ezrin表達與宮頸鱗癌組織微血管微淋巴管生成、癌細胞增殖、浸潤轉移及預后的關系
2012-11-21 02:48:30齊碧如江忠清連成瑛黃勉林岷劉兆董方嬌寧
中國實用醫藥 2012年12期
齊碧如 江忠清 連成瑛 黃勉 林岷 劉兆董 方嬌寧
埃茲蛋白(ezrin)是ezrin-radixin-moesin(ERM)家族的成員之一,1981年首先作為表皮生長因子受體蛋白酪氨酸激酶的底物被發現,之后從雞小腸微絨毛中提取鑒定[1]。ezrin作為膜細胞骨架連接蛋白,主要參與上皮細胞中細胞骨架與細胞膜之間的連接,介導信號轉導,對維持細胞形態、分化、運動性、細胞與細胞以及細胞與間質之間的的黏附有重要調節作用,并參與多種疾病的發病過程,影響腫瘤的分化、侵襲性和轉移[2]。研究表明ezrin過度表達與不同來源的多種人體腫瘤進展密切相關,可促進腫瘤侵襲轉移[3]。ezrin與宮頸鱗癌關系的研究國內少見報道。本研究采用免疫組化SP法和圖像分析技術檢測ezrin在宮頸鱗癌組織的表達情況,探討其表達與局部腫瘤微血管微淋巴管生成、癌細胞增殖、侵襲轉移及預后的關系。
1 材料與方法
1.1 材料 選自1998年3月26日至2005年6月21日在福建醫科大學附屬第一醫院手術治療的36例宮頸上皮內瘤變(cervical intraepithelial neoplasm,CIN)和68例宮頸鱗狀細胞癌(squamous carcinoma of cervix,SCC)患者作為研究對象。所有研究對象術前均未經過化療、放療及免疫治療,并均經病理確診。選取28例因功血或子宮肌瘤行子宮切除術的正常宮頸上皮(normal cervical epithelium,NCE)作為對照。CIN分級及宮頸癌組織學分級和FIGO分期參照文獻[4]。68例宮頸鱗癌患者年齡24~72歲,中位年齡45歲,<45歲者30例,≥45歲者38例。CINⅠ、Ⅱ、Ⅲ級分別為12、11和13例。鱗癌FIGO分期Ⅰ期32例,Ⅱ期36例;組織學分級Ⅰ、Ⅱ、Ⅲ級分別2、19和47例;脈管浸潤34例,盆腔淋巴結轉移17例;鏡下早期浸潤癌5例,明顯間質浸潤癌63例(淺、深肌層浸潤各15和48例)。從手術當日到2006年12月31日止,對本組68例宮頸鱗癌患者進行隨訪,共隨訪到53例,隨訪率77.94%,隨訪時間5~93個月,平均54.8個月。在隨訪到的53例中死亡14例,3年生存率74.51%(38/51),5年生存率67.44%(29/43)。
1.2 試劑 鼠抗人ezrin單克隆抗體(克隆號3 c12)、鼠抗人CD34單克隆抗體(克隆號QBEND/10)、鼠抗人MIB1單克隆抗體(克隆號 MIB1)、即用型 UltraSensitiveTMSP超敏試劑盒(Cat.No:KIT-9710)及DAB染色劑等購自福州邁新生物技術有限公司。濃縮型兔抗人VEGFR-3多克隆抗體(產品編號ZA-0267,工作濃度1∶100)購自北京中杉金橋生物技術有限公司。
