與水稻品種特異性、一致性、穩(wěn)定性(DUS)測試性狀相關(guān)聯(lián)的分子標(biāo)記分析
2013-02-23 04:57:20王艷平李華勇張繼紅
江蘇農(nóng)業(yè)學(xué)報 2013年2期
關(guān)鍵詞:水稻
王艷平, 李華勇, 沈 奇, 張繼紅, 王 平, 吳 燕
(江蘇省農(nóng)業(yè)科學(xué)院糧食作物研究所,江蘇 南京210014)
植物新品種權(quán)作為一種知識產(chǎn)權(quán)的保護(hù)在中國乃至世界的發(fā)展極為迅速,因此,建立完善、先進(jìn)、可行的植物DUS(植物新品種特異性、一致性和穩(wěn)定性)測試體系十分必要。水稻是中國最主要的糧食作物之一。水稻生產(chǎn)在國民經(jīng)濟(jì)中占有重要地位,使得中國的水稻育種發(fā)展極快,每年申請品種權(quán)的數(shù)量迅猛增加。如何保護(hù)新品種的知識產(chǎn)權(quán),對于中國水稻育種的健康、持續(xù)發(fā)展具有重要的現(xiàn)實意義。DUS 測試是新品種授權(quán)、判斷品種真實性的重要依據(jù),也是植物品種權(quán)侵權(quán)等案件判定的重要技術(shù)手段[1]。在品種授權(quán)、市場準(zhǔn)入和品種鑒定等方面起著不可替代的作用,是現(xiàn)代種植業(yè)健康發(fā)展的重要技術(shù)保障。申請品種的特異性(Distinctness)、一致性(Uniformity)和穩(wěn)定性(Stability)(簡稱DUS)是國際植物新品種保護(hù)聯(lián)盟(UPOV)規(guī)定的授權(quán)條件之一。因此要做到品種權(quán)審查和授權(quán)的客觀、公正和準(zhǔn)確,對申請品種的DUS 測試就顯得十分重要。
DUS 測試是一個復(fù)雜的技術(shù)檢測過程,它不僅要判定品種的特異性,而且還要判定該品種是否具有穩(wěn)定性、一致性。目前,中國及國際上通行的新品種DUS 測試標(biāo)準(zhǔn)主要是建立在部分生物學(xué)性狀(包括農(nóng)藝性狀、品質(zhì)性狀和抗逆性狀等)的田間測定基礎(chǔ)上的,在DUS 測試上包括了50 個以上的性狀和系列標(biāo)準(zhǔn)比較品種。如水稻DUS 測試必需檢測的性狀共有52 個。這些性狀分成3 種性狀類型:質(zhì)量性狀、假質(zhì)量性狀和數(shù)量性狀。在對測試品種與標(biāo)準(zhǔn)品種、近似品種的外觀和農(nóng)藝性狀、經(jīng)濟(jì)性狀的田間測定和比較分析時,鑒定結(jié)果容易受環(huán)境因素的影響,如在不同測試地點(diǎn)、不同測試時期某些性狀(如有關(guān)顏色的性狀)會表現(xiàn)不穩(wěn)定;在測試過程中也經(jīng)常出現(xiàn)因不同測試人員對同一作物同一性狀的認(rèn)識存在偏差,導(dǎo)致某些性狀代碼判定結(jié)果不一致的問題。同時根據(jù)田間測定分析判定品種的特異性、一致性和穩(wěn)定性工作量十分巨大,也會導(dǎo)致測試工作的誤差加大。因此,探索更加穩(wěn)定、可靠、簡便,并有較高區(qū)分能力的鑒定技術(shù)對水稻新品種權(quán)保護(hù)和遺傳資源登記工作尤其重要。
