單增李斯特氏菌磷脂酶C(lm-plcB)基因的克隆表達(dá)及其在油脂脫膠中的應(yīng)用*
2013-08-12 00:57:04余榛榛常明劉睿杰金青哲劉元法王興國
食品與發(fā)酵工業(yè) 2013年12期
余榛榛,常明,劉睿杰,金青哲,劉元法,王興國
(食品科學(xué)與技術(shù)國家重點(diǎn)實(shí)驗(yàn)室 食品安全與營養(yǎng)協(xié)同創(chuàng)新中心 江南大學(xué)食品學(xué)院,江蘇 無錫,214122)
單核細(xì)胞增多性李斯特菌(L. monocytogenes,簡稱Lm)是一種重要的食源性致病菌,兼性厭氧。溫度適應(yīng)性很強(qiáng),-20℃時(shí)仍可部分存活,巴氏消毒也不能完全滅活。菌株對營養(yǎng)要求不高,能在物體表面形成生物膜對抗菌物質(zhì)有抵抗作用,抗逆性較強(qiáng),包括耐酸,耐反復(fù)凍溶,耐干燥以及耐鹽[1]。Lm 在自然界中廣泛存在,易在野生動(dòng)物、家畜、鳥類、昆蟲、污水、土壤和植物中生長,相關(guān)產(chǎn)品如牛、羊、家禽、乳制品、水果、蔬菜、熟肉制品、海產(chǎn)品中也常能檢測到該菌。
研究表明,磷脂酶C(Phospholipase C,PLC)是Lm 的主要毒力因子[1],在細(xì)胞代謝、信息傳遞和抗血小板功能等方面起著重要作用。國內(nèi)外已將其作為重要生理活性成分及醫(yī)藥原料,廣泛應(yīng)用于藥品、化妝品、食品加工等領(lǐng)域[2-4]。PLC 的作用位點(diǎn)為甘油磷脂C3 位酯鍵,水解生成甘油二酯(DAG)及磷脂酸化合物,如磷酸膽堿、磷酸乙醇胺、磷酸絲氨酸以及磷酸肌醇等(圖1)。由于PLC 水解磷脂生成的DAG 是油脂的正常組分,DAG 的變化不僅不會(huì)影響精煉油脂的品質(zhì),而且可以提高精煉油得率,近年來日益得到油脂行業(yè)的關(guān)注,將其應(yīng)用于油脂酶法脫膠。
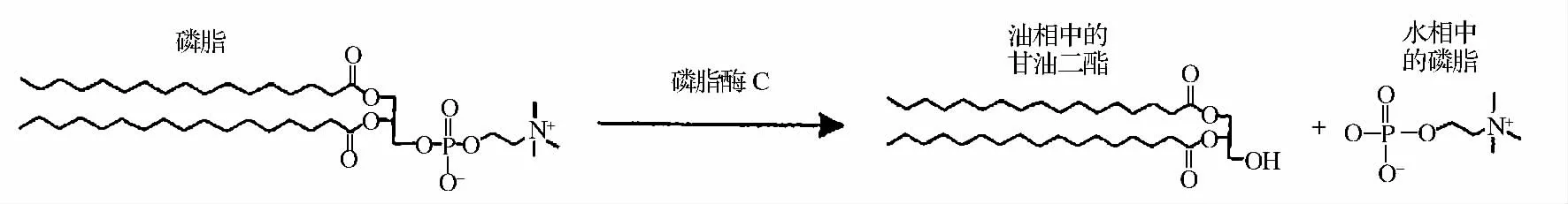
圖1 磷脂酶C 水解磷脂圖Fig.1 Phospholipase C hydrolyze phospholipids
PLC 酶法脫膠具有適用性廣,反應(yīng)條件溫和,生產(chǎn)中節(jié)省酸堿化學(xué)品的消耗,幾乎不產(chǎn)生皂腳和廢水,經(jīng)濟(jì)效益增長明顯等一般酶法脫膠的共同優(yōu)點(diǎn),但相較于其他的酶法脫膠,如磷脂酶A (Phospholipase A ,PLA)酶法脫膠,PLC 酶法脫膠不僅能大大縮短脫膠反應(yīng)時(shí)間,還能有效提高毛油得率,降低毛油損耗,通常每500 mg/L 磷就可以提高約1%的油產(chǎn)量[5],該特點(diǎn)在糧食價(jià)格持續(xù)上漲的現(xiàn)今社會(huì)顯得尤為重要。本文通過克隆單增李斯特氏菌plcB 基因,重組表達(dá)PLC,并應(yīng)用該酶對不同的植物毛油進(jìn)行脫膠研究,具有較大的經(jīng)濟(jì)價(jià)值和現(xiàn)實(shí)意義。