1.3 方法 所有新鮮標本均經4%甲醛溶液固定,48 h內取材,石蠟包埋,連續3~5 μm厚度切片。采用免疫組化SP法檢測CIN和宮頸鱗癌組織中ezrin、CD34、VEGFR3和MIB1的表達情況。具體步驟按說明書進行。ezrin、CD34、VEGFR3和MIB1均采用檸檬酸緩沖液高壓熱力修復抗原,DAB顯色。ezrin未進行蘇木素復染,CD34、VEGFR3和MIB1進行蘇木素復染。用已知陽性片作為陽性對照,用PBS代替第一抗體作為陰性對照。
1.4 結果判斷 ezrin表達水平測定:ezrin在細胞漿和(或)細胞膜表達,陽性染色者在細胞漿和(或)細胞膜被染色呈棕黃色。用JD801圖像分析系統測定每張切片9個200倍視野ezrin陽性染色區的灰度值和光密度值,以平均光密度值的大小來表示ezrin表達水平的高低。CD34標記的微血管密度(microvessel density,MVD):CD34表達于血管內皮細胞質,呈棕黃色染色,其判斷標準及脈管浸潤的判斷標準參照文獻[5]。VEGFR3標記的微淋巴管密度(lymphatic microvessel density,LMVD):VEGFR-3陽性染色主要位于腫瘤邊緣及間質微淋巴管內皮細胞質,呈棕黃色染色,可見微淋巴管內皮細胞圍成壁薄、形態不規則的管腔結構,內無紅細胞。LMVD判斷標準參照文獻[6]。MIB1標記的增殖指數(proliferation index,PI):MIB1在細胞核表達,陽性染色者細胞核呈棕黃色。在光鏡下隨機觀察具有代表性的10個高倍視野(400),每個視野記數100個腫瘤細胞,計算其陽性標記細胞數即為PI。
1.5 統計學分析 采用SPSS 11.5統計軟件包進行分析,定量資料比較采用方差分析及t檢驗,變量間的關系采用直線相關分析,生存情況采用Kaplan-Meier曲線表示,Log rank檢驗。檢驗水準α=0.05,以P<0.05差異有統計學意義。
2 結果
2.1 ezrin在NCE、CIN及SCC組的表達 從NCE到CIN再到SCC組,ezrin表達水平顯著升高(P<0.05)。ezrin在NCE組主要表達于上皮細胞膜,在CIN組主要表達于異型細胞質,在SCC組主要彌漫性表達于癌細胞質和(或)細胞膜。ezrin在CINⅠ、Ⅱ、Ⅲ的表達水平分別為(0.0819±0.0153)、(0.0933±0.0107)、(0.1018±0.0159)。ezrin在 CINⅢ的表達水平顯著高于CINⅠ(SNK方差分析:CINⅢvs CINⅠ,P<0.05)。ezrin在 NCE、CIN及 SCC組的表達見圖 1A、1B及1C。