DNA 分子標(biāo)記的快速發(fā)展,為品種的準(zhǔn)確鑒定及DUS 測試提供了新的有力手段。隨著水稻基因圖譜、基因組測試的完成,越來越多的控制生物學(xué)性狀的基因被定位,并得到了與其連鎖的分子標(biāo)記,如與紅色種皮基因Rc 連鎖的標(biāo)記RID14[2]、紫色種皮基因Pb 連鎖的標(biāo)記CAPSRa[3]、與水稻半矮稈基因Sd-1 連鎖的標(biāo)記RG220、RG109[4]、與長穂頸基因eui 連鎖的標(biāo)記RM3673、RM0012[5]、與披葉基因dl(t)連鎖的標(biāo)記RM1324[6]、與粒長基因GL3 連鎖標(biāo)記PSM379、RM16[7]等等。諸如此類的性狀在水稻DUS 測試指南中占很大比重。使用與這些性狀基因緊密連鎖的DNA 標(biāo)記,有助于在DUS 測試中快速篩選近似品種和測試性狀代碼的判定,從而提高水稻DUS 測試的準(zhǔn)確性和測試的效率。
關(guān)聯(lián)分析方法在水稻、大豆、小麥等作物的農(nóng)藝性狀研究中已得到較多應(yīng)用[8-12]。Yan 等[13]利用美國農(nóng)業(yè)部水稻核心種質(zhì)的90 份材料與109 個DNA 標(biāo)記對柱頭和穗部性狀進(jìn)行了關(guān)聯(lián)分析。Huang 等[14]通過測序517 個水稻地方品種,分析了近360 萬個SNP(single nucleotide polymorphisms)標(biāo)記與14 個農(nóng)藝性狀之間的關(guān)聯(lián)。近年來,作物新品種保護(hù)日益受到重視,但到目前為止對水稻新品種保護(hù)的DUS 測試中很多性狀尚未進(jìn)行標(biāo)記的關(guān)聯(lián)分析研究。本試驗用45 份材料構(gòu)成的品種群體對38 個DUS 測試性狀與45 個分子標(biāo)記進(jìn)行關(guān)聯(lián)分析,旨在發(fā)掘與水稻DUS 測試性狀相關(guān)聯(lián)的分子標(biāo)記,為水稻新品種測試提供輔助技術(shù)手段。
1 材料與方法
1.1 材料
供試材料45 份,由農(nóng)業(yè)部植物新品種保護(hù)辦公室提供,為用于水稻DUS 測試的標(biāo)準(zhǔn)品種,品種名稱及編號見表1。
1.2 方法
1.2.1 農(nóng)藝性狀鑒定 45 個水稻品種2011 年正季種植在江蘇省農(nóng)業(yè)科學(xué)院溧水植物科學(xué)基地。5月6 日播種,6 月7 日單株移栽。每個品種均種植4行,每行12 株,株行距20 cm×20 cm,2 次重復(fù)。常規(guī)田間管理。按照《植物新品種特異性、一致性和穩(wěn)定性測試指南—水稻》(GB/T 19557.7,2010 年報批稿)調(diào)查45 個品種的38 個性狀。目測性狀(VG/VS)按照測試指南判定性狀代碼,代碼最小值為1,最大值為9,性狀“穗:芒顏色(初期)”最大值為10;測量性狀(MS)調(diào)查20 個個體,數(shù)據(jù)處理在Excel 2003 程序中進(jìn)行。
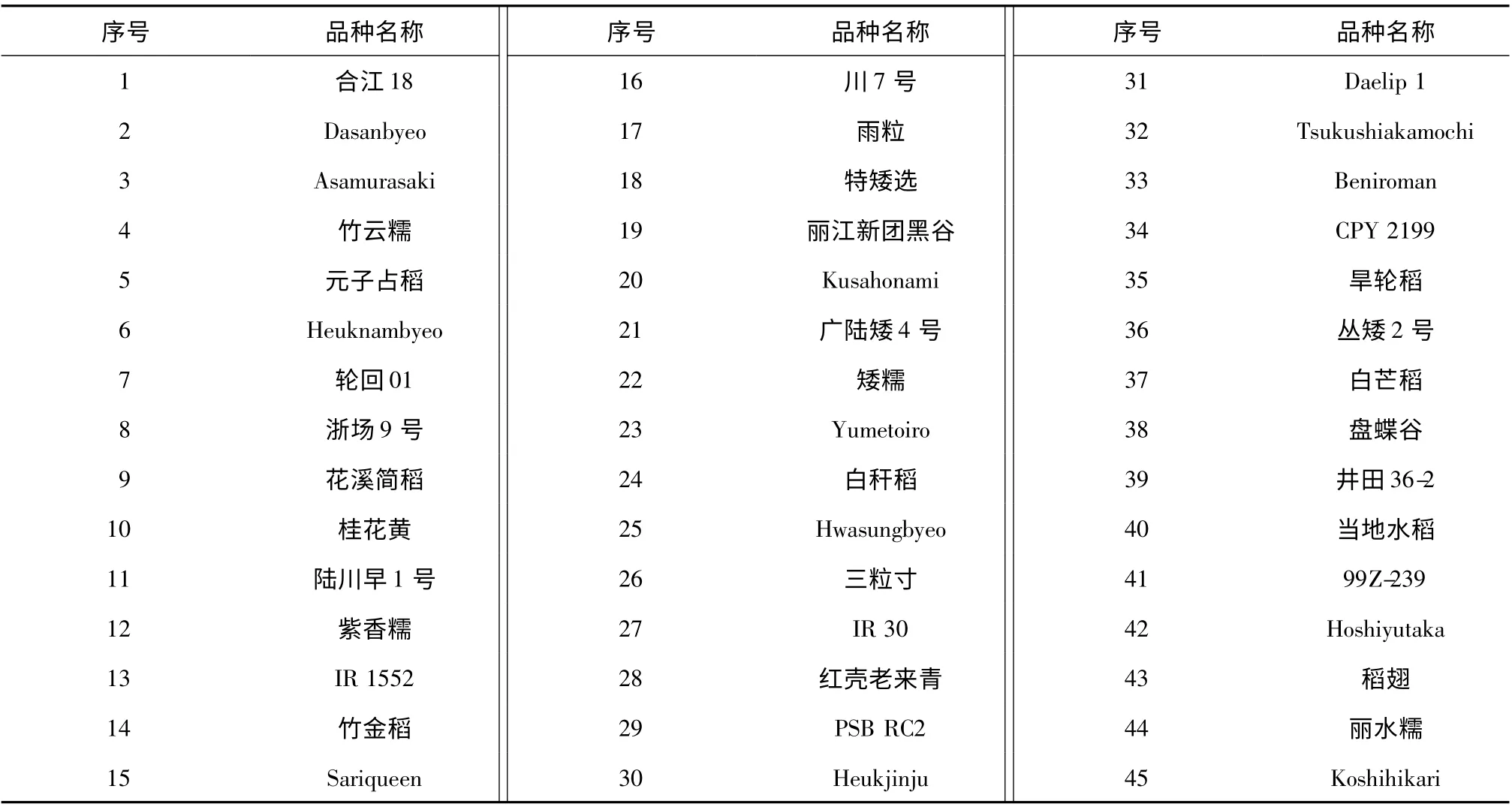
表1 供試材料Table 1 List of cultivars used in this study
