魚源小腸結腸炎耶爾森菌5種毒力基因的檢測和分析
2013-02-24 01:09:52茍小蘭
中國獸醫雜志 2013年2期
關鍵詞:檢測
王 利,茍小蘭
(西南民族大學生命科學與技術學院 動物遺傳育種學國家民委-教育部重點實驗室,四川 成都 610041)
小腸結腸炎耶爾森菌(Yersiniaenterocolitica)屬于耶爾森菌屬,腸桿菌科。小腸結腸炎耶爾森菌廣泛存在于自然界中,是人、畜、魚等共患的致病菌,可引發人和動物患胃腸炎和腹瀉,還可引起關節炎、結節性紅斑,嚴重時可引起敗血癥,造成死亡。從20世紀80年代開始,研究者用體外法測定小腸結腸炎耶爾森菌的毒力以代替昂貴的動物試驗。體外法主要包括:小腸結腸炎耶爾森菌的自凝性、血清抵抗性和毒力質粒的測定等[1-2]。至今為止,對小腸結腸炎耶爾森菌的毒力的研究已經取得了一些進展[3-4]。對其致病機制有了初步的認識,但仍有許多的問題有待深入研究。隨著分子生物學技術的發展,聚合酶鏈反應(PCR)已經用于檢測小腸結腸炎耶爾森菌毒力基因,迅速對該菌進行毒力鑒定,甚至在毒力質粒丟失后仍然可以預測該菌的致病力[5]。本試驗采用PCR方法檢測和分析了魚源小腸結腸炎耶爾森菌5種重要的毒力基因:毒力活化因子基因(vir F)、粘附素基因(yad A)、粘附侵襲位點基因(ail)、毒力島基因(HPI-int)和耐熱腸毒素基因(ystB)。這有利于為深入探討魚源小腸結腸炎耶爾森菌的毒力和致病性積累科學資料。
1 材料與方法
1.1 菌種來源 小腸結腸炎耶爾森菌分離自四川某胭脂魚養殖場,以甘油菌的形式保存于本實驗室-80℃冰箱,備用。
1.2 主要試劑 改良Y培養基和CIN-1培養基,購自杭州天和微生物試劑有限公司;細菌基因組DNA提取試劑盒,購自TIANGEN BIOTECH(BEIJING);TaqDNA聚合酶、dNTP、DNA Marker(2000)等,購自寶生物工程(大連)有限公司。
1.3 方法
1.3.1 引物設計 參考文獻[6],設計小腸結腸炎耶爾森菌5對毒力基因(ail、yad A、virF、ystB和 HPI-int)的引物,詳細見表1。引物由上海生工生物工程技術服務有限公司合成。

表1 5對毒力基因引物序列
1.3.2 DNA模板制備 采用兩種方法制備小腸結腸炎耶爾森菌DNA模板。第一方法是煮沸法,主要步驟是:取營養平板上生長的單個菌落加入含50μL dd H2O的1.5 m L離心管中,混勻后放入-80℃冰箱中過夜,取出后立即放入沸水中煮10 min,12 000 r/min離心5 min,取上清液作為模板,-20℃凍存備用。第二種方法是用細菌基因組DNA提取試劑盒,具體步驟參照試劑盒說明書進行。
1.3.3 PCR反應 PCR反應的體系:dd H2O 8.5 μL,2×TaqPCR Master Mix 12.5μL,上游引物和下游引物(濃度均為10μmol/L)各1μL,模板DNA 2 μL,共25μL。ail和HPI-int毒力基因PCR擴增反應條件為:94℃5 min;94℃60 s,50℃60 s,72℃60 s,35個循環,72℃10 min。virF毒力基因PCR擴增反應條件為:94℃5 min;94℃60 s,48℃60 s,72℃60 s,35個循環,72℃10 min。另外,兩種毒力基因(ystB和yad A)則采用梯度PCR反應進行優化:94℃5 min;94℃60 s,退火溫度分別為45℃、50℃、55℃和60℃,35個循環,72℃10 min。PCR結束后,取PCR產物5μL,進行1.2%瓊脂糖凝膠電泳,80V電壓,電泳30 min,凝膠成像儀下拍照。