1 材料和方法
1.1 材料
1.1.1 菌株和質(zhì)粒
單核細(xì)胞增多性李斯特菌(L. monocytogenes)CICC No.21540,購自中國工業(yè)微生物菌種保藏管理中心(CICC);大腸桿菌(Escherichia coli)JM109、BL21(DE3)、表達(dá)載體pET-28a(+)均由本實(shí)驗(yàn)室保存;克隆載體pMD18-T simple,購自大連寶生物(TaKaRa)公司。
1.1.2 主要試劑
T4 DNA 連接酶、限制性核酸內(nèi)切酶NotⅠ、EcoRⅠ,均購自NEB 公司;對硝基磷酸膽堿(p-nitrophehylphosphoryl-choline,簡稱p-NPPC)、考馬斯亮藍(lán)R-250,購自Sigma 公司;蛋白分子質(zhì)量標(biāo)準(zhǔn)(低)、Ex Taq DNA 聚合酶、DNA Marker DL10000,均購自大連寶生物(TaKaRa)公司;飽和酚、硫酸卡那霉素(Kana)、氨芐青霉素(Amp)、SanPrep 柱式質(zhì)粒DNA 小量抽提試劑盒、SanPrep 柱式DNA 膠回收試劑盒,San-Prep 柱式PCR 產(chǎn)物純化試劑盒、丙烯酰胺,N,N’-甲叉雙丙烯酰胺(Bis),購自上海生工生物工程公司;大豆毛油,由中糧提供;菜籽毛油,由湖州新市油廠提供;米糠毛油,由鎮(zhèn)江丹陽油廠提供;其他試劑為國產(chǎn)分析純。
1.1.3 培養(yǎng)基
腦心瓊脂培養(yǎng)基:心提取物5 g/L,腦提取物12.5 g/L,蛋白胨10 g/L,葡萄糖2 g/L,NaCl 5 g/L,Na2HPO42.5 g/L,瓊脂15 g/L,pH 7.4。
LB 液體培養(yǎng)基:蛋白胨10 g/L,NaCl 10 g/L,酵母粉5 g/L,pH 7.4。抗性篩選時(shí)加入氨芐青霉素或卡那霉素至終濃度為100 g/mL;LB 固體培養(yǎng)基,添加20 g/L 的瓊脂粉。
TB 液體培養(yǎng)基:蛋白胨12 g/L,酵母粉24 g/L,甘油4 mL/L,磷酸鹽緩沖液(K2HPO412.54 g、KH2PO42.31g)100 mL/L。
1.2 L. monocytogenes 基因組DNA 的提取及l(fā)mplcB 基因的克隆
L. monocytogenes 基因組的提取參照/氯仿-CTAB法[6]。
根據(jù)NCBI 上提供的L. monocytogenes 的plc 基因序列(YP 005964174.1)為模板,去掉信號肽后設(shè)計(jì)上下游引物:上游引物P1:5’-CGGAATTCATGTGTTGTGATGAATACTTACAAA-3'(EcoRI);下游引物P2: 5 ’-ATGCGGCCGCTTATTCATTTGTTTTTTT-3'(NotI)。PCR 反應(yīng)條件為:94℃,5 min;94℃,30 s,50℃,30 s,72℃,1 min,30 個(gè)循環(huán);72℃,10 min,4 ℃保溫。