1.2.2 基因組DNA 的提取 基因組DNA 提取采用改進(jìn)的SDS 方法:取水稻幼苗葉片0.5 g,剪碎,放入2 ml 離心管中,往離心管中加入液氮,放入研磨儀,磨成粉狀后立即加入65 ℃預(yù)熱的DNA 提取液800 μl 左右,劇烈搖動混勻,并在65 ℃水浴中保溫30 ~50 min,保溫期間搖動幾次。冷卻至室溫后加入等體積三氯甲烷/異戊醇(24∶ 1),顛倒混勻,室溫下靜置5 ~10 min,使水相和有機(jī)相分層。12 000 g 離心10 min。移取上清液至另一2 ml 離心管中,加入約1.5 倍體積的預(yù)冷乙醇,輕緩顛倒混勻,經(jīng)12 000 g 離心3 min 至分相,棄上清液。用70%乙醇溶液洗滌2 遍,自然條件下干燥后,加入200 μl 1 ×TE 緩沖液溶解沉淀。用瓊脂糖凝膠電泳檢測DNA 濃度和質(zhì)量,將DNA 稀釋到20 ng/μl。置于-20 ℃中保存?zhèn)溆谩?/p>
1.2.3 引物 查閱文獻(xiàn),選擇與部分農(nóng)藝性狀連鎖的21 個分子標(biāo)記和用于水稻品種DNA 鑒定的24個SSR 標(biāo)記(表2),用于45 個水稻品種的標(biāo)記基因型鑒定。引物由上海英駿生物技術(shù)有限公司合成。
1.2.4 PCR 擴(kuò)增及擴(kuò)增產(chǎn)物的檢測 PCR 擴(kuò)增反應(yīng)在SENSQUEST labcycler PCR 儀上進(jìn)行。反應(yīng)總體積為10 μl,包括DNA 模板(20 ng/μl)1 μl,Super TaqMix 5 μl,正反向引物(10 μmol/L)各1 μl,雙蒸水補(bǔ)足10 μl。PCR 反應(yīng)程序為94 ℃預(yù)變性5 min,35 個循環(huán)(94 ℃30 s,55 ℃30 s,72 ℃30 s),72 ℃延伸10 min。擴(kuò)增產(chǎn)物在8%非變性聚丙烯酰胺凝膠上80 V 恒電壓上電泳分離,用硝 酸銀法染色,拍照。
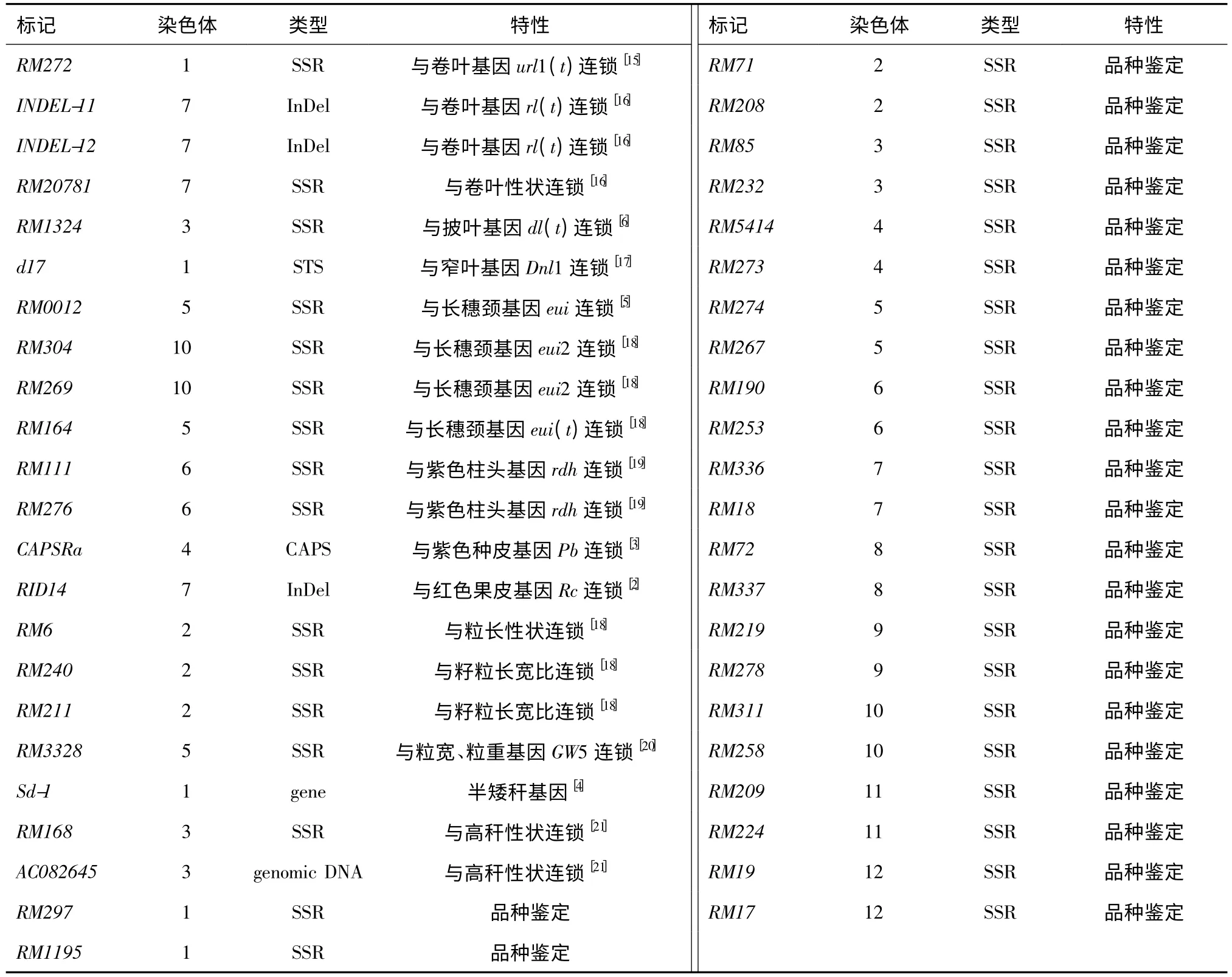
表2 45 個分子標(biāo)記的基本信息Table 2 Basic information on 45 molecular markers
1.3 數(shù)據(jù)分析
1.3.1 群體結(jié)構(gòu)分析 品種群體結(jié)構(gòu)分析應(yīng)用STRUCTURE 2.3.3 軟件[22],對標(biāo)準(zhǔn)品種總?cè)后w進(jìn)行基于數(shù)學(xué)模型的類群劃分。首先確定亞群數(shù),然后計算各品種歸屬第k 亞群的概率Q 值(第i 品種基因組變異源于第k 群體的概率)。在確定亞群數(shù)時,選擇混合模型和獨(dú)立等位基因頻率模型,依次設(shè)定亞群數(shù)目(K)為1 ~10,將MCMC(Markov chain monte carlo)開始時的不作數(shù)迭代(Length of burn-in period)設(shè)為100 000次,再將不作數(shù)迭代后的MCMC設(shè)為100 000次,然后依據(jù)似然值最大的原則選取一個合適的K 值。
1.3.2 關(guān)聯(lián)分析 與性狀相關(guān)的分子標(biāo)記的確認(rèn)使用TASSEL 2.1 軟件的GLM(General linear model)程序(http://www.maizegenetics.net/index.php?option = com_content&task = view&id =89&Itemid =119)。將STRUCTURE 2.3.3 軟件運(yùn)行后形成的Q值作為協(xié)變量,然后利用標(biāo)記變異分別對水稻38 個性狀的表型值逐一進(jìn)行回歸分析[23]。
2 結(jié)果與分析
2.1 水稻45 個品種38 個性狀的變異分析
