外源MeJA誘導煙草葉面防御反應
2013-01-17 05:36:06武東玲魏躍偉
中國煙草科學 2013年5期
關鍵詞:煙草
馮 琦,王 永,武東玲,魏躍偉,崔 紅*
(1.河南農業大學煙草學院,鄭州 450002;2.三門峽市煙草公司澠池分公司,河南 澠池 472044)
腺毛是植物表面的特化器官。作為植物“感受器”,它能夠感受外界環境細微的壓力并誘導一系列防御反應[1]。人們發現茉莉酸及其衍生物(JAs)對腺毛防御具有重要作用。外源茉莉酸可替代傷害刺激,有效地促進植物新生葉片腺毛密度大幅度增加[2-4]。煙草莖葉腺毛密布,種類繁多,分泌旺盛。在水分脅迫條件下,腺毛細胞中葉綠體類囊體膜降解,高電子致密物質逐漸增多[5],煙葉表面西柏三烯二醇的含量升高[6];在遮蔭條件下,腺毛葉綠體類囊體片層發達,葉綠素熒光強烈,黑色嗜鋨顆粒稀少,葉面西柏烷類化合物的含量大大降低[7]。逆境條件下腺毛形態學和物質代謝所發生的變化,說明煙草腺毛在煙株防御過程中具有重要作用,對其反應機制的深入解析是實現高效調控的重要前提。
本試驗采用外源茉莉酸甲酯(MeJA)處理煙苗,并對葉片腺毛密度結構、葉面化學組分及蚜蟲趨避性進行分析檢測,研究MeJA對煙草腺毛形態發生、物質代謝和煙株抗性的影響,并探討煙草腺毛防御系統的反應機制,為煙草腺毛防御代謝工程奠定理論基礎。
1 材料與方法
1.1 材料
以煙草品種中煙100為材料,依照《優質煙栽培技術規范》進行漂浮育苗,成苗后移栽到花盆。1個月后選擇生長態勢一致的煙株,用 0.8 mM MeJA(無水乙醇含量 0.8%)充分噴施,對照組用0.8%的無水乙醇溶液處理,對照和處理組隔離3 d后,隨機擺放在實驗網棚中培養。試驗于2011年6月1日移栽,8月30日結束,試驗期間平均氣溫26.1 ℃。
1.2 試驗方法
1.2.1 腺毛密度統計及形態觀察 自煙株處理后,待頂部第1片新葉長到10 cm時,小心摘取,直接撕取葉片下表皮在MOTIC光學顯微鏡下統計腺毛密度[8]。新鮮葉片沿主脈兩側切下約5 mm×8 mm的平整小塊葉片,直接使用VEGA Ⅱ LMU掃描電子顯微鏡(Tescan公司,捷克)觀察葉片下表面腺毛形態及類型,并拍照。掃描電鏡采用氣體二次電子成像,工作電壓 20 kV,工作距離 21 mm(21.25~21.70 mm),樣品室和鏡筒狀態選用高真空(10-6Pa)。
1.2.2 葉面化學成分分析 采用有機溶劑萃取的方法[9],對MeJA處理后0、3、7、11 d的葉片進行葉面化學成分提取,并采用紫外分光光度計法[10]進行葉面化學成分總量檢測。