PCR 產(chǎn)物經(jīng)SanPrep 柱式PCR 產(chǎn)物純化試劑盒純化回收后,與pMD18-T simple 連接,連接產(chǎn)物轉(zhuǎn)化大腸桿菌感受態(tài)細(xì)胞E. coil(JM109),經(jīng)Amp-LB 培養(yǎng)基過夜培養(yǎng)篩選陽性克隆,PCR 鑒定后測序,命名為pMD18-T-lm-plcB。
1.3 重組表達(dá)載體的構(gòu)建
用限制性內(nèi)切酶EcoR I 和Not I 對重組載體pMD18T-lm-plcB 和表達(dá)載體pET28a(+)進(jìn)行雙酶切。酶切產(chǎn)物經(jīng)SanPrep 柱式DNA 膠回收試劑盒純化后利用T4 DNA 連接酶連接,連接產(chǎn)物轉(zhuǎn)化大腸桿菌感受態(tài)細(xì)胞E. coli(JM109),經(jīng)Kana-LB 培養(yǎng)基篩選,并對陽性菌落進(jìn)行PCR 和酶切鑒定。
1.4 重組磷脂酶C 的誘導(dǎo)表達(dá)
分別將表達(dá)質(zhì)粒pET28a-lm-plcB 和pET28a(+)轉(zhuǎn)化原核表達(dá)菌株E. coli BL21(DE3),將獲得的陽性克隆E. coil BL21(DE3)/pET28a-lm-plcB 接種于Kana-LB 液體培養(yǎng)基中,37℃,200 r/min 培養(yǎng)12 h作為種子液。隨后將種子液按5%接種量接種于Kana-TB 培養(yǎng)基中,37℃、200 r/min 振蕩培養(yǎng)進(jìn)行發(fā)酵。當(dāng)培養(yǎng)物OD600達(dá)到0.6 ~0.8 時(shí),加入終濃度為1 g/L 的乳糖于37℃誘導(dǎo)16 h。離心收集菌體,超聲破壁后利用卵黃平板法和SDS-PAGE 鑒定分析重組蛋白[7-8]。
1.5 p-NPPC 法測定磷脂酶C 酶活力
對硝基磷酸膽堿(p-nitrophehylphosphoryl-choline,簡稱p-NPPC)是卵磷脂的一種結(jié)構(gòu)類似物。PLC 可以分解p-NPPC 生成一種顯黃色的對硝基苯酚,對硝基苯酚在410 nm 處有最大吸收,可反映PLC水解p-NPPC 產(chǎn)生對硝基苯酚的量,從而根據(jù)對硝基苯酚標(biāo)準(zhǔn)曲線定量計(jì)算出相應(yīng)磷脂酶C 活力[9-10]。
酶活單位定義為pH 7.2,37℃條件下,每分鐘水解p-NPPC 生成1 nmol 對硝基苯酚所需的酶量為1個(gè)活力單位(U)[11]。酶反應(yīng)體系:50mmol/L Tris-HCl(pH 7.2),10 mmol/L p-NPPC。100μL 的反應(yīng)系統(tǒng)中加入10μL 的酶樣品溶液,37℃反應(yīng)30 min[10]。
1.6 E. coil BL21(DE3)/pET28a-lm-plcB 誘導(dǎo)條件的優(yōu)化
