S,Zainuddin A,Chua K H,et al.Gamma-tocotrienol modulation of senescence-associated gene expression prevents cellular aging in human diploid fibroblasts [J].Clinics,2012,67:135-143.
[26]Catalgol B,Ozer N K.Protective effects of vitamin E against hypercholesterolemia-induced age-related diseases [J].Genes Nutr,2012,7:91-98.
[27]Sylvester P W.Synergistic anticancer effects of combined c-tocotrienol with statin or receptor tyrosine kinase inhibitor treatment [J].Genes Nutr,2012,7:63-74.
[28]Kannappan R,Gupta S C,Kim J H,et al.Tocotrienols fight cancer by targeting multiple cell signaling pathways [J].Genes Nutr,2012,7:43-52.
[29]Shirode A B,Sylvester P W.Mechanisms mediating the synergistic anticancer effects of combined γ-tocotrienol and Celecoxib treatment [J].J Bioanal Biomed,2011,3:1-7.
[30]李大鳴,邸金茹,王春花,等.δ-生育三烯酚對人結腸癌SW620細胞抑制作用 [J].中國公共衛生,2010,26(9):1136-1137.
[31]曾 攀.d-δ-三烯生育酚對鼻咽癌細胞增殖與凋亡的影響及分子機制 [D].桂林:桂林醫學院,2011.
[32]Li X H,Fu D,Latif N H,et al.δ-tocotrienol protects mouse and human hematopoietic progenitors from γ-irradiation through extracellular signal-regulated kinase/mammalian target of rapamycin signaling [J].Haematologica,2010,95:1996-2004.
[33]Qureshi A A,Tan X Y,Reis J C,et al.Suppression of nitric oxide induction and pro-inflammatory cytokines by novel proteasome inhibitors in various experimental models [J].Lipids in Health and Disease,2011,10:177.
[34]Qureshi A A,Tan X Y,Reis J C,et al.Inhibition of nitric oxide in LPS-stimulated macrophages of young and senescent mice by δ-tocotrienol and quercetin [J].Lipids in Health and Disease,2011,10:239.
[35]Wells S R,Jennings M H,Rome C,et al.Alpha-,gamma and delta-tocopherols reduce inflammatory angiogenesis in human microvascular endothelial cells [J].J Nutr Biochem,2010,21:589-597.
[36]Husain K,Francois R A,Hutchinson S Z,et al.Vitamin E δ-tocotrienol levels in tumor and pancreatic tissue of mice after oral administration [J].Pharmacology,2009,83:157-163.
[37]Li H Y,Liu H C,Han Y P,et al.Identification of QTL underlying vitamin E contents in soybean seed among multiple environments [J].Theor Appl Genet,2010,120:1405-1413.
[38]Marwede V,Schierholt A,Mollers C,et al.Genotype×environment interactions and heritability of tocopherol contents in canola [J].Crop Sci,2004,44:728-731.
[39]Almeida J,Quadrana L,Asís R,et al.Genetic dissection of vitamin E biosynthesis in tomato [J].J Experimental Botany,2011,62:3781-3798.
[40]黃智明,翁海波,席 宇,等.轉入HPT1基因的油菜種子中維生素E 含量的提高 [J].植物生理學通訊,2006,42(5):888-890.
[41]劉風珍,萬勇善,王洪剛.γ-維生素E甲基轉移酶基因轉化花生研究 [J].中國糧油學報,2005,20 (1):61-64,68.
[42]Cahoon E B,Hall S E,Ripp K G,et al.Metabolic redesign of vitamin E biosynthesis in plants for tocotrienol production and increased antioxidant content [J].Nature Biotechnology,2003,21:1082-1087.
[43]Falk J,Andersen G,Kernebeck B,et al.Constitutive over-expression of barley 4-hydroxyphenylpyruvate dioxygenase in tobacco results in elevation of vitamin E content in seeds but not in leaves [J].FEBS Lett,2003,540:35 -40.
[44]Van Eenennaam A L,Li G,Venkatramesh M,et al.Elevation of seed [alpha]-tocopherol levels using plant-based transcription factors targeted to an endogenous locus [J].Metab Eng,2004,6:101-108.
[45]Kumar R,Raclaru M,Schü Beler,et al.Characterisation of plant tocopherol cyclases and their overexpression in transgenicBrassicanapusseeds [J].FEBS Lett,2005,579:1357-1364.
[46]Blum S,Vardi M,Brown J B,et al.Vitamin E reduces cardiovascular disease in individuals with diabetes mellitus and the haptoglobin 2-2 genotype [J].Pharmacogenomics.2010; 11:675-684.
[47]Abdul Hafid S R,Radhakrishnan A K,Nesaretnam K.Tocotrienols are good adjuvants for developing cancer vaccines [J].BMC Cancer,2010,10:5.
[48]Yang Z H,Lee M J,Zhao Y,et al.Metabolism of tocotrienols in animals and synergistic inhibitory actions of tocotrienols with atorvastatin in cancer cells [J].Genes Nutr,2012,7:11-18.
10.3969/j.issn.1673-1409(S).2012.04.011
Q566
A
1673-1409(2012)04-S041-05
2012-03-20
湖北省教育廳資助項目(Q20101302)。
范麗萍(1988-),女,海南文昌人,現從事生物技術研究。
趙福永,E-mailfyzhao@yangtzeu.edu.cn。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