從表3、表4 可以看出,24 個目測性狀的代碼分布范圍較廣,試驗材料在所分析的這些性狀上有較大的變異。在品種總?cè)后w中,14 個測量性狀變異幅度差異較大,其中性狀“穗:最長芒長度”的變異系數(shù)最大,為108.64%,其次為“穗:抽出度”(66.03%),“植株:單株穗數(shù)”、“莖稈:長度(不包括穗)”和“穗:每穗粒數(shù)”的變異系數(shù)均超過30%,劍葉長度和劍葉寬度的變異系數(shù)分別為20.57%和22.10%,“抽穗期”、“莖稈:粗細(xì)”、“穗:長度”、“谷粒:長度”、“糙米:長度”和“糙米:寬度”變異系數(shù)為13% ~20%,“谷粒:寬度”的變異系數(shù)最小,為12.20%。說明供試水稻標(biāo)準(zhǔn)品種間各性狀差異較大,具有較豐富的多樣性,可為水稻品種的測試提供很好的參照。
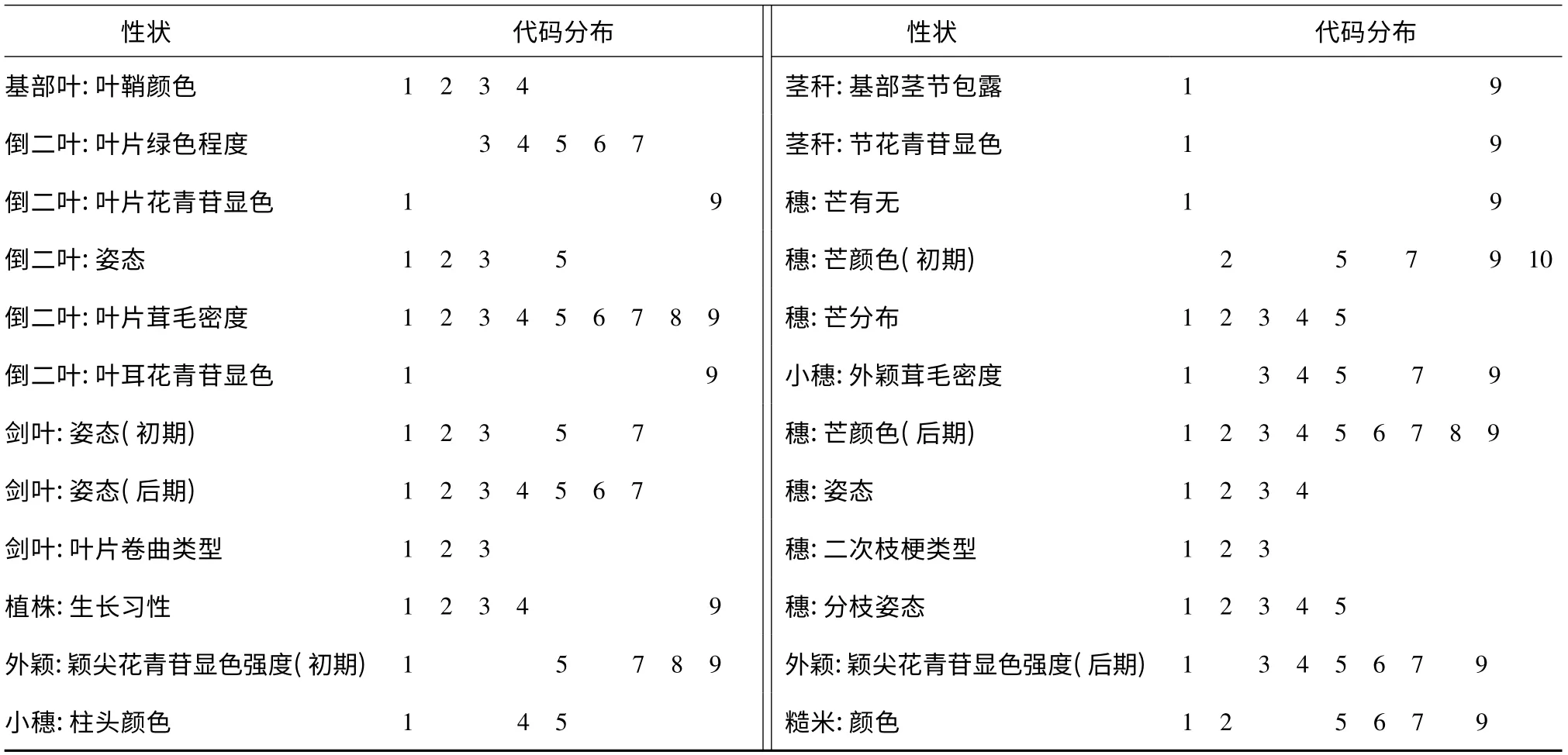
表3 45 個水稻品種的目測性狀Table 3 Visual test of 24 traits in 45 rice varieties

表4 45 個水稻品種的測量性狀Table 4 Measurement of 14 trait in 45 rice varieties
2.2 水稻品種總?cè)后w的結(jié)構(gòu)
利用STRUCTURE 2.3.3 軟件,對水稻品種總?cè)后w進(jìn)行結(jié)構(gòu)分析,發(fā)現(xiàn)對數(shù)似然函數(shù)值在亞群數(shù)K=9 時最大(圖1),因而確定該品種群體的適宜亞群數(shù)為9。確定亞群數(shù)為9 后,用STRUCTURE 2.3.3軟件計算出每個品種歸屬于9 亞群的后驗概率值(圖2)。
2.3 與性狀相關(guān)聯(lián)的分子標(biāo)記位點(diǎn)及其對表型變異的解釋率
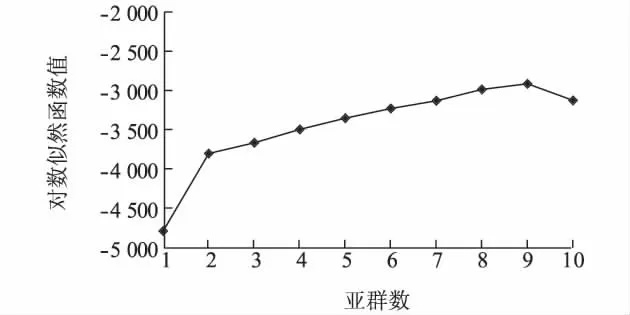
圖1 對數(shù)似然函數(shù)值lnP(D)和K 值隨亞群數(shù)的變化Fig.1 Changes of the log-likelihood function value lnP(D)and K value on the number of subgroups
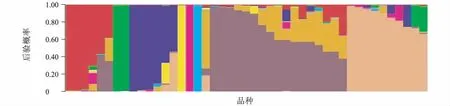
圖2 用STRUCTURE 2.3.3 軟件計算的品種總?cè)后w中每個水稻品種歸屬于9 亞群的后驗概率Fig.2 Posterior probability of each rice variety out of total variety population belonging to 9 subgroups calculated by STRUCTURE 2.3.3 software
對45 個位點(diǎn)與38 個性狀進(jìn)行關(guān)聯(lián)分析,共檢測到與23 個性狀相關(guān)聯(lián)(P <0.01)的20 個標(biāo)記(表5)。與“基部葉:葉鞘顏色”、“倒二葉:葉耳花青苷顯色”、“劍葉:葉片寬度”、“植株:生長習(xí)性”、“抽穗期”、“莖稈:長度(不包括穗)”、“穗:長度”、“穗芒有無”、“穗:芒顏色(初期)”、“穗:芒顏色(后期)”、“穗:分枝姿態(tài)”、“穗:每穗粒數(shù)”相關(guān)聯(lián)的標(biāo)記各有1 個,與“外穎:穎尖花青苷顯色強(qiáng)度(初期)”、“小穗:柱頭顏色”、“穗:最長芒長度”、“穗:抽出度”、“外穎:穎尖花青苷顯色強(qiáng)度(后期)”、“谷粒:長度”、“糙米:長度”相關(guān)聯(lián)的標(biāo)記各有2個;與“莖稈:節(jié)花青苷顯色”、“小穗:外穎茸毛密度”相關(guān)聯(lián)的標(biāo)記各有3 個;與“倒二葉:葉片綠色程度”相關(guān)聯(lián)的標(biāo)記有4 個;與“糙米:顏色”相關(guān)聯(lián)的標(biāo)記有5 個。