對處理后第 7天的葉面化學成分進行 GC/MS檢測:于二氯甲烷提取液中加入內標(2.020 mg/mL的蔗糖八乙酸酯和 2.542 mg/mL的正十七烷醇的500 mL二氯甲烷溶液)1 mL,旋轉蒸發濃縮,硅烷化處理后,采用 GC/MS與微機聯用進行定性、定量分析。色譜儀為HP-5890,質譜儀是vc-70SE。GC條件:色譜柱:DB-5MSUI石英毛細管柱(30 m×0.25 mm i.d.×0.25 μm d.f.);進樣口溫度:250 ℃;程序升溫:40 ℃保持2 min,然后以6 ℃/min升溫至180 ℃保持2 min,以2 ℃/min升溫至280 ℃保持20 min;載氣:高純氦氣;載氣流速:恒流0.8 mL/min;進樣量:1.0 μL,分流比:15:1。MS條件:傳輸線溫度:250 ℃;EI離子源溫度:280 ℃;電離能量:70 eV;質量數范圍50~650 amu;檢索譜庫:NIST08。
1.2.3 葉面腺毛基因表達分析 葉片腺毛的獲得:參照文獻[11]的方法進行改良。選取幼嫩健康的葉片液氮中低溫固定形態5 min,使用硬度適中的毛刷由葉基向葉尖方向將腺毛刷至盛有適量液氮的研缽中,每個樣品刷取約100 cm2的葉片,刷取完成后直接研磨提取。
腺毛 RNA提取及反轉錄:RNA提取使用QIAGEN公司的RNeasy Plant Mini Kit試劑盒,反轉錄使用 NEWBIO INDUSTRY公司生產的M-MLV反轉錄酶,反轉錄的cDNA用于PCR檢測。
引物設計:采用Premier Prime 5.0軟件,根據Genbank發布的L25、DXR、CBT和CYP71D16序列設計各基因擴增引物(表1),用于PCR檢測。其中L25是煙草核糖體蛋白[12],本研究中作為內參基因。
PCR檢測:PCR反應體系為20 μL。PCR擴增程序為:94 ℃ 5 min 預變性;94 ℃ 30 sec變性,48~51 ℃ 30 sec退火(具體按各引物Tm值設定),72 ℃1 min延伸,共30個循環;72 ℃延伸10 min。
結果檢測:PCR產物使用 1.5%瓊脂糖凝膠進行檢測,并使用 Manual BioSpectrum Imaging System(UVP公司,美國)進行觀察拍照。
1.2.4 蚜蟲選擇性試驗 分別選擇處理與對照兩片幼葉,分別置于15 cm直徑的培養皿中;挑選健康的無翅蚜于一小玻璃杯中,將玻璃杯等距離倒扣在葉片之間,輕敲杯底使蚜蟲掉落于培養皿上,移去玻璃杯,使蚜蟲自由活動;于 24 ℃恒溫光照培養箱中培養7 h后,統計處理與對照葉片上的蚜蟲數。結果為3次重復的平均值。
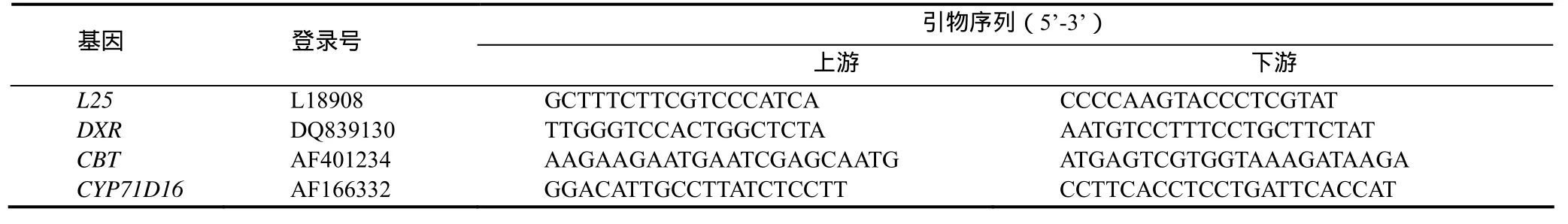
表1 擴增引物序列Table 1 Primer sequence
2 結 果
2.1 MeJA處理對腺毛形態發生的影響
