異常ABO表型遺傳的分子背景研究
2013-03-01 01:42:19喻瓊蘇宇清綜述鄧志輝審校
實驗與檢驗醫學 2013年1期
喻瓊,蘇宇清綜述,鄧志輝審校
(深圳市血液中心,廣東深圳518035)
異常ABO表型遺傳的分子背景研究
喻瓊,蘇宇清綜述,鄧志輝審校
(深圳市血液中心,廣東深圳518035)
ABO血型;ABO基因;CisAB;血型開米拉;親權鑒定
ABO系統是人類最早發現的血型系統,其血清學分型簡便、穩定。1925年Bernstein確定ABO血型遺傳學說之后,開始被應用于法醫學上真正科學意義的親權關系鑒定。
ABO血型的遺傳符合經典的遺傳規律,直到1964年cis-AB血型的發現,人們才開始進一步研究血型的遺傳規律。到目前為止,國內外陸續報道了數個罕見家系,在DNA多態性分析結果顯示不能排除或肯定其親權關系的家族成員中,ABO血型表型遺傳不符合孟德爾遺傳規律,這引起了研究者對經典遺傳規律的質疑。經過血型工作者對ABO血型的分子遺傳背景深入研究,已能從ABO基因水平解釋此異常現象。
1 ABO血型分子生物學結構
人類ABO基因定位于9q34.1-9q34.2,包含7個外顯子和6個內含子。編碼ABO血型系統糖基轉移酶的3個主要的等位基因(A1、B1、O)首次被Yamamoto等于1990年克隆測序。A、B基因產物分別編碼控制A型或B型糖基轉移酶,負責將相應的糖基連到H物質上,表現出不同抗原特異性。而O基因編碼一種無轉移酶活性功能的蛋白,不能將糖基連接到H物質上,抗原表現為O型。ABO基因的核苷酸序列高度保守,與一致序列A101等位基因的序列相比:O1基因的差異只是在第261核苷酸G缺失,這引起閱讀框移動,肽鏈合成在117位氨基酸終止;B基因僅在297A>G、526C>G、657C>T、703G>A、796C>A、803G>C、903G>A和1096G>A等8個堿基位置有改變,導致糖基轉移酶多肽鏈上有4個氨基酸發生了替代[1]。到目前為止,已發現百余種ABO亞型基因的序列[2],這些序列多態性的等位基因構成了ABO血型分子生物學分型的基礎。
ABO血型遺傳有常染色體顯性和隱性兩種方式:A、B是顯性基因,O是隱性基因,基因型為AB的血型表型為AB型,基因型為AA或AO的血型表型均為A型,只有基因型為OO的人的血型表型為O。
2 親權鑒定中異常ABO表型的ABO分子機制
在日常司法親權鑒定和臨床血型鑒定過程中,已出現STR遺傳多態性分析結果顯示不能排除或肯定其親權關系的家族成員中,ABO血型遺傳與經典孟德爾遺傳法則出現矛盾。結合我們實驗室多年的工作,匯總文獻資料,目前親權鑒定中造成異常ABO表型遺傳原因分為5大類。
2.1 ABO基因的重組和轉換ABO等位基因通過交換或顛換重組等方式形成各個亞型基因[3]。A、B型的特異性抗原是由ABO基因第7外顯子編碼的相應糖基轉移酶決定的。在減數分裂時期,B和O等位基因會融合雜交,這一基因的雜交重組多發生于第6內含子中,重組后的雜交基因表現為A基因活性。這是因為第7外顯子之前的核苷酸為B基因來源,而第7外顯子為O基因來源,這部分的基因序列與第6外顯子nt261位G未有缺失的A等位基因很相似,故表現為A型。我國中山大學法醫學系曾在2000年報道[4]:母親的血型為O型,父親的血型為B型,小孩為A型,17個STR的DNA多態性系統的檢測結果可以肯定他們之間的親生關系。從現在掌握的ABO分子背景可以解釋這個現象:母親的B和O1等位基因發生了整段重組(de novo recombination),即B等位基因的外顯子和O1基因的外顯子7所組成一個雜合基因,因糖基轉移酶的特異性主要由第7外顯子的堿基序列組成,故表達成A型,見示意圖1。這種雜合體在日本人群中產生的發生率為0.009,在日本也有一例類似報道[5]。

圖1 B基因的外顯子6與O基因的外顯子7序列重組成A101等位基因的序列