其中標(biāo)記RM111 與“基部葉:葉鞘顏色”、“倒二葉:葉片綠色程度”、“外穎:穎尖花青苷顯色強(qiáng)度(初期)”、“小穗:柱頭顏色”和“抽穗期”5 個性狀相關(guān)聯(lián),與“基部葉:葉鞘顏色”和“小穗:柱頭顏色”2 個性狀極顯著(P <0.01)相關(guān),對5個性狀變異的解釋率分別為48.47%、26.09%、24.76%、39.31%和21.62%;標(biāo)記AC082645 與“外穎:穎尖花青苷顯色強(qiáng)度(初期)”、“莖稈:節(jié)花青苷顯色”、“穗:芒顏色(初期)”、“穗:芒顏色(后期)”、“外穎:穎尖花青苷顯色強(qiáng)度(后期)”、“糙米:顏色”和“穗:抽出度”7 個性狀相關(guān)聯(lián),與“外穎:穎尖花青苷顯色強(qiáng)度(初期)”極顯著相關(guān),分別解釋7個 性 狀 變 異 的 53.81%、44.46%、38.92%、40.30%、38.86%、59.37%和26.29%;標(biāo)記RM278與“莖稈:長度(不包括穗)”、“小穗:外穎茸毛密度”、“穗:每穗粒數(shù)”和“谷粒:長度”4 個性狀相關(guān)聯(lián),可以解釋4 個性狀變異的14.74%、12.73%、21.31%和16.32%。RM297 與“谷粒:長度”、“糙米:長度”和“糙米:顏色”3 個性狀相關(guān)聯(lián),分別解釋3 個性狀變異的33.48%、36.55%、56.97%;標(biāo)記RM85 與“倒二葉:葉片綠色程度”和“糙米:顏色”2個性狀相關(guān)聯(lián),與“倒二葉:葉片綠色程度”極顯著相關(guān),對2 個性狀變異的解釋率分別為55.72%和35.76%;RM253 也與“倒二葉:葉片綠色程度”極顯著相關(guān);RM258 與“倒二葉:葉耳花青苷顯色”、“糙米:長度”2 個性狀相關(guān)聯(lián);RM276 與“小穗:柱頭顏色”、“小穗:外穎茸毛密度”2 個性狀相關(guān)聯(lián);RM232與“莖稈:節(jié)花青苷顯色”、“穗:最長芒長度”2 個性狀相關(guān)聯(lián);RM336 與“穗:最長芒長度”、“穗:抽出度”2 個性狀相關(guān)聯(lián);RM168 與“穗:長度”相關(guān)聯(lián);與“莖稈:節(jié)花青苷顯色”相關(guān)聯(lián)的標(biāo)記中RM304 和RM232 與“莖稈:節(jié)花青苷顯色”極顯著相關(guān),其變異的解釋率分別達(dá)到67.19% 和48.94%;標(biāo)記RM3328 與“植株:生長習(xí)性”極顯著相關(guān),變異的解釋率56%。
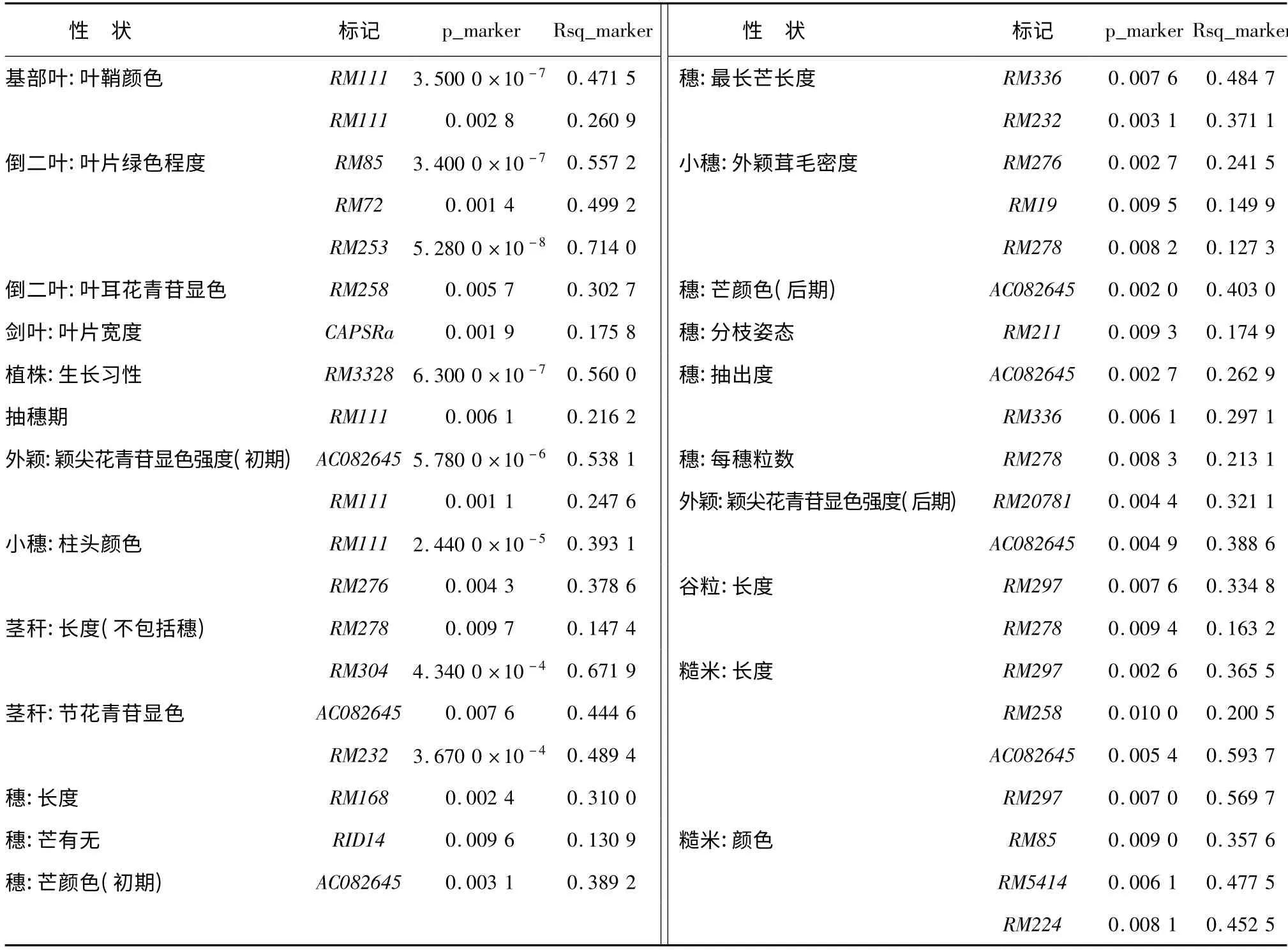
表5 與性狀顯著相關(guān)的標(biāo)記位點(diǎn)及其對表型變異的解釋率Table 5 Marker loci significantly associated with traits and the contributions to phenotypic variation