MeJA噴施煙株后,對新葉進行掃描電鏡觀察,結果如圖1所示。可以看出,MeJA處理煙株的新葉,腺毛密度高,且長柄多腺頭腺毛較多,體型較大(圖1-A),而對照煙株葉片腺毛密度相對較低,體型較小,長柄單細胞腺頭腺毛較多(圖1-B)。體視鏡下對葉片下表皮的腺毛腺密度進行統計,結果如圖2所示。可以看出,MeJA處理與對照相比,煙株新葉腺毛密度有所提高;但短柄腺毛以及無頭腺毛密度差異不顯著,長柄腺毛尤其是長柄多腺頭細胞差異達到顯著水平。
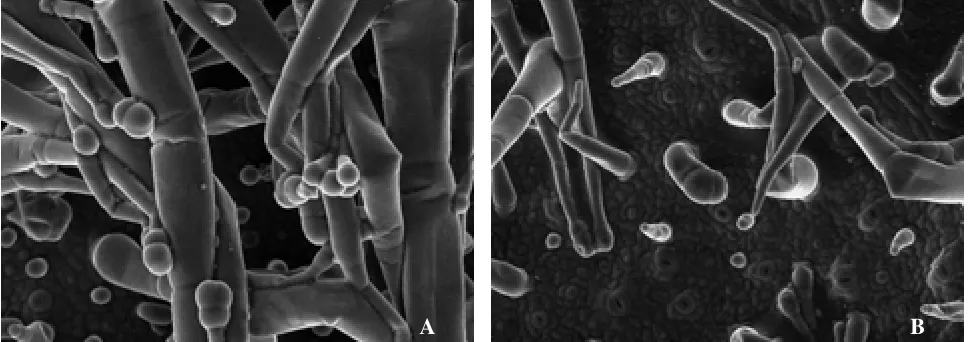
圖1 MeJA處理對葉面腺毛形態的影響Fig.1 Effect of treatment on leaf trichomes shape

圖2 MeJA處理對煙株第1片新葉腺毛密度的影響Fig.2 Effect of MeJA spraying on trichomes density of the first new leaf
2.2 葉面化學成分分析
利用分光光度法,對MeJA處理后不同時間的葉片進行葉面成分分析,葉面化學成分總量變化趨勢如圖3所示。可以看出,在第3天,MeJA處理的葉片和對照葉片在葉面化學成分含量上就有明顯不同。雖然,隨著時間的延長,對照煙株葉面分泌物也呈現緩慢上升的趨勢,但MeJA處理能使煙株葉面分泌物含量大幅度提高,并在處理后第7天達到積累高峰,隨后有所下降,但仍顯著高于對照。
采用GC/MS技術,對處理后第7天的煙株葉面分泌物成分進行檢測,峰值圖如圖4所示,結果見表2。從中可以發現,無論是在處理還是對照中,西柏三烯二醇都是葉面化學成分中含量最高的組分;在MeJA處理后,該組分含量顯著增加,說明MeJA刺激誘導了該物質的生物合成和積累。烷烴類物質種類繁多,難以準確定性;與對照相比,MeJA處理葉片中烷烴類物質含量大多表現出小幅度增加的趨勢。實驗中檢測到5種蔗糖酯類物質,與對照相比,在MeJA處理葉片中各有升降,但變幅不大,說明該類物質受MeJA的影響并不顯著。

圖3 葉面分泌物含量變化趨勢Fig.3 Changes of leaf secretion

圖4 MeJA噴施第7天葉面分泌物化學峰值比較Fig.4 Comparison of the leaf secretion on the 7th day
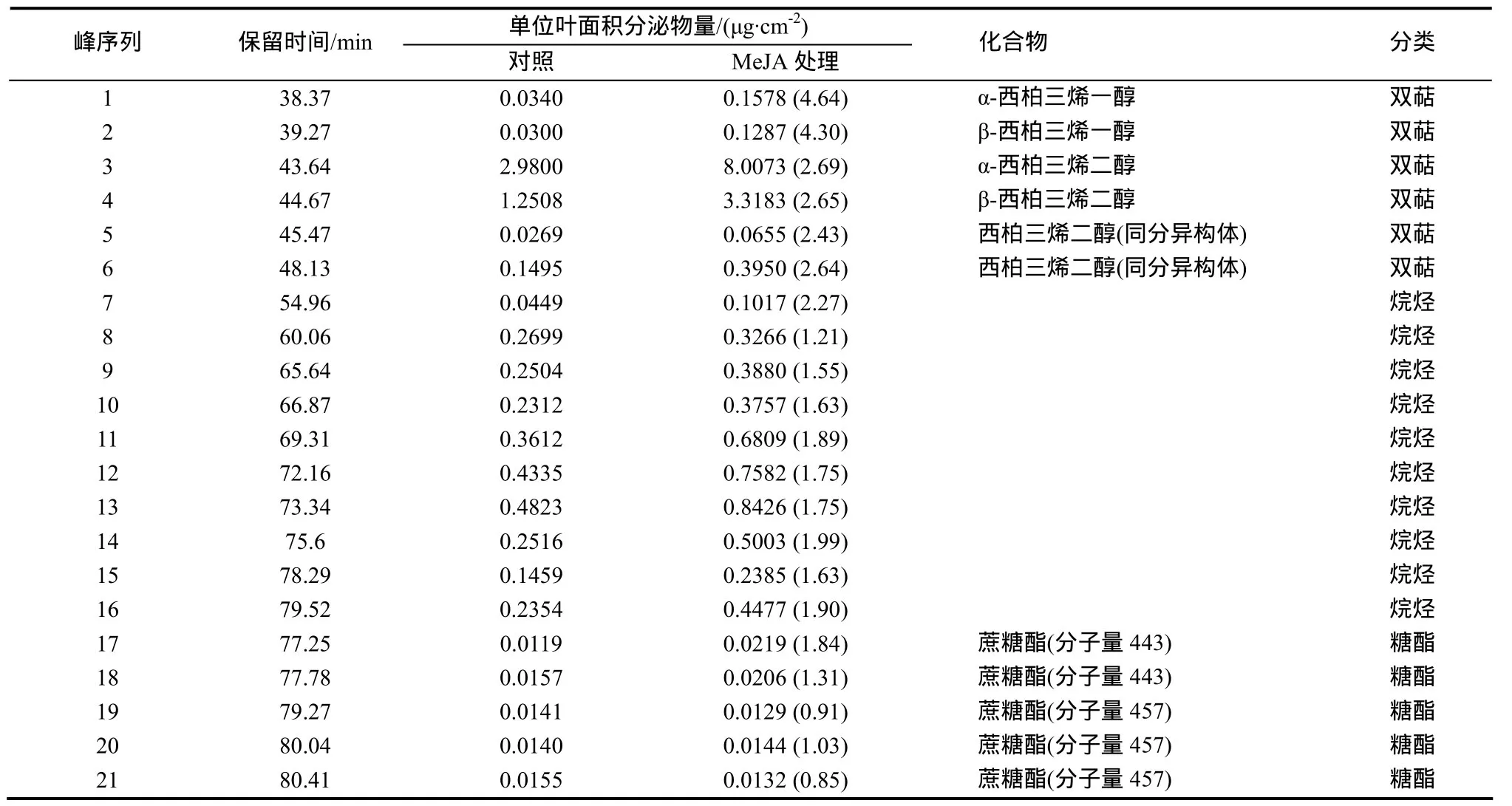
表2 MeJA噴施第7天葉面分泌物化學成分分析Table 2 Chemical constituents of the leaf secretion on the 7th day
2.3 基因表達分析