2.2 ABO等位基因的順反式影響抗原的表達一些亞型的表達程度主要與該等位基因表達增強或連鎖遺傳有關,或者可以說ABO基因在表達過程中會相互影響。日本Ogasawara K等報道一個AB型的父親與O型的母親生育出AB型的小孩[5],遺傳圖譜見圖2。
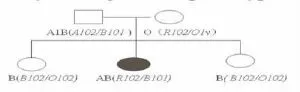
圖2 AB型父親與O型母親生育AB型小孩的遺傳圖譜
在此案例中發現一個雜合基因R102,它是A和O1v等位基因的重組,交換點在第6內含子。當R102/O雜交時因有261核苷酸位G的缺失,而表現為O型,而與B等位基因共同出現時基因加強作用(allelic enhancement)表現為AB型,說明等位基因的順反式能夠影響A或B抗原的表達。重組基因因為基因加強作用而使得在家系調查中ABO表現型異常的案例還有另外的報道:在一個瑞典家系中Ax04位基因遇到O01時表現為O型,而Ax04等位基因遇到O03時表現為A型[6];在一個韓國家系中Av與O01雜合時表現為O型,而Av與B雜合時表現為AwB型[7]。
2.3 孟買型或類孟買型由于H前體物質的全部或部分缺失,導致A或B抗原合成減少,血清學檢測中不出現凝集或弱凝集,造成ABO血型不能正確定型。決定H抗原的H基因與ABO基因是兩個單獨的系統,分子遺傳機制不同,此類案件中ABO基因是正常的。這樣往往會有ABO基因為A和/或B的但血型為O表型的父母親可能生育出正常的A或B血型的子代的報道[8,9]。
2.4 血型開米拉現象開米拉(Chimeras)是指一個個體中同時存在兩個或以上不同來源的細胞的現象。血型開米拉也常常會導致ABO血型遺傳學上的不相符[10,11]。人類血型開米拉較為罕見,常常是在ABO血型鑒定中檢測到同時存在兩種紅細胞而被發現的。我們實驗室在一個4歲中國漢族男孩,在外科手術前輸血的常規血型鑒定中,檢出為A3B3型,但其父母血型分別為O型、AB型。家屬對其親子關系產生懷疑,通過對其家系的調查得知4年前,他的母親自然分娩了一男一女,而女嬰在出生后的第二天因缺氧癥窒息而死。在其血液標本研究中發現,ABO血型基因、HLA-A、DR基因、和D8S1179,D13S317,和vWA位點均有三條單倍體存在[12]。這是一例的典型的血型開米拉嵌合體現象,可以很好解釋其遺傳方式。
2.5 包括Cis-AB型和B(A)型在內的雙重復合ABO糖基轉移酶型Cis-AB或B(A)血型基因當與A或B基因雜合由于A或B抗原的遮避,往往不會造成遺傳異常現象,而這兩個特殊的基因與O型基因雜合時會引起經典的遺傳規律的質疑:AB型的雙親能夠生育出O型的孩子或O型的母親或父親能夠生育出AB型的孩子。這種酶型是造成文獻報道最多的ABO表型遺傳異常的原因。
在ABO血型系統里有一類特殊的血型,經典血清學反應格局和相關的家系調查結果都會出現異常,這類血型同時具有A和B表達的ABO血型抗原,但又與正常的AB型不同,表達出來的A抗原和B抗原不會分離,能夠在子代中穩定遺傳下去,定義為Cis-AB型。還有一類在ABO血型正反定型中不符的血型——B(A)型,與不同的抗血清試劑反應有不同的結果,由于當時此類標本罕見報道,B(A)血型在文獻中籠統定義為:紅細胞與一些單克隆抗體試劑反應時,單克隆抗體在高能力識別B型細胞的同時,與某些A細胞也發生很弱凝集,而此類細胞與人源抗血清不反應。隨著當前高效價單克隆抗血清的廣泛使用,B(A)血清學表型常常易定為AB型。
隨著對人類ABO血型基因研究的深入,發現Cis-AB和B(A)表型的分子基因的基礎極其相似,以往認為血清學反應有所不同、異常表達原因不同的血型,可以歸為一類分析。
人們把普遍的A等位基因編碼的酶定為AAAA型,普遍的B等位基因編碼的酶定為BBBB型,這里的A和B代表A型糖基轉移酶特異性或B型糖基轉移酶特異性的氨基酸,AAAA型依次為176位精氨酸、235位甘氨酸、266位亮氨酸、268位甘氨酸和BBBB型為176位甘氨酸、235位絲氨酸、266位甲硫氨酸、268位丙氨酸。Cis-AB和B(A)型基因就是在常見的A等位基因或B等位基因