誘導(dǎo)接種量、誘導(dǎo)劑添加時(shí)間、誘導(dǎo)溫度、誘導(dǎo)劑濃度、誘導(dǎo)時(shí)間等因素對產(chǎn)酶的影響隨重組蛋白的種類不同而有所不同。本實(shí)驗(yàn)通過改變單因子的方法對誘導(dǎo)條件進(jìn)行優(yōu)化,以p-NPPC 法獲得的酶活大小作為優(yōu)化參考標(biāo)準(zhǔn)。
1.6.1 誘導(dǎo)接種量的優(yōu)化
在培養(yǎng)基中分別接入1%、2%、3%、4%、5%的菌種,不同接種量分別做3 個(gè)平行。擴(kuò)培4 h 后添加終濃度為1 g /L 的乳糖開始誘導(dǎo)表達(dá),37℃、200 r/min 誘導(dǎo)24h 后檢測發(fā)酵液OD600值,并取等體積的發(fā)酵液超聲破碎后測定酶活力,確定最適接種量。
1.6.2 誘導(dǎo)劑添加時(shí)間的優(yōu)化
分別在重組菌接種后0、2、4、6、8、10、12、14、16、18、20 h 添加終濃度為1 g /L 的乳糖進(jìn)行誘導(dǎo),37℃、200 r/min 誘導(dǎo)24 h 后確定最適誘導(dǎo)時(shí)間。實(shí)驗(yàn)方法同上,每個(gè)時(shí)間梯度設(shè)置3 個(gè)平行實(shí)驗(yàn)。
1. 6. 3 誘導(dǎo)溫度的優(yōu)化
添加乳糖至終濃度1 g /L 后分別在18、25、30、37℃對重組菌株進(jìn)行誘導(dǎo)表達(dá),每個(gè)溫度梯度設(shè)置3個(gè)平行實(shí)驗(yàn)。實(shí)驗(yàn)方法同上,根據(jù)實(shí)驗(yàn)結(jié)果確定最適誘導(dǎo)溫度。
1. 6. 4 誘導(dǎo)劑濃度的優(yōu)化
在上述已優(yōu)化的誘導(dǎo)基礎(chǔ)上,添加乳糖至終濃度為1、2.5、5、8、10、15、20 g /L 進(jìn)行梯度誘導(dǎo),不同的乳糖濃度設(shè)置3 個(gè)平行實(shí)驗(yàn)。實(shí)驗(yàn)方法同上,根據(jù)結(jié)果確定最適誘導(dǎo)劑濃度。
1. 6. 5 誘導(dǎo)時(shí)間的優(yōu)化
在上述已優(yōu)化的誘導(dǎo)條件下進(jìn)行誘導(dǎo),分別誘導(dǎo)4、6、8、10、12、14、16、18、20、22、24、26、28、30 h 后,檢測發(fā)酵液OD600值,并取等體積的發(fā)酵液,超聲破碎細(xì)胞后測定酶活力大小,確定最適誘導(dǎo)時(shí)間。不同的誘導(dǎo)時(shí)間設(shè)置3 個(gè)平行實(shí)驗(yàn)。
1.7 磷脂酶C 脫膠效果