在MeJA處理后,分別在處理0 d和處理后1、2、3、4、5、6、7 d刷取葉面腺毛,提取腺毛總 RNA,利用RT-PCR技術檢測萜類代謝相關基因轉錄水平的表達變化,如圖5所示。脫氧磷酸木酮糖還原異構酶(DXR)催化脫氧磷酸木酮糖(DXP)形成甲基赤鮮糖醇-4-磷酸(MEP),是類萜代謝前體物質異戊二烯(IPP)合成的關鍵酶。西柏三烯一醇合成酶(CBT)是二萜環化酶,催化牻牛兒基牻牛兒基焦磷酸(GGPP)向西柏三烯一醇的轉化,西柏三烯一醇羥化酶(CYP71D16)則催化西柏三烯一醇向西柏三烯二醇的轉化。這三個酶的基因與葉面腺毛主要分泌物的合成密切相關,但RT-PCR結果表明,三基因的轉錄均未隨MeJA的處理而發生明顯的變化。
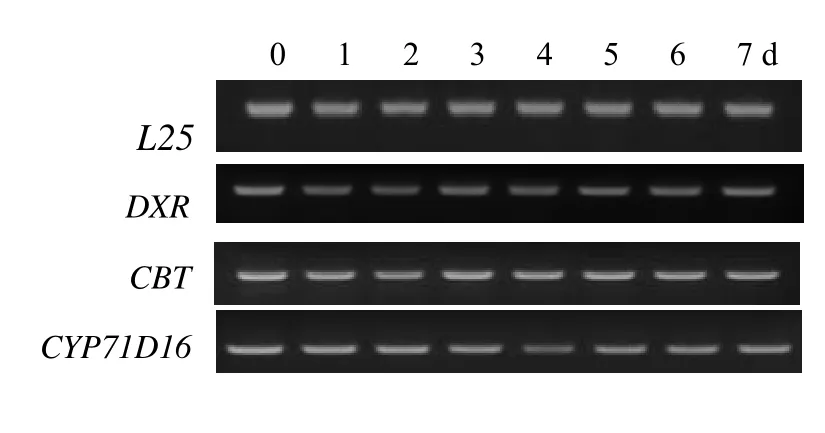
圖5 腺毛二萜代謝相關基因表達分析Fig.5 Gene expression analysis by RT-PCR
2.4 MeJA處理對煙株蚜蟲抗性的影響
MeJA噴施處理7 d后,對煙株幼葉進行蚜蟲選擇性實驗。在放有處理和對照葉片的密閉容器中(圖6-A),無翅蚜通過爬行尋找喜好的棲身之地。7 h之后,統計葉片上的蚜蟲數目,發現更多的蚜蟲集聚在未噴施 MeJA的葉片上(圖 6-B)。表明MeJA處理使得葉片對蚜蟲的驅避性增強。

圖6 葉片對蚜蟲的抗性比較Fig.6 Comparison of the leaf resistance to aphid
3 討 論
煙株莖、葉表面覆蓋著濃密的腺毛。它不僅形成了煙株與環境間的物理屏障,腺毛主要分泌物-西柏烷類二萜化合物也構筑了煙株抵御病原微生物侵襲的化學防線。由煙草腺毛構筑的煙草葉面防御體系,是煙株防御體系的重要組成部分,但目前人們對其反應模式及作用機制尚缺乏認識。
本實驗采用MeJA噴灑煙株后,葉片對蚜蟲的驅避效果證明煙草葉面防御能力得到了增強。處理后煙株新生葉片腺毛密度發生了明顯變化,但只有長柄多腺頭腺毛的差異達到了顯著水平。長柄多細胞腺頭腺毛是煙草腺毛中分泌最旺盛的類型,說明MeJA對煙草分泌型腺毛誘導的選擇性。這與同樣為多腺毛類型植物的番茄研究結果相同[2]。但在Traw等[4]的報道中,擬南芥腺毛雖為單一的非分泌型腺毛,它的發生也受到MeJA的強烈誘導,表明了不同植物中腺毛發生誘導機制的不同。在已發育葉片中,MeJA處理對腺毛密度和類型產生影響不大,但卻使煙株葉面化學成分發生了明顯變化,尤其是腺毛分泌物——西柏烷類化合物含量明顯提高。在有關曼陀羅腺毛的研究[13]中報道,MeJA處理并不改變腺毛的密度及類型,卻可以使腺毛主要分泌物acylsugars的含量提高44%。在番茄研究[14]中發現,MeJA處理可誘導腺毛單萜環化酶LeMTS1的活躍表達。但本實驗中,對煙草葉面腺毛基因表達分析表明,與煙草腺毛分泌物——西柏烷類化合物合成相關的基因在轉錄水平并未受到MeJA的誘導。關于植物類萜代謝中DXS,DXR和HDR酶受到轉錄后調節已被研究所證實[15],因此推測煙草腺毛中二萜化合物的生物合成存在著相同的調節機制。