中,發生一個或二個堿基突變,使得編碼的糖基轉移酶具有雙重功能活性,能同時發生這兩種特異性糖連接的酶促反應:既可以催化A抗原合成,又能催化B抗原的形成[13-14]。從理論上說,對A酶或B酶催化活力起決定性作用的235位、266位、268位這3個氨基酸位點的隨機配合,應有8種酶型,目前國際上已報道了7種,只有BAA型目前還未見有報道。
AB型的父或母生出O型的子代,按照上述ABO雙重復合型酶的理論,已在分子基礎上闡明其原理,避免了對經典遺傳原則的懷疑,在法醫學物證鑒定上有特殊意義。
在1999年Yu LC報道的一個家系[15],見圖3。對此家系研究后發現,家庭成員帶有一個B(A)型基因,此基因位點是一個同時具有A和B糖基因轉移酶活性的基因,而不是兩個單獨的等位基因。而另外一條單倍體為O等位基因,故在基因水平可以解釋O型的雙親可以生育出AB型的子代。
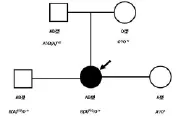
圖3 B(A)型家系遺傳圖譜
我國臺灣在2004年[16]報道了一例O型的父親生育出AB型的兒子(先證者A),16個短串聯重復位點計算父權概率達到了99.97%。經對兒子的ABO血型與基因進行分析,發現其一條單倍體為O1,另一條為CisAB04即B(A)700等位基因,父親遺傳O1等位基因給先證者,母親傳遞B(A)700等位基因給先證者,先證者血清學上AB表現型遮蓋了O型,因而表現為AB型。國內我實驗室[17]也在2007年報道了一例祖孫三代家系中,O型母親與AB型父親已生出了一個7歲O型男孩和一個AB型發生嚴重溶血病的新生兒,祖父母分別為O型和AB型,15個常染色體STR的基因型完全符合遺傳學規律,計算RCP值超過了99.99%。
本案例ABO血清學家系調查的結果,出現三個“異常”:(1)妻子0型、丈夫為AB型,卻生育了一個O型的孩子;(2)這對夫妻再次生育了AB型新生兒;(3)新生兒的祖父和祖母分別為AB型和A型,姑姑卻為O型。經ABO基因分析,在祖父、父親、新生兒中出現了一個新的ABO等位基因(見圖4),它與B101的差異,在于第7外顯子單個nt803C>G點突變,導致第268位AA發生Ala>Gly產生BBBA型糖基轉移酶,該酶具有A酶和B酶雙重復合催化功能。這個新等位基因在三代個體中能穩定地遺傳,比較其它人種,Cis-AB或B(A)型表型在亞洲人種較為常見,系統性研究中國人群雙重復合型ABO糖基轉移酶的抗原表達和分子基礎在法醫學有重要意義。
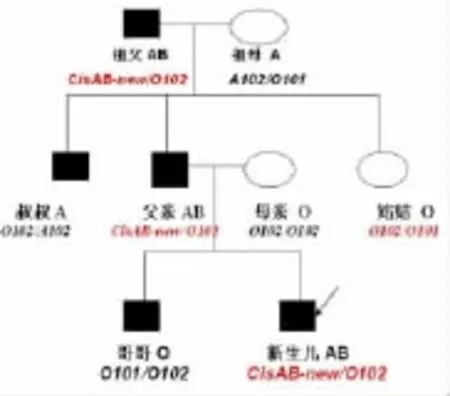
圖4 CisAB型家系遺傳圖譜
3 ABO血型基因在親權鑒定中的前景
ABO血型研究歷史悠久,分型標準化,群體數據詳盡,在法醫學、輸血醫學、人類學研究廣泛應用。ABO正反定型不符的標本時有發現[18],ABO血型基因的運用可以很好地解決血型鑒定困難問題。本文5類ABO表型遺傳異常的家系都表明:雖ABO表型不符合遺傳規律,但從ABO基因水平證實了ABO血型的遺傳完全符合孟德爾遺傳規律。在親權鑒定中發現ABO與其它多態性標記發生矛盾時,就必須進行ABO系統的DNA分型,才能確定其遺傳關系,不能單純以ABO血型的血清學分型結果違反孟德爾遺傳規律懷疑親生關系的結論。
系統地總結ABO表型遺傳異常的家系和案例,揭示和豐富ABO血型遺傳本質,避免親權鑒定中錯誤地排除親子關系,具有重要意義。
隨著多態性DNA的應用以及對ABO血型基因分子生物學研究的進一步深入,相信司法鑒定工作中可能會為更多ABO血型異常的案件釋疑。
[1]Chester MA,Olsson ML.The ABO Blood Group Gene:A Locus of Considerable Genetic Diversity[J].Transfus Med Rev,2001,15(3): 177-200.