將植物毛油(大豆毛油、菜籽毛油、米糠毛油)于具塞三角燒瓶中水浴加熱到80 ℃,加入總毛油質(zhì)量0.5%,濃度45%(w/w)的檸檬酸溶液,在500 r/min攪拌20 min,進(jìn)行酸預(yù)處理。將經(jīng)過酸預(yù)處理的油樣冷卻至47℃,加入4% NaOH 溶液調(diào)節(jié)pH 至5 。再加入油重1%的蒸餾水和油重1%的重組磷脂酶C,均勻混合后,在62℃下300 r/min 攪拌1 h,進(jìn)行酶反應(yīng)。反應(yīng)結(jié)束后將油樣置于90℃水中加熱10 min 滅酶,再測定磷含量[12]。
油相pH 的測定:用50 mL 離心管取油水混合物40 g,在5 000 r/min 離心10 min,棄去上層油相,向沉淀中加入5 mL 的蒸餾水,充分?jǐn)嚢杌旌虾笤俅卧? 000 r/min 離心10 min,取離心管中水相用pH 計(jì)測定pH 值,經(jīng)校正后采用。油相pH 為5.0 左右時(shí)的經(jīng)驗(yàn)校正公式:pH 實(shí)際=pH 測定-0.3[13]。
磷含量的測定:將脫膠后的油樣于10 000 r/min下離心10 min,取5 g 上層油樣進(jìn)行磷含量測定。油脂中磷含量測定參照GB/T5537 -2008。
2 結(jié)果與分析
2.1 L. monocytogenes 基因組DNA 的提取及l(fā)mplcB 基因的制備
以L. monocytogenes 全基因序列為模板,以引物P1 和P2 進(jìn)行PCR 擴(kuò)增,產(chǎn)物進(jìn)行1%瓊脂糖凝膠電泳,擴(kuò)增出的DNA 片段在約810bp 處,與預(yù)期大小相符(圖2)。測序結(jié)果顯示目的片段與L. monocytogenes 的plc 基因序列(YP 005964174.1)相似度達(dá)到99%,蛋白質(zhì)序列相似度為100%。
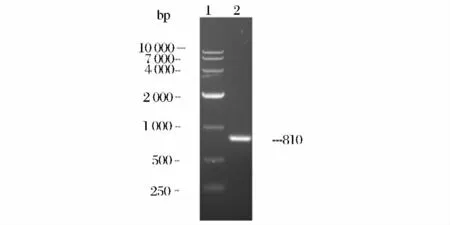
圖2 lm-plcB 的PCR 鑒定Fig.2 PCR result of lm-plcB
2.2 重組表達(dá)載體pET28a-lm-plcB 的鑒定
對構(gòu)建的表達(dá)質(zhì)粒pET28a-lm-plcB 進(jìn)行菌落PCR 驗(yàn)證,在810bp 處有目的條帶;進(jìn)一步對pET28a-lm-plcB 進(jìn)行EcoRⅠ和NotⅠ雙酶切鑒定,分別獲得約5 300 bp 和約810 bp 的條帶(圖3),與預(yù)期結(jié)果相符。
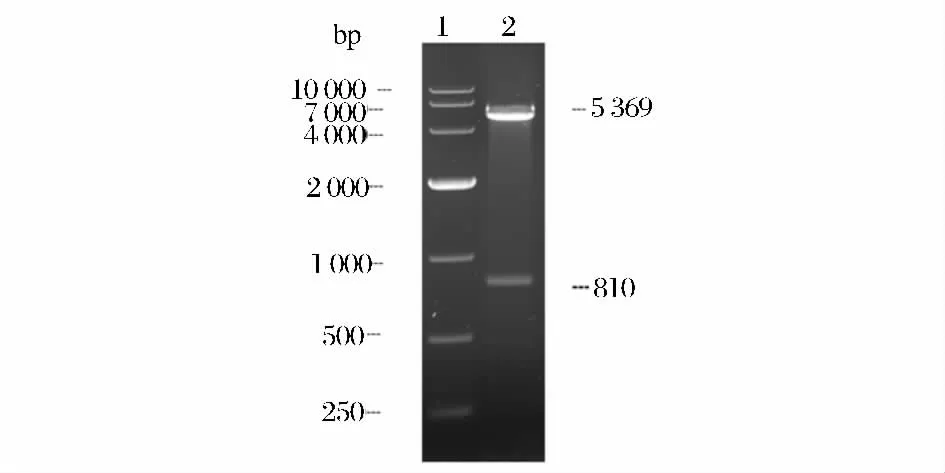
圖3 pET28a-lm-plcB 的酶切鑒定Fig.3 Restriction enzyme digestion analysis pET28a-lm-plcB by EcoRⅠand NotⅠ
2.3 E. coil BL21(DE3)/pET28a-lm-plcB 誘導(dǎo)表達(dá)及產(chǎn)物活性鑒定
E. coil BL21(DE3)/pET28a-lm-plcB 經(jīng)誘導(dǎo)表達(dá)后超聲破碎,12 000 r/min 分離上清液。利用卵黃硼砂平板分析重組PLC 的活性,結(jié)果顯示:E. coil BL21(DE3)/pET28a 在卵黃硼砂平板上沒有出現(xiàn)水解圈,重組PLC 產(chǎn)生乳白色水解圈,且添加10 mol/L Mg2+后水解圈明顯增大,說明該重組蛋白對天然磷脂底物具有PLC 活性,Mg2+對其分解天然底物的活性具有促進(jìn)作用(圖4)。