1.3.4 HPI-int基因測序和序列分析 HPI-int基因PCR反應采用50μL體系:dd H2O 17μL,2×TaqPCR Master Mix 25μL,上游引物和下游引物(濃度均為10μmol/L)各2μL,模板DNA 4μL。PCR擴增反應條件為:94℃ 5 min;94℃ 60 s,50℃ 60 s,72℃60 s,35個循環,72℃10 min。PCR產物經1.2%瓊脂糖凝膠電泳,檢測出單一條帶后,送交上海生工生物工程技術服務有限公司,采用ABI3730測序儀進行正反向測序。所得序列提交到NCBI的GenBank數據庫,并采用生物信息學軟件DNAStar和Blastn等對該序列進行分析。
2 結果
2.1 毒力基因檢測 分別用兩種方法制備的DNA作為模板,用PCR方法檢測小腸結腸炎耶爾森菌5種毒力基因(ail、yad A、virF、ystB和 HPI-int),得到的結果相似,無明顯差異。ail基因目的條帶大小約為351 bp,HPI-int基因條帶大小約為714 bp,而virF基因目的條帶大小約為561 bp,這3種基因所得的PCR擴增產物大小與試驗預期結果一致,結果詳見圖1。另外兩種基因(ystB和yad A)在反復優化PCR條件后都未擴增出目的條帶,表明該菌株未攜帶ystB和yad A這兩種毒力基因。
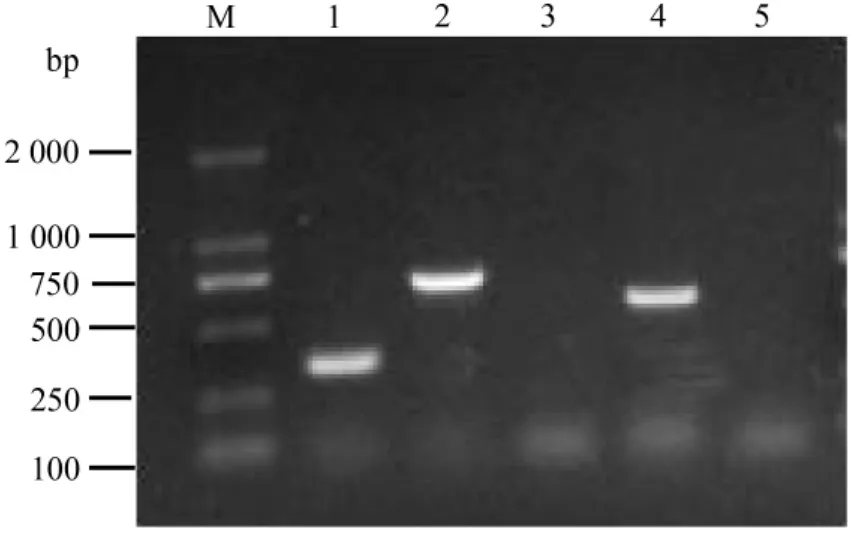
圖1 毒力基因的擴增1:ail;2:HPI-int;3:yad A;4:vir F;5:ystB;M:DNA Marker分子量
2.2 HPI-int基因序列分析 HPI-int基因分別使用正反向引物測序,采用DNAStar軟件中的Seqman對正反向序列進行組裝、拼接,最終所得序列長度為722 bp,將該序列提交到 NCBI,GenBank序列號為JX041513。采用NCBI的Blastn工具對該序列進行比對,結果顯示,該序列與小腸結腸炎耶爾森菌強毒力島基因(Yersiniaenterocoliticaleft arm of the highpathogenicity island Score,GenBank 序 列 號 為AJ132668.1)序列的相似性為99% (705/710),因此表明HPI-int基因的測序結果和預期相符合。
3 討論
耶爾森菌的致病因素主要包括:毒力質粒編碼的分泌系統、外膜蛋白、侵襲性與毒素、鐵攝取系統和超抗原等方面[7]。致病性小腸結腸炎耶爾森菌的致病因素主要是該菌所具有的特殊染色體基因、毒力質粒和菌毛。ail、HPI-int、yst A、ystB和 HPI毒力基因位于染色體上,而yad A和virF毒力基因位于質粒上。致病性的耶爾森菌可分為低致病性和高致病性兩類。這種分類與染色體上是否攜帶有 HPI毒力島有關,因為HPI是高毒力細菌表型表達的必需條件。這個染色體上的片段涉及生物合成、調節和耶爾森菌含鐵鐵載體的轉運作用。在小腸結腸炎耶爾森菌中,毒力島只出現于高致病性的小腸結腸炎耶爾森菌生物1B型 ,而低致病性的則無[4]。
至今為止,對小腸結腸炎耶爾森菌的毒力基因已經進行了一些研究[8-9]。典型的致病性小腸結腸炎耶爾森菌攜帶ail、yst A、yad A基因和virF毒力基因。致病性菌株不攜帶ystB,而非致病性菌株不攜帶ail、yst A、yad A、vir F。ystB主要為生物1A型小腸結腸炎耶爾森菌攜帶的編碼一種性質類似于yst A的耐熱性腸毒素。ystB僅存在于生物1A型的菌株,而且在這個生物型中,目前資料表明這類菌株通常都是非致病性菌株。81株生物1A型小腸結腸炎耶爾森菌均未檢測出ail、virF、yst A基因和ystC基因[10]。王鑫等(2005)運用PCR和DNA探針雜交方法檢測了致病性和非致病性小腸結腸炎耶爾森菌攜帶的ystB基因[11]。48株致病性小腸結腸炎耶爾森菌均不攜帶該基因。98株非致病性小腸結腸炎耶爾森菌中有52株攜帶ystB基因,占53.06%。這表明ystB基因僅存在于部分生物1A型非致病性小腸結腸炎耶爾森菌,而不存在于致病性菌株中。以上多個研究結果基本一致。本試驗通過多次優化PCR條件也未檢測出ystB基因,這提示該菌株很可能為致病性小腸結腸炎耶爾森菌。