3 討論
本研究在45 個水稻品種中共檢測到20 個與23 個DUS 測試性狀相關(guān)聯(lián)的分子標(biāo)記。其中AC082645 標(biāo)記同時與7 個性狀關(guān)聯(lián),RM111 標(biāo)記同時與5 個性狀關(guān)聯(lián),RM278 同時與4 個性狀關(guān)聯(lián),RM297 同時與3 個性狀關(guān)聯(lián),其它16 個標(biāo)記與1 個或2 個性狀關(guān)聯(lián)。與AC082645 相關(guān)聯(lián)的7 個性狀中,6 個不同器官部位與顏色有關(guān)的性狀“莖桿:節(jié)花青苷顯色”、“外穎:穎尖花青苷顯色強(qiáng)度(初期)”、“外穎:穎尖花青苷顯色強(qiáng)度(后期)”、“穗:芒顏色(初期)”、“穗:芒顏色(后期)”、“糙米:顏色”與之相關(guān)聯(lián),其中與“外穎:穎尖花青苷顯色強(qiáng)度(初期)”達(dá)到了極顯著相關(guān),對該性狀的解釋率達(dá)到53.81%。說明AC082645 位點(diǎn)與控制水稻不同器官部位的花色素苷基因有關(guān)。與RM111 相關(guān)聯(lián)的5 個性狀中,“基部葉:葉鞘顏色”、“倒二葉:葉片綠色程度”、“外穎:穎尖花青苷顯色強(qiáng)度(初期)”、“小穗:柱頭顏色”均是花色素基因控制的性狀。RM111 與“基部葉:葉鞘顏色”和“小穗:柱頭顏色”兩性狀達(dá)到極顯著相關(guān),對兩性狀的解釋率分別為47.15%和39.31%。中國水稻研究所范方軍等[24]研究發(fā)現(xiàn)水稻第6 染色體短臂色素原基因C與RM111 和RM253 連鎖。韓磊等[19]對rdh 紫色柱頭定位群體的研究結(jié)果顯示,位于水稻第6 染色體的RM276、RM253 以及RM111 標(biāo)記與rdh 紫色柱頭基因有連鎖關(guān)系。本研究結(jié)果也顯示RM253 與“倒二葉:葉片綠色程度”極顯著相關(guān),該標(biāo)記對性狀解釋率達(dá)到了71.4%;RM276 與“小穗:柱頭顏色”相關(guān)聯(lián),與前人的研究結(jié)果一致。本研究結(jié)果還顯示,RM85、RM72 與“倒二葉:葉片綠色程度”相關(guān)聯(lián),RM258 與“倒二葉:葉耳花青苷顯色”相關(guān)聯(lián),RM304、RM232 與“莖稈:節(jié)花青苷顯色”相關(guān)聯(lián),RM20781 與“外穎:穎尖花青苷顯色強(qiáng)度(后期)”相關(guān)聯(lián),RM85、RM5414、RM224 和RM297 與“糙米:顏色”相關(guān)聯(lián),其中RM85 與“倒二葉:葉片綠色程度”和RM304、RM232 與“莖稈:節(jié)花青苷顯色”達(dá)到極顯著相關(guān),這3 個標(biāo)記對相關(guān)性狀的解釋率分別為55.72%、67.19%和48.94%。