SDS-PAGE 電泳結(jié)果表明 E. coil (BL21)/pET28a-lm-plcB 表達(dá)了相對分子質(zhì)量約為30.4 kDa的重組蛋白,與預(yù)期大小相符(圖5)。
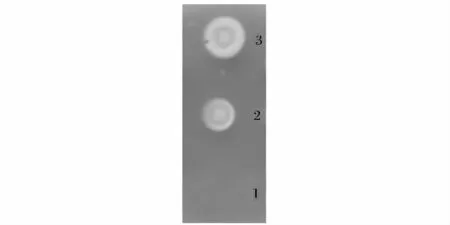
圖4 重組蛋白的活性檢測Fig.4 Analysis of recombinant PLC activity
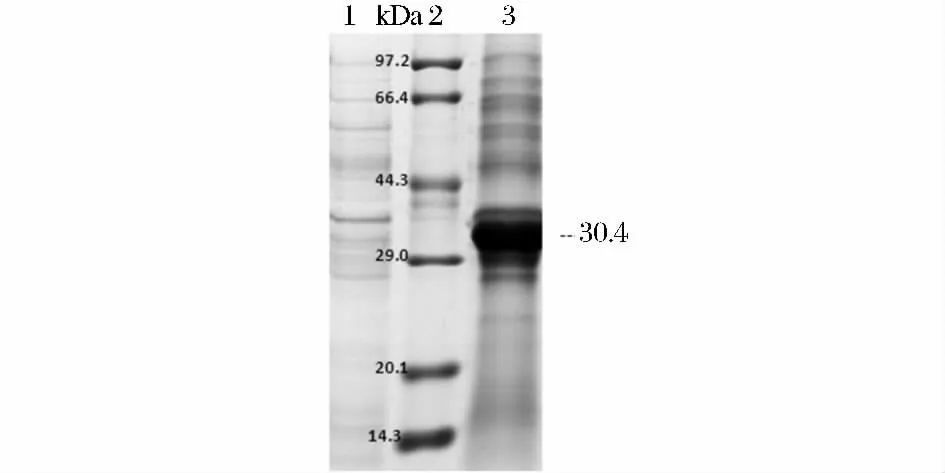
圖5 重組PLC 的SDS-PAGEFig.5 SDS-PAGE analysis of recombinant PLC in E.coil BL21(DE3)/pET28a-lm-plcB
2.4 E. coil BL21(DE3)/pET28a-lm-plcB 誘導(dǎo)條件的優(yōu)化
2.4.1 誘導(dǎo)接種量的優(yōu)化
由圖6 可知,在接種量為4%時(shí),重組菌產(chǎn)酶量達(dá)到最大,約159.1 U/mL,故最適接種量為4%。
2.4.2 誘導(dǎo)劑添加時(shí)間的優(yōu)化
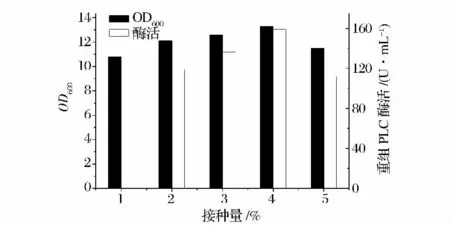
圖6 不同接種量對產(chǎn)酶的影響Fig.6 Effect of inoculation on cell density and PLC activity
由圖7 可知,在接種后2h 添加乳糖,重組菌產(chǎn)酶量達(dá)到最大值,約411.9 U/mL 左右。故乳糖添加時(shí)間為轉(zhuǎn)接后2 h。
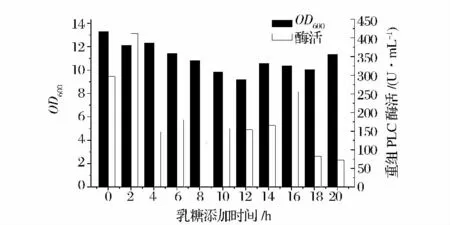
圖7 乳糖添加時(shí)間對酶活力的影響Fig.7 Effect of lactose addion time on cell density and PLC activity
2.4.3 誘導(dǎo)溫度的優(yōu)化
如圖8,誘導(dǎo)溫度為25℃時(shí),重組蛋白的酶活力達(dá)到最大,可達(dá)到431.5 U /mL 左右。故選取發(fā)酵產(chǎn)酶的誘導(dǎo)溫度為25℃。
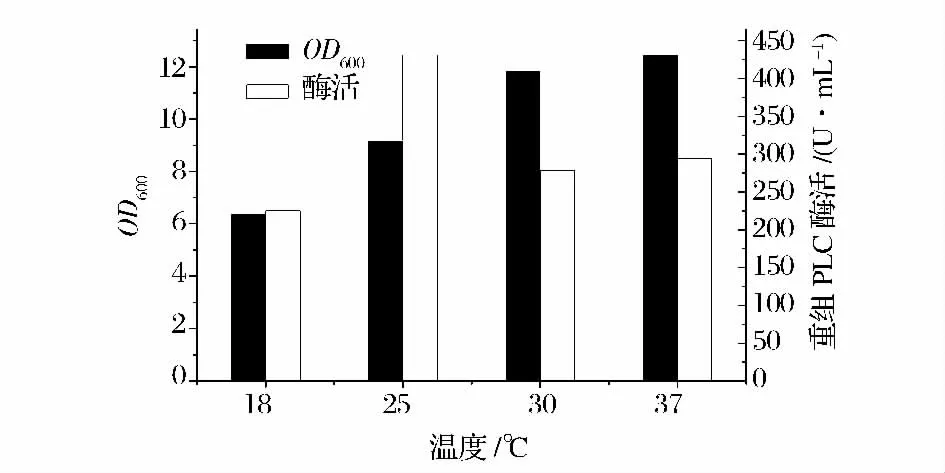
圖8 添加乳糖時(shí)誘導(dǎo)溫度對產(chǎn)酶的影響Fig.8 Effect of lactose concentration on cell density and PLC activity
2.4.4 誘導(dǎo)劑濃度的優(yōu)化
由圖9 可見,在乳糖濃度為2.5 g /L 時(shí),重組菌產(chǎn)酶量達(dá)到最大,約633.1 U /mL 左右,故乳糖誘導(dǎo)劑的最適誘導(dǎo)濃度為2.5 g/L。