裴耀文等(2011)發現3株致病性小腸結腸炎耶爾森菌株的毒力基因檢測中均表現為ail和yst A陽性,而yad A、virF毒力基因缺失[12]。吉林省致病性小腸結腸炎耶爾森菌血清型毒力基因的分布情況表明,血清型O:3和O:9小腸結腸炎耶爾森菌毒力基因的分布主要為ail+、yst A+、ystB-、yad A+、virF+型,其次是ail+、yst A+、ystB-、yad A-、vir F-型[13]。南京地區不同來源的123株小腸結腸炎耶爾森菌致病生物型菌株,均包含染色體上的毒力基因ail[14]。因此,ail與致病生物型之間存在明顯關聯。Zheng等(2008)用PCR方法檢測了從腹瀉病人糞便中分離出160株致病性小腸結腸炎耶爾森菌的毒力基因。檢出率分別 為:ail(94%)、inv(100%)、yst A (93%)、ystB(7.5%)、ystC(5%)、yad A(89%)和virF(82%)[5]。該結果表明,并非所有的致病性小腸結腸炎耶爾森菌都攜帶了能引起疾病的染色體和質粒上的全部毒力基因。由此推測,其中一些缺乏某種毒力基因的菌株可能具有其他的、未知的毒力標記,從而在致病性小腸結腸炎耶爾森菌的多種發病機理中發揮著重要的作用。以上報道的結果存在一些較小的差異,可能是由于不同地區、不同菌株的差異或毒力基因的檢測片段的差異等原因。
本試驗中檢測出了ail、vir F和HPI-int,而未檢測到yad A,這與以上報道基本相一致。這也提示該菌具有致病性。HPI-int基因的測序結果分析表明,該序列與小腸結腸炎耶爾森菌強毒力島基因的相似性達到99%,從而驗證了本試驗HPI-int基因檢測的正確性,也進一步說明該菌株具有較強的致病性。yad A基因未被檢出到可能是由于該基因位于質粒上,在實驗室分離培養過程中菌株多次傳代及長時間平板上保存后都很容易丟失質粒。質粒上的毒力基因與致病生物型之間的關聯性要略低一些,因此依賴于質粒毒力基因檢測通常是存在較大誤差。致病性耶爾森菌遺傳背景復雜,染色體DNA指紋分析具有多態性。在檢測細菌毒力基因時必須檢測染色體上的毒力因子,不能僅用毒力質粒的檢測。選擇染色體上的毒力基因時可優先選擇ail基因,同時實驗室應避免質粒丟失而使有毒株被誤判為無毒株。綜上所述,本試驗運用PCR方法檢測了小腸結腸炎耶爾森菌株的5種毒力基因。本菌株檢測并分析了染色體上的毒力島(HPI-int)基因,因此推測它是強毒力株,具有較強的致病性。下一步,將進行本菌株對多種魚類的致病性試驗,深入研究該菌株的毒性及其危害。
[1]Stern N J,Kotula A W,Pierson MD.Virulence prediction ofYersiniaenterocoliticaby pyrolysis gas-liquid chromatography[J].Appl Environ Microbiol,1980,40(3):646-651.
[2]Lee W H,Smith R E,DamaréJ M,etal.Evaluation of virulence test procedures forYersiniaenterocoliticarecovered from foods[J].J Appl Bacteriol,1981,50(3):529-539.