與AC082645 相關(guān)聯(lián)的性狀“穗:抽出度”,實際也就是育種上常用的穗頸性狀。1981 年,Rutger 和Carnahan 在粳稻雜交后代中發(fā)現(xiàn)1 個最上節(jié)間明顯伸長的高稈隱性突變體,并將控制該性狀的基因命名為eui(Elongateduppermost internode)[25]。隨后,國內(nèi)外的一些學(xué)者Kita 等[26]、Librojo 等[27]、吳玉良等[28]、楊蜀嵐等[29]對eui 基因進(jìn)行了初步定位。張所兵等[5]將長穗頸eui 基因定位在第5 染色體上的RM3673 和RM0012 之間。馬玉銀等[21]運(yùn)用一個高稈突變體對水稻的隱性高稈基因進(jìn)行研究,發(fā)現(xiàn)1個新的隱性高稈基因lc3 與第3 條染色體上RM168與AC082645 共分離。本研究中AC082645 與“穗:抽出度”相關(guān)聯(lián),與前人報道的相近。
在已有報道中CAPSRa 與紫色種皮基因Pb 連鎖,RID14 與紅色種皮基因Rc 連鎖[2-3],但在本研究中未發(fā)現(xiàn)這兩個標(biāo)記與“糙米:顏色”相關(guān)。這可能是由于CAPSRa 只與紫皮和白皮性狀連鎖,RID14只與紅皮和白皮性狀連鎖。而在本研究中,水稻DUS 測試指南中將“糙米:顏色”性狀分為白色、淺棕色、棕色斑駁、深棕色、淺紅色、紅色、紫色斑駁、紫色和紫黑色9 級,顏色分級過細(xì)致使CAPSRa 和RID142 標(biāo)記未檢測出與“糙米:顏色”性狀的關(guān)聯(lián)。RID14 是楊杰等[2]根據(jù)普通野生稻(O. rufipogon)紅色果皮基因Rc 的cDNA 序列設(shè)計的引物而開發(fā)的標(biāo)記,該標(biāo)記與野生稻存在較大的相關(guān)性。本研究中發(fā)現(xiàn)RID14 與“穗:芒有無”顯著相關(guān),而芒的有無是野生稻和栽培稻的區(qū)別之一,野生稻都是有芒的,栽培稻中有的有芒,有的無芒,所以RID14 標(biāo)記可以用于水稻的起源研究中。
除上述相關(guān)聯(lián)的標(biāo)記與性狀外,本研究還發(fā)現(xiàn)RM3328 與“植株:生長習(xí)性”極顯著相關(guān),該標(biāo)記對性狀的解釋率達(dá)到了56.00%;以及RM278 與“莖稈:長度(不包括穗)”相關(guān)聯(lián)的SSR 標(biāo)記,RM336 與“穗:抽出度”相關(guān)聯(lián)的SSR 標(biāo)記,RM168 與“穗:長度”相關(guān)聯(lián)的SSR 標(biāo)記,RM232、RM336 與“穗:最長芒長度”相關(guān)聯(lián)的SSR 標(biāo)記,RM278 與“谷粒:長度”相關(guān)聯(lián)的SSR 標(biāo)記,RM258、RM297 與“糙米:長度”相關(guān)聯(lián)的SSR 標(biāo)記,RM278 與“穗:每穗粒數(shù)”相關(guān)聯(lián)的SSR 標(biāo)記,RM278、RM276、RM19 與“小穗:外穎茸毛密度”相關(guān)聯(lián)的SSR 標(biāo)記。在秈、粳稻的許多差別中,谷粒和穎毛的差別最為明顯,秈稻谷粒細(xì)長,穎毛短而散生;粳稻谷粒寬而厚,穎毛長而密生。本研究中用于品種鑒定的部分與水稻DUS 測試性狀存在緊密關(guān)聯(lián)的SSR 標(biāo)記,可以為水稻的秈粳分類研究提供參考信息。
[1] UPOV. Document TG/1/3 General introduction to the examination of distinctness,uniformity and stability and the development of harmonized descriptions of new varieties of plants[S].2002.