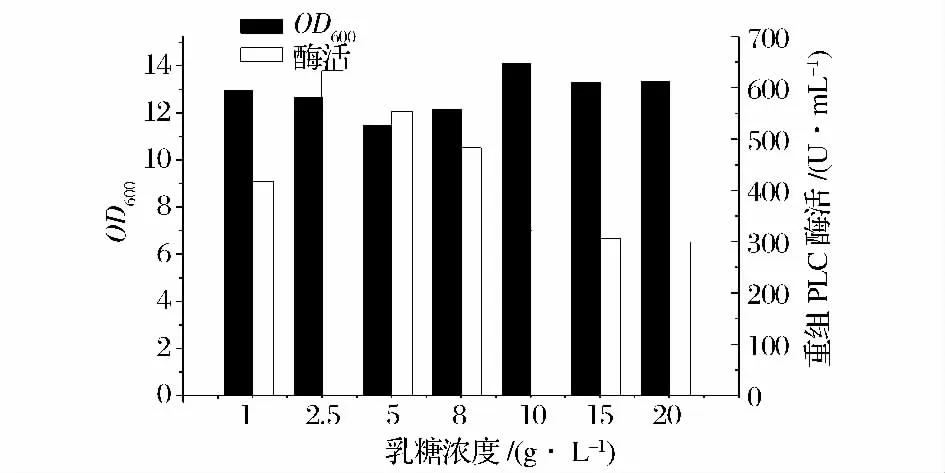
圖9 乳糖質(zhì)量濃度對酶活力的影響Fig.9 Effect of lactose concentration on cell density and PLC activity
2.4.5 誘導(dǎo)時(shí)間的優(yōu)化
由圖10 可知,乳糖添加后26 h,重組蛋白的酶活力達(dá)到最大值,約為754.6 U /mL,故最適誘導(dǎo)時(shí)間為26 h。
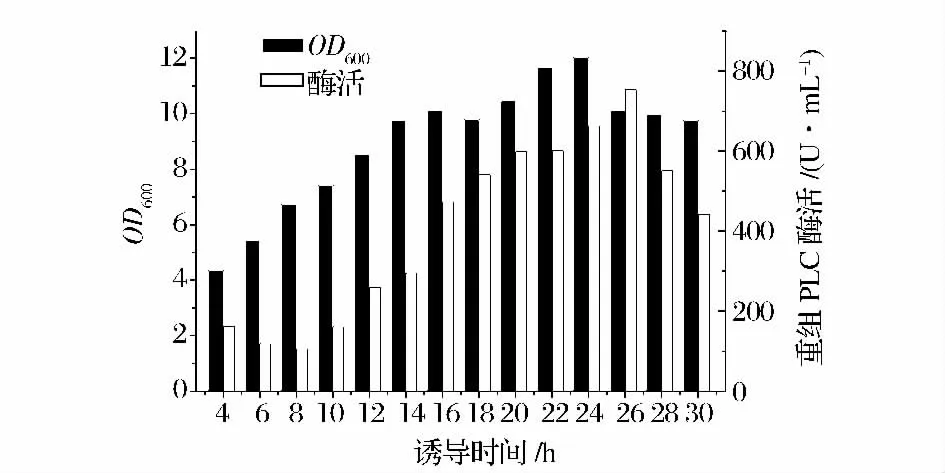
圖10 添加乳糖時(shí)誘導(dǎo)時(shí)間對產(chǎn)酶的影響Fig.10 Effect of lactose induction time on phospholipase activity
2.5 重組磷脂酶C 脫膠效果的初步研究
利用該重組PLC 對不同植物毛油進(jìn)行脫膠,最終將大豆毛油中的含磷量從216.67 mg/kg 下降到3.705 mg/kg,磷含量降低了98.40%;菜籽毛油中的含磷量從183.7 mg/kg 下降到4.503 mg/kg,磷含量比脫膠前降低了97.55%;米糠毛油中的含磷量從306.8 mg/kg 下降到50.191 mg/kg,磷含量比脫膠前降低了83.63%。
3 討論
本文對單增李斯特氏菌來源的lm-plcB 基因在大腸桿菌中進(jìn)行了重組表達(dá),并首次應(yīng)用單增李斯特氏菌來源的重組PLC 對不同的植物毛油進(jìn)行脫膠,最終使大豆毛油和菜籽毛油的磷含量降低到了5 mg/kg以下,避免了傳統(tǒng)油脂精煉環(huán)節(jié)的脫酸處理,可以實(shí)現(xiàn)油脂的物理精煉。利用PLC 脫膠,油脂中的水化磷脂可以被水解生成DAG 和水溶性磷酸酯,DAG 作為油脂的固有組分可以伴隨油相被回收,避免了中性油的損失,大大降低了毛油煉耗率,較溫和的處理?xiàng)l件也顯著提高了油脂的品質(zhì)。因此,單增李斯特氏菌來源的重組磷脂酶C 在油脂工業(yè)中的應(yīng)用具有較大的經(jīng)濟(jì)意義和社會(huì)效益。
為了進(jìn)一步開發(fā)符合工業(yè)化生產(chǎn)的磷脂酶C 制劑,還需對該重組磷脂酶C 的酶學(xué)性質(zhì)進(jìn)一步深入研究,探索其底物特異性,針對不同毛油產(chǎn)品,開發(fā)混合型酶制劑,增強(qiáng)產(chǎn)品的適應(yīng)性,提升制劑的穩(wěn)定性。相信隨著PLC 結(jié)構(gòu)功能研究的不斷深入和油脂酶法脫膠技術(shù)的不斷完善,PLC 酶法脫膠將得到進(jìn)一步推廣,油脂精煉也將更加符合健康、環(huán)保和低耗的理念。
[1] 邸薇. 單增李斯特菌iap、actA、plcB 基因的原核表達(dá)、產(chǎn)物的免疫原性分析以及重組p60 蛋白多抗的制備[D]. 廣州:暨南大學(xué),2012:8 -11.