[2]Patnaik SK,Helmberg W,Blumenfeld OO.BGMUT:NCBI dbRBC database of allelic variations of genes encoding antigens of blood group systems[J].Nucleic Acids Res,2012,40(1):1023-1029.
[3]Suzuki K,Iwata M,Tsuji H,et al.A de novo recombination in the
ABO blood group gene and evidence for the occurrence of recombination products[J].Hum Genet,1997,99(4):454-461.
[4]呂德堅,李建金,劉秋玲,等.ABO血型異常遺傳的親子鑒定案[J].法醫學雜志,2000,16(3):179-180.
[5]Ogasawara K,Yabe R,Uchikawa M,et al.Recombination and gene conversion-like events may contribute to ABO gene diversity causing various phenotypes[J].Immunogenetics,2001,53(3):190-199.
[6]Olsson ML,Michalewska B,Hellberg A,et al.A clue to the basis of allelic enhancement:occurrence of the Ax subgroup in the offspring of blood group O parents[J].Transfus Med,2005,15(5):435-442.
[7]Cho D,Shin MG,Yazer MH,et al.The genetic and phenotypic basis of blood group A subtypes in Koreans[J].Transfus Med, 2005,15(4):329-334.
[8]Kaneko M,Nishihara S,Shinya N.Wide variety of point mutations in the H Gene of Bombay and Para-Bombay individuals that inactivate H enzyme[J].Blood,1997,9(2):839-849.
[9]洪小珍,許先國,朱發明,等.罕見類孟買型的鑒定[J].中國輸血雜志,2005,18(1):8-10.
[10]Drexler C,Glock B,Vadon M,Staudacher E,Dauber EM,Ulrich S et al.Tetragametic chimerism detected in a healthy woman with mixed-field agglutination reactions in ABO blood grouping[J]. Transfusion 2005,45(5):698-703.
[11]Cho D,Lee SK,Yazer MH,et al.A dispermic chimera with mixed field blood group B and mosaic 46,XY/47,XYY karyotype[J].J Korean Med Sci,2007;22(3):553-556.
[12]Yu Q,Li Q,Gao S,et al.Congenital tetragametic blood chimerism explains a case of questionable paternity[J].Forensic Sci,2011,56 (5):1346-1348.
[13]鄧志輝.復合型ABO糖基轉移酶的分子生物學與酶動力學的研究[J]中國輸血雜志2007,20(4):352-355.
[14]Seto NO,Palcic MM,Compston CA,et al.Sequential interchange of four amino acids from blood group B to blood group A glycosyltransferase boosts catalytic activity and progressively modifies substrate recognition in human recombinant enzymes[J].J Bio Chem 1997,272(22):14133-14138.
[15]Yu LC,Lee HL,Chan YS,et al.The molecular basis for the B(A) allele:an amino acid alteration in the human histoblood group B alpha-(1,3)-galactosyltransferase increase its intrinsic alpha-(1,3)-N-acetylgalactosaminyltransferase activity[J].Biochem Biophys Res Commun 1999,262(2):487-493
[16]Chen,DP,Tseng,CP,Wang,WT,et al.A novel cis-AB allele derived from the A transferase gene by nucleotide substitution C796A[J].Ann Clin Lab Sci,2004,34(4):437-442
[17]Deng ZH,Seltsam A,Ye Y,et al.Haemolytic disease of fetus and newborn caused by ABO antibodies in a cisAB offspring[J]. Transfus Apher Sci,2008,39(2):123-128
[18]周炳能.29例ABO血型正反定型不一致原因探討[J].實驗與檢驗醫學,2010,28(2):189-190.
R457.1+2,Q331
A
1674-1129(2013)01-0049-03
10.3969/j.issn.1674-1129.2013.01.017
深圳市科技計劃重點項目(編號:201001021)
喻瓊,女,1971年11月生,中山大學,碩士,主任技師。輸血醫學專業。
鄧志輝,男,1967年10月生,中山大學,博士,主任技師,分子生物學專業,研究方向為免疫遺傳、輸血醫學。