[3]Fabrega A,Vila J.Yersiniaenterocolitica:pathogenesis,virulence and antimicrobial resistance[J].Enferm Infecc Microbiol Clin,2012,30(1):24-32.
[4]Kot B,Piechota M,Jakubczak A.Analysis of occurrence of virulence genes amongYersiniaenterocoliticaisolates belonging to different biotypes and serotypes[J].Pol J Vet Sci,2010,13(1):13-19.
[5]Zheng H,Sun Y,Mao Z,Jiang B.Investigation of virulence genes in clinical isolates ofYersiniaenterocolitica[J].FEMS Immunol Med Microbiol,2008,53(3):368-374.
[6]王增國.我國部分地區攜帶ystB基因小腸結腸炎耶爾森菌的分子流行病學研究[D].鎮江:江蘇大學,2008.
[7]古文鵬,景懷琦.耶爾森菌致病機理研究[J].中國人獸共患病學報,2010,26(9):862-866.
[8]Drummond N,Murphy B P,Ringwood T,etal.Yersiniaenterocolitica:a brief review of the issues relating to the zoonotic pathogen,public health challenges,and the pork production chain[J].Foodborne Pathog Dis,2012,9(3):179-189.
[9]Schutz M,Weiss E M,Schindler M,etal.Trimer stability of Yad A is critical for virulence ofYersiniaenterocolitica[J].Infect Immun,2010,78(6):2677-2690.
[10]Bhagat N,Virdi J S.Distribution of virulence-associated genes inYersiniaenterocoliticabiovar 1A correlates with clonal groups and not the source of isolation[J].FEMS Microbiol Lett,2007,266(2):177-183.
[11]王鑫,邱海燕,肖玉春,等.小腸結腸炎耶爾森菌耐熱性腸毒B基因(ystB)初步研究[J].中國人獸共患病雜 志,2005,21(6):449-454.
[12]裴耀文,馮開軍,房玉英,等.山東省宿主動物中小腸結腸炎耶爾森菌分布狀況及毒力研究[J].醫學動物防制,2011,27(2):104-106.
[13]顧峰,張貴軍,劉亞東,等.吉林省耶爾森菌O:3和O:9血清型主要毒力基因分布調查[J].中國地方病防治雜志,2006,21(6):340-342.
[14]許文炯,丁潔,陳曉蔚,等.小腸結腸炎耶爾森菌主要毒力基因分析[J].中國人獸共患病學報,2007,23(7):675-677.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