[2] 陳明錯(cuò). 磷脂酶C 抗血小板功能極其機(jī)制的研究[D].武漢:武漢大學(xué),2002.
[3] De Maria L,Vind J,Oxenboll K M,et al. Phospholipases and their industrial applications[J]. Applied Microbiology and Biotechnology,2007,74(2):290 -300.
[4] 余榛榛,常明,劉睿杰,等. 磷脂酶C 在酶法脫膠中的研究進(jìn)展[J]. 中國油脂,2013,38(7):19 -22.
[5] Barton N R.A new process for degumming:the use of phospholipase C to improve yields during refining of high phosphorus vegetable oils[C]. Seattle:The 99th AOCS Annual Meeting & Expo,2008:120.
[6] 徐義剛,李蘇龍,謝曉峰,等. 食源性致病菌基因組DNA 的高效提取方法[J]. 東北林業(yè)大學(xué)學(xué)報(bào),2009,37(2):73 -78.
[7] Chrisope G,F(xiàn)ox C,Marshall R T.Lecithin agar for dectection of microbial phospholipase[J].Appl ied and Environmental Microbiology.1976,31(5):784 -786.
[8] Gubash S.Improved egg-yolk agar plate medium for the detection of Clostridial phospholipase C activity[J].Research in Microbiology,1991,142(1):87 -93.
[9] 彭愈生,周定. 磷脂酶C 活性測定方法及其應(yīng)用的研究[J]. 哈爾濱工業(yè)大學(xué)學(xué)報(bào),1989,6(3):34 -38.
[10] Kurioka S,Matsuda M.Phospholipase C assay using p-nitrophehylphosphoryl-choline together with sorbitol and its application to studying the metal and detergent requirement of the enzyme[J].Analytical Bichemistry,1976,75(1):281 -289.
[11] Berka R M,Vasil M L.Phospholipase C (heat-labile)hemolysin of Pseudomonas aeruglnosa:purification and preliminary characterization[J].Journal of Bacteriology,1982,152(1):239 -245.
[12] 楊嬌,金青哲,王興國. 磷脂酶C 用于大豆油脫膠的工藝優(yōu)化[J]. 中國油脂,2012,37(12):14 -17.
[13] Clausen K.Enzymatic oil-degumming by a novel microbial phospholipase[J]European Journal of Lipid Science and Technology,2001,103(6):333 -340.
猜你喜歡
食品工業(yè)(2023年12期)2023-12-12 12:56:16
房地產(chǎn)導(dǎo)刊(2022年5期)2022-06-01 06:20:14
建材發(fā)展導(dǎo)向(2021年12期)2021-07-22 08:06:48
建材發(fā)展導(dǎo)向(2021年7期)2021-07-16 07:07:52
中學(xué)生數(shù)理化(高中版.高二數(shù)學(xué))(2021年12期)2021-04-26 07:43:48
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2021年12期)2021-03-08 01:28:50
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
中國糧油學(xué)報(bào)(2019年4期)2019-07-12 09:06:38
食品與機(jī)械(2018年11期)2019-01-03 06:15:34
中國糧油學(xué)報(bào)(2018年12期)2018-01-26 13:32:04