4 結 論
本研究發現:(1)噴施MeJA能誘導煙草葉面防御反應,使葉片對蚜蟲的驅避性增強;(2)煙草葉面防御反應的主要表現是誘導了分泌型腺毛的發生,是導致西柏烷類化合物為主的葉面分泌物增加的主要原因;(3)西柏烷類化合物合成相關的基因在轉錄水平上并不受MeJA的誘導,推測煙草二萜代謝具有較復雜的轉錄后調節機制。
[1]Tooker J F, Peiffer M, Luthe D S, Felton G W.Trichome as sensors[J].Plant signaling & behavior, 2010(5):73-75.
[2]Boughton A J, Hoover K, Felton G W.Methyl jasmonate application induces increased densities of glandulartrichomes on tomato, Lycopersicon esculentum[J].Chem.Ecol, 2005, 31: 2211-2216.
[3]Kobayashi H, Yanaka M, Ikeda T M.Exogenous Methyl Jasmonate Alters Trichome Density on Leaf Surfaces of Rhodes Grass (Chloris gayana Kunth)[J].Plant Growth Regul, 2010 29: 506-511.
[4]Traw B M, Bergelson J.Interactive effects of jasmonic acid, salicylic acid and gibberellin on induction of trichomes in Arabidopsis[J].Plant Physiol, 2003, 133:1367-1375.
[5]張華,崔紅.水分脅迫對煙草腺毛超微結構的影響[J].中國煙草學報,2008,14(5):45-47.
[6]Johnson A W.Tobacco leaf trichomes and their exudates[J].Tobacco Science, 1985, 29: 67-72.
[7]翁夢苓,崔紅.遮陰對煙草葉片腺毛形態結構及分泌物含量的影響[J].河南農業大學學報,2008,42(6):613-616.