表1 ezrin在NCE、CIN及SCC組的表達(±s)
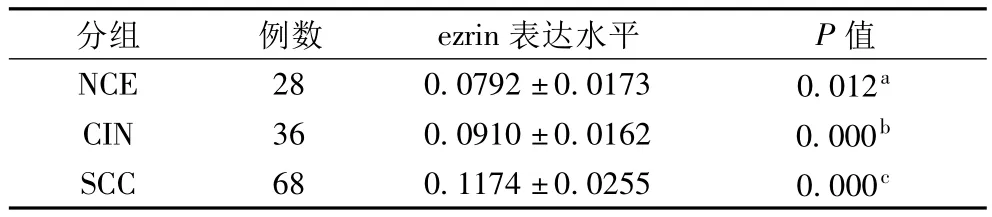
表1 ezrin在NCE、CIN及SCC組的表達(±s)
注:Dunnett`s T3方差分析:a指NCE vs CIN,P=0.012;b指CIN vs SCC,P=0.000;c指 SCC vs NCE,P=0.000
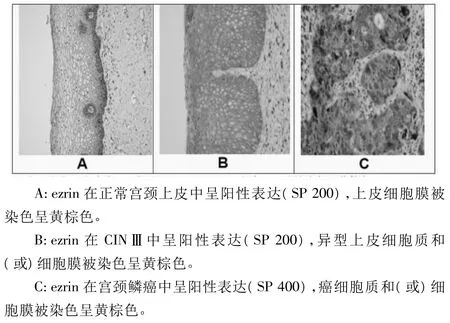
圖1 ezrin在NCE、CIN及SCC組表達
2.2 ezrin在CIN及SCC組的表達分別與MVD、LMVD及PI的關系 在NCE、CIN及SCC組,CD34標記的MVD分別為(6.3±2.9)、(18.0±4.7)及(58.2±19.6)條;VEGFR-3標記的LMVD分別為(2.24±3.8)、(6.37±5.7)和(8.3±2.8)條;Ki-67標記的 PI分別為(96.0±54.6)、(300.0±157.1)及(378.7 ±199.6)。從NCE到CIN再到SCC組,MVD、LMVD及PI均顯著升高(MVD 組:CIN vs NCE,P=0.000,SCC vs CIN,P=0.000,SCC vs NCE,P=0.000;LMVD 組:CIN vs NCE,P=0.003,SCC vs CIN,P=0.176,SCC vs NCE,P=0.000;PI組:CIN vs NCE,P=0.000,SCC vs CIN,P=0.043,SCC vs NCE,P=0.000)。將ezrin在CIN及SCC組的表達均以中位值為界分為高和低表達兩組。在CIN及SCC組,ezrin高表達組MVD、LMVD及PI分別與低表達組相比均無顯著性差異(P>0.05)。Pearson相關分析顯示,ezrin在CIN和SCC組表達均分別與MVD(CIN組:r=0.059,P=0.735;SCC組:r=0.148,P=0.233)、LMVD(CIN 組:r=0.173,P=0.314;SCC組:r=0.230,P=0.059)及 PI(CIN 組:r=0.160,P=0.359:SCC 組:r=-0.067,P=0.589)無關。CD34、VEGFR3和Ki-67在SCC組表達分別見圖2A、2B和2D。
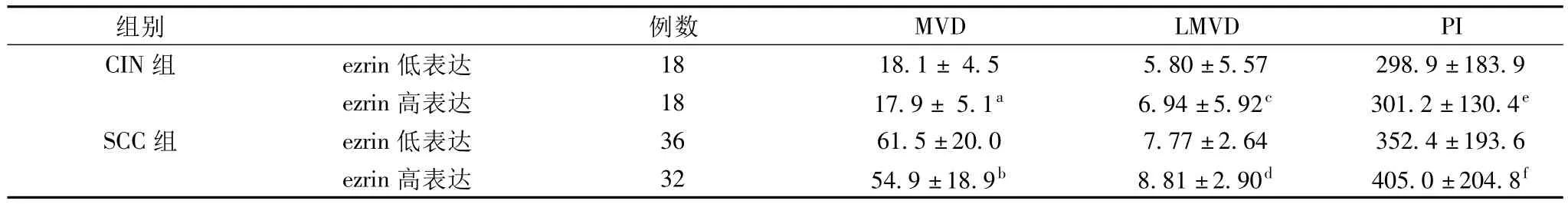
表2 ezrin在CIN和SCC組表達分別與MVD、LMVD及PI的關系
2.3 ezrin在SCC組的表達與臨床病理特征的關系 ezrin在SCC組表達與FIGO分期、浸潤深度、有否淋巴結轉移以及脈管浸潤密切相關(P<0.05)。FIGO分期為Ⅱ期、有盆腔淋巴結轉移、突破深肌層間質浸潤及有脈管浸潤者,其ezrin表達顯著高于FIGO分期為Ⅰ期、無盆腔淋巴結轉移、浸潤深度在淺肌層間質以內及無脈管浸潤者(P<0.05)(表3)。