[2] 楊 杰,王 軍,曹 卿,等.雜草稻紅色果皮基因的功能標(biāo)記開發(fā)[J]. 分子植物育種,2009,7(4):721-726.
[3] 王彩霞,舒慶堯. 水稻紫色種皮基因Pb 的精細(xì)定位與候選基因分析[J]. 科學(xué)通報,2007,52(21):2517-2523.
[4] CHO Y G,DARVASI A. Optimu spacing of genetic markers for determining linkage between marker loci and quantitative trait loci[J]. Theor Appl Genet ,1994,89:54-55.
[5] 張所兵,朱 鎮(zhèn),趙 凌,等. 水稻長穗頸基因eui 緊密連鎖SSR 標(biāo)記獲得[J]. 遺傳,2007,29(3):365-370.
[6] 王 楠,趙芳明,凌英華,等. 一個水稻披葉突變體的遺傳分析和基因定位分子植物育種[J]. 分子植物育種,2007,5(1):54-58.
[7] 高 虹,王嘉宇,姜樹坤,等. 水稻粒長基因GL3 的遺傳分析和分子標(biāo)記定位[J]. 植物生理學(xué)通訊,2010,46(3):236-240.
[8] LI X B,YAN W G,AGRAMA H,et al. Mapping QTLs for improving grain yield using the USDA rice mini-core collection [J].Planta,2011,234:347-361.
[9] 陳 蘭,張 紅,張啟武,等. 水稻6 個異交相關(guān)性狀的SSR 關(guān)聯(lián)分析[J]. 南京農(nóng)業(yè)大學(xué)學(xué)報,2012,35(2):1-9.
[10] 文自翔,趙團(tuán)結(jié),鄭永戰(zhàn),等. 中國栽培和野生大豆農(nóng)藝及品質(zhì)性狀與SSR 標(biāo)記的關(guān)聯(lián)分析Ⅰ:群體結(jié)構(gòu)及關(guān)聯(lián)標(biāo)記[J].作物學(xué)報,2008,34(7):1169-1178.
[11] BRESEGHELLO F,SORRELLS M E. Association mapping of kernel size and milling quality in wheat(Triticum aestivum L.)cultivars[J]. Genetics,2006,172:1165-1177.
[12] 武玉國,吳承來,秦保平,等. 黃淮冬麥區(qū)175 個小麥品種的遺傳多樣性及SSR 標(biāo)記與株高和產(chǎn)量相關(guān)性狀的關(guān)聯(lián)分析[J].作物學(xué)報,2012,38(6):1018-1028.
[13] YAN W G,LI Y,AGRAMA H A,et al. Association mapping of stigma and spikelet characteristics in rice(Oryza sativa L.)[J].Molecular Breeding,2009,24:277-292.
[14] HUANG X H,WEI X H,SANG T,et al. Genome-wide association studies of 14 agronomic traits in rice landraces[J]. Nature Genetics,2010,42:961-969.
[15] 余 東,吳海濱,楊文韜,等. 水稻單側(cè)卷葉突變體B157 遺傳分析及基因初步定位[J]. 分子植物育種,2008,6(2):220-226.
[16] 施勇烽,陳 潔,劉文強(qiáng),等. 一個新的水稻卷葉突變體的遺傳分析與基因定位[J]. 中國科學(xué)(C 輯:生命科學(xué)),2009,39(4):407-412.
[17] 曾生元,郭 旻,李 敏,等. 一個水稻動態(tài)窄葉突變體的鑒定和基因定位[J]. 科學(xué)通報,2010,55(21):2106-2111.
[18] 陳 瑞,程在全,黃興奇,等. 水稻優(yōu)良性狀控制基因的定位進(jìn)展及其在染色體上的分布[J]. 遺傳,2007,29(4):399-412.
[19] HAN L,ZHANG T,XU J D,et al. Genetic analysis and gene mapping of purple stigma in rice[J] . Acta Genetica Sinica,2006,33(7):642 -646.
[20] WENG J F,GU S H,WAN X Y,et al. Isolation and initial characterization of GW5,a major QTL associated with rice grain width and weight[J]. Cell Research,2008,18(12):1199-1209.
[21] 馬玉銀,李 磊,李育紅,等. 一個新的水稻隱性高稈突變體的遺傳分析和基因定位[J]. 中國農(nóng)業(yè)科學(xué),2008,41(12):3967-3973.
[22] PRITCHARD J K,WENA X,F(xiàn)ALUXH D. Documentation for structure software:version 2.3[CP/OL]. Chicago:Department of Human Genetics,University of Chicago,2010[2012-09-12].http://pritch. bsd. uchicago. edu/structure_software/release_versions/v2.3.3/structure_doc.pdf.
[23] BRADBURY P J,ZHANG Z,KROON D E ,et al. TASSEL:software for association mapping of complex traits in diverse samples[J]. Bioinformatics,2007,23:2633-2635.
[24] 范方軍,樊葉楊,莊杰云,等. 水稻色素原基因C 的精細(xì)定位[J]. 中國水稻科學(xué),2007,21(5):454 -458.
[25] TUTGER J N,CARNAHAN H L. A forth genetic element to facilitate hybrid cereal production—a recessive tall in rice[J]. Crop Science,1981,21:373-376.
[26] KITA M M. F1 Interaction of eui gene for the elongation of uppermost internode and some genes for elongation of internode[J].Japanese Journal of Plant Breeding,1983,33 (11):124-125.
[27] LIBROJO L,KHUSH G S. Chromosomal location of some mutant genes through the use of primary trisomics in rice[C]. Manila:Island Press Inc,1986:249-255.
[28] 吳玉良,何祖華,董繼新,等. 水稻株高基因eui 的初步定位[J]. 中國水稻科學(xué),1998,12(2):119-120.
[29] 楊蜀嵐,馬洪麗,張書標(biāo),等. 水稻長穗頸高稈隱性基因eui2的分子標(biāo)記和定位[J]. 福建農(nóng)業(yè)大學(xué)學(xué)報,2002,31(4):480-483.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