[8]時向東,劉國順,韓錦峰,等.不同類型肥料對烤煙葉片腺毛密度、種類及分布規律的影響[J].中國煙草學報,1999,5(2):19-22.
[9]Nielson M T, Severson R F.Variation of flavor components on leaf surfaces of tobacco genotypes differing in trichome density[J].Agric Food Chem, 1990,38: 467-471.
[10]張小全,楊志曉,李欽奎,等.分光光度法定量測定煙草葉面分泌物含量研究[J].華南農業大學學報,2011,32(4):18-20.
[11]Erming W, Joseph D P.Suppression of P450 hydtoxylase gene in plant trichome glands enhances naturalproduct-based aphid resistance[J].Nature Biotechnology,2001, 19: 371-374.
[12]Schmidt G W, Delaney S K.Stable internal reference genes for normalization of real-time RT-PCR in tobacco(Nicotiana tabacum) during development and abiotic stress[J].Mol Genet Genomics, 2010, 283: 233-241.
[13]Hare J D, Walling L L.Constitutive and jasmonateinducible traits of Datura wrighti[J].Journal of Chemical Ecology, 2006, 32: 29-47.
[14]Van Schie C C, Haring M A, Schuurink R C.Tomato linalool synthase is induced in trichomes by jasmonic acid[J].Plant Molecular Biology, 2007, 64: 251-263.
[15]Cordoba, E M Salmi, P Leon.Unravelling the regulatory mechanisms that modulate the MEP pathway in higher plants[J].Exp Bot, 2009, 60(10): 2933-2943.
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