表3 ezrin 在SCC組的表達與臨床病理特征的關系(±s)
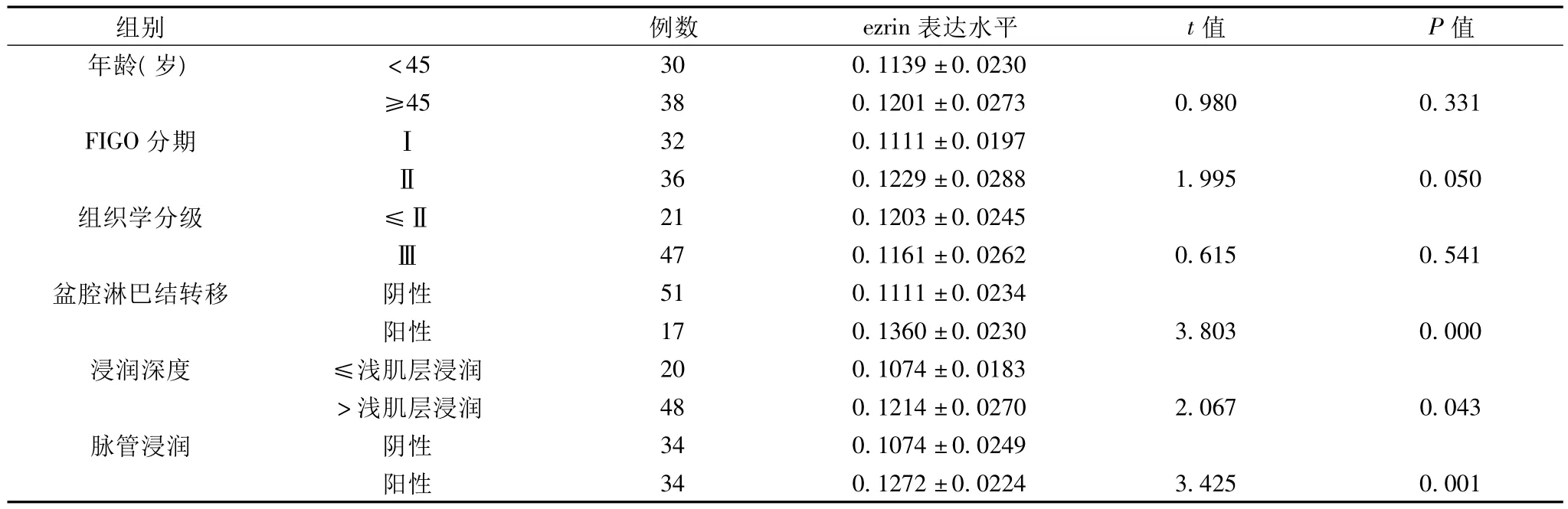
表3 ezrin 在SCC組的表達與臨床病理特征的關系(±s)
2.4 ezrin在SCC組的表達與預后的關系 將ezrin在SCC組的表達以中位數為界分為高表達組和低表達組,兩組Kaplan-Meier生存曲線見圖3。ezrin高表達組患者的生存率明顯低于低表達組,但無統計學差異(log rank檢驗,P=0.0631)。
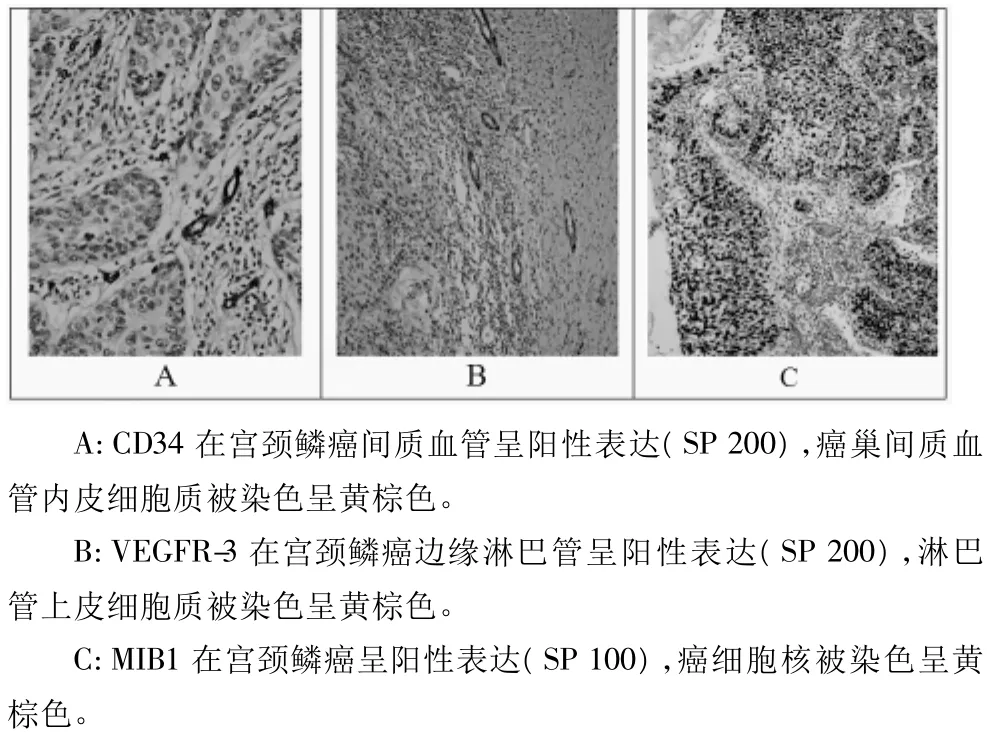
圖2 CD34、VEGFR3和MIB-1在宮頸鱗癌的表達
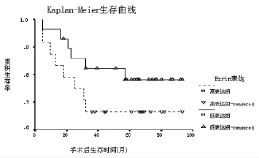
圖3 ezrin高表達組和低表達組宮頸鱗癌患者Kaplan-Meier生存曲線
3 討論
ezrin是一個分子量為82KDa的磷酸化蛋白,由585個氨基酸組成,其編碼基因為Villin2(Vil2),定位于6q25.2-6q26。ezrin蛋白N-端與細胞膜結合,C-端與F-肌動蛋白連接,參與亞細胞骨架構建和接觸識別,維持細胞膜表面的一些特殊結構如微絨毛、絲狀假足、尾足等,在細胞形態調節和細胞運動中發揮重要作用,此外還將膜蛋白如CD44、E-Cadherin等與F-肌動蛋白連接起來[2]。ezrin與膜表面受體分子、粘附分子相互作用,參與細胞信號轉導,將各種細胞膜外信號通過ezrin與膜整合蛋白直接連接而傳入細胞內[2]。ezrin參與的這些細胞活動與腫瘤的發生發展、浸潤轉移等密切相關,可能是腫瘤轉移的核心蛋白。ezrin與宮頸癌進展及預后的研究國內外少見報道。本研究結果顯示,從NCE到CIN再到SCC組,ezrin表達水平顯著增強(P<0.05)。在CIN中,ezrin在CINⅢ的表達水平顯著高于CINⅠ(P<0.05)。提示宮頸上皮細胞從受損到癌變,ezrin蛋白合成增加。ezrin與腫瘤細胞表面黏附分子CD44及其變異體形成復合物,在腫瘤細胞的浸潤轉移中有共同表達現象,并參與腫瘤細胞的轉移過程。CD44過度表達可引起膜-細胞骨架連接蛋白-ezrin的功能激活[1]。E-cadherin能反向調節CD44活性而抑制其介導的腫瘤轉移。E-cadherin丟失或調節功能降低將破壞其下游信號轉導,進而促進腫瘤轉移擴散。ezrin在E-cadherin的膜定位上起著重要作用,ezrin激活或過度表達使E-cadherin在胞內蓄積而在細胞表面表達減少,使E-cadherin被隔離在細胞內,從而導致細胞與細胞間連接破壞[7]。可見,ezrin過度表達可能導致E-cadherin功能缺失,后者對CD44的降調作用解除而使CD44表達升高,最終引起E-cadherin與CD44功能失衡,從而導致腫瘤細胞侵襲轉移能力的增強。故筆者認為,宮頸鱗癌ezrin過度表達可能在其發生發展過程中發揮重要作用。Ohtani等[8]報道,ezrin在正常子宮內膜和增殖癥子宮內膜細胞中僅表達于細胞膜,而在非典型增生子宮內膜細胞及子宮內膜癌細胞中表達于細胞漿,認為ezrin不同著色部位對子宮內膜癌病變的性質具有標識作用。本研究結果也顯示,ezrin在NCE、CIN和SCC細胞中定位不同,ezrin在NCE組幾乎表達于細胞膜上,而在CIN和SCC組異型或癌細胞質內呈彌漫性表達。上述研究提示,隨宮頸上皮惡性轉化的發生,ezrin蛋白表達上調并出現表達定位改變。可見,ezrin蛋白的表達改變可能與宮頸上皮惡性轉化密切相關,但有待于進一步研究證實。
Zhang等[9]發現,ezrin蛋白過表達與肝細胞癌增殖、侵襲和遷移正相關,下調其表達可抑制高轉移潛能肝癌細胞系侵襲力。張肖肖等[10]研究發現,ezrin蛋白在胃癌組織中高表達,并與胃癌淋巴結轉移正相關。Shen等[11]研究發現,高侵襲性人食管癌細胞株ezrin表達顯著高于對照組,且高表達ezrin的SHEEMT細胞在體內更易于侵犯軟組織、系膜和胰腺而表現更高的侵襲特性。Khanna等[12]發現高轉移能力的小兒骨肉瘤細胞株K7 m2 ezrin表達量是低轉移能力的K12的3倍,高轉移能力的K7 m2細胞轉染反義RNA抑制ezrin表達,能顯著阻止肺部轉移,而轉染ezrin-T567A(相當于失活ezrin)后,肺部轉移則完全被阻止。可見,ezrin不但足以導致腫瘤轉移,而且對于腫瘤轉移是必不可少的。本研究結果顯示,ezrin在SCC組表達與FIGO分期、浸潤深度、有否淋巴結轉移以及脈管浸潤密切相關(P≤0.05)。FIGO分期為Ⅱ期、有盆腔淋巴結轉移、突破深肌層間質浸潤及有脈管浸潤者,其ezrin表達顯著高于FIGO分期為Ⅰ期、無盆腔淋巴結轉移、浸潤深度在淺肌層間質以內及無脈管浸潤者(P≤0.05)。隨FIGO分期增加、間質浸潤加深、盆腔淋巴結轉移和脈管浸潤的發生,宮頸鱗癌組織ezrin表達水平顯著升高。張艷廷等[13]報道,ezrin和CD44V6在喉鱗癌中表達升高與組織學分級、TNM分期及淋巴結轉移關系密切,聯合檢測 ezrin和CD44V6表達有助于綜合判斷喉鱗癌的惡性度和轉移潛能。Ohtani等[14]研究發現,ezrin在高轉移能力的子宮內膜癌細胞系mEⅡL表達顯著高于低轉移能力的細胞系Ishikawa,過度表達ezrin的腫瘤細胞穿透基底膜的能力增強,而抑制ezrin表達后的兩種細胞系的侵襲能力均下降,穿透基質膜的瘤細胞數量均明顯減少,但ezrin表達對癌細胞增殖沒有影響。Chuan等[15]研究發現,前列腺癌細胞珠LNCaP-FGC采用siRNA抑制或采用過表達T567A和Y353F-ezrin突變體干擾ezrin表達活性,能顯著降低癌細胞穿透基質膜的能力,但不影響癌細胞的增殖能力。本文研究結果顯示,宮頸鱗癌ezrin表達與癌細胞增殖指數沒有顯著相關性(r=-0.067,P=0.589),與 Ohtani等[14]和 Chuan 等[15]的結果一致。ezrin 與腫瘤新生血管生成關系的研究國內外少見報道。有學者報道,Ezrin能抑制內皮細胞擴增和血管生成[16]。筆者發現宮頸鱗癌組織ezrin表達與微血管密度無顯著相關性(r=0.148,P=0.233)。ezrin與腫瘤新生淋巴管生成關系的研究國內外未見報道。筆者結果顯示,宮頸鱗癌組織ezrin表達與微淋巴管密度無顯著相關性(r=0.230,P=0.059)。故筆者認為,ezrin在CIN中表達增強提示癌細胞易于突破基底膜向間質浸潤,ezrin在SCC中過度表達顯示癌細胞向深部浸潤的同時進而突破脈管基底膜浸潤淋巴管和血管,最終發生遠處轉移。在宮頸鱗癌發生發展過程中,ezrin可能不直接參與癌細胞增殖和新生血管淋巴管形成的調節,而直接參與增強癌細胞穿透基底膜,致使腫瘤侵襲轉移。
文獻報道,多種人體惡性腫瘤ezrin表達上調與患者預后密切相關,ezrin過度表達生存期短、預后差[17-20]。但也有研究認為,惡性腫瘤ezrin表達下調者預后差。Moilanen等[21]報道,漿液性卵巢癌ezrin蛋白表達下調者預后差。絨毛膜癌ezrin蛋白表達降低后侵襲力反而增強[22]。筆者資料可以看出,ezrin高表達組患者的生存率明顯低于低表達組,但未達統計學差異(log rank檢驗,P=0.0631)。故筆者認為,ezrin表達可能具有組織和器官的特異性,其表達水平并非完全與腫瘤轉移及預后平行,其在腫瘤轉移中的復雜作用有待進一步研究探討。
[1]Martin TA,Harrison G,Mansel RE,et al.The role of the CD44/ezrin complex in Cancer metastasis. Crit Rev Oncol Hematol,2003,46(2):165-186.
[2]Haedicke J,de Los Santos K,Goff SP,et al.The ezrin-radixinmoesin family member ezrin regulates stable microtubule formation and retroviral infection.J Virol,2008,82(9):4665-4670.
[3]Sizemore S,Cicek M,Sizemore N,et al.Podocalyxin increases the aggressive phenotype of breast and prostate cancer cells in vitro through its interaction with ezrin.Cancer Res,2007,67(13):6183-6191.
[4]樂杰.婦產科學.第7版.北京:人民衛生出版社,2008:261-268.
[5]林岷,江忠清,朱鳳川,等.宮頸癌CD44V6表達與局部腫瘤血管生成、癌細胞增殖及浸潤轉移的關系.福建醫科大學學報,2005,39(1):41-44.
[6]林岷,江忠清,連成瑛,等.宮頸鱗狀細胞癌局部淋巴管生成與浸潤轉移及預后的關系.福建醫科大學學報,2008,42(2):116-120.
[7]Xu Y,Yu Q.E-cadherin negatively regulates CD44-hyaluronan interaction and CD44-mediated tumor invasion and branching morphogenesis.J Biol Chem,2003,278(10):8661-8668.
[8]Ohtani K,Sakamoto H,Rutherford T,et al.Ezrin,a membranecytoskelet al linking protein,is highly expressed in atypical endometrial hyperplasia and uterine endometrioid adeno-carcinoma.Cancer Lett,2002,179(1):79-86.
[9]Zhang Y,Hu MY,Wu WZ,et al.The membrane-cytoskeleton organizer ezrin is necessary for hepatocellular carcinoma cell growth and invasiveness.J Cancer Res Clin Oncol,2006,132(11):685-697.
[10]張肖肖,武洋,楊君,等.ezrin和c-Met基因編碼蛋白表達與胃癌發生和轉移的關系和意義.中國腫瘤臨床,2009,36(7):404-408.
[11]Shen ZY,Xu LY,Chen MH,et al.Upregulated expression of ezrin and invasive phenotype in malignantly transformed esophageal epithelial cells.World J Gastroenterol,2003,9(6):1182-1186.
[12]Khanna C,Wan X,Bose S,et al.The membrane-cytoskeleton linker ezrin is necessary for osteosarcoma metastasis.Nat Med,2004,10(2):182-186.
[13]張艷廷,皇甫輝,田俊.ezrin蛋白和CD44V6在喉鱗癌組織中的表達及臨床意義.山西中醫學院學報,2009,10(1):50-52.
[14]Ohtani K,Sakamoto H,Rutherford T,et al.Ezrin,a membranecytoskelet al linking protein,is involved in the process of invasion of endometrial cancer cells.Cancer Lett,1999,147(1-2):31-38.
[15]Chuan YC,Pang ST,Cedazo-Minguez A,et al.Androgen induction of prostate cancer cell invasion is mediated by ezrin.J Biol Chem,2006,281(40):29938-29948.
[16]Kishore R,Qin G,Luedemann C,et al.The cytoskelet al protein ezrin regulates EC proliferation and angiogenesis via TNF-alpha-induced transcriptional repression of cyclin A.J Clin Invest,2005,115(7):1785-1796.
[17]Carneiro A,Bendahl PO,Akerman M,et al.Ezrin expression predicts local recurrence and development of metastases in soft tissue sarcomas.J Clin Pathol,2011,64(8):689-694.
[18]Elzagheid A,Korkeila E,Bendardaf R,et al.Intense cytoplasmic ezrin immunoreactivity predicts poor survival in colorectal cancer.Hum Pathol,2008,39(12):1737-1743.
[19]Kobel M,Lanshammer T,Huttelmaier S,et al.Ezrin expression is related to poor prognosis in FIGO stage I endometrioid carcinomas.Mod Pathol,2006,19(4):581-587.
[20]Deng X,Tannehill-Gregg SH,Nadella MV,et al.Parathyroid hormone-related protein and ezrin are up-regulated in human lung cancer bone metastases.Clin Exp Metastasis,2007,24(2):107-119.
[21]Moilanen J,Lassus H,Leminen A,et al.Ezrin immunoreactivity in relation to survival in serous ovarian carcinoma patients.Gynecol Oncol,2003,90(2):273-281.
[22]Karmakar S,Das C.Modulation of ezrin and E-cadherin expression by IL-1 beta and TGF-beta 1 in human trophoblasts.J Reprod Immunol,2004,64(1-2):9-29.
