海南島海草床群落種間關系研究
2013-08-14 05:49:16陳石泉吳鐘解王道儒張光星蔡澤富
海洋通報 2013年1期
陳石泉,吳鐘解,王道儒,張光星,蔡澤富
(海南省海洋開發規劃設計研究院,海南 海口 570125)
海草(seagrass)是生活于熱帶和溫帶海域淺水的單子葉植物(楊宗岱,1979),一般分布在低潮帶和潮下帶;大多數海草種分布在20 m以淺海域內(Dennison et al,1993);6 m以淺范圍區域一般是海草分布的主要區域,最深可分布在水下90 m(Den,1970)處。根據楊宗岱、林鵬、范航清等人海草調查工作統計,海南海草種類為2科、6亞科、8屬、13種(中國科學院中國植物志編輯委員會,1992;范航清等,2009)。海草可以阻止和吸附水流中的懸浮顆粒,能夠消除污染、凈化水質,改善水質環境,是控制近岸淺水水質的關鍵植物;能減弱海浪能、水流能,對維護海岸,保持海床穩定方面起著很大作用,為許多海洋生物提供食物來源,如:儒艮、綠海龜、海膽、海馬、蟹類、沙蟲、海葵、海鞘等等,近年來,由于養殖活動、陸源污染、航道、港口建設以及填海造地等海洋工程開發,導致海草群落和海草底棲生物區域性滅絕,海草棲息地不斷減少。
物種的聯結性(association between species)指不同種類在空間分布上的相互關聯性,通常以物種的存在與否為依據,是一種定性的數據,種間關聯(interspecific association)指種間相互吸引或排斥的性質,這種關聯可以是正的,表示相互“喜歡”,也可以是負的,表示相互排斥,種間關聯不局限于物種存在與否的二元數據,還涉及他們的數量多少,是一種定量的關系(張金屯,2004)。種間聯結測定值可以在一定程度上衡量物種之間的相互關系以及對環境生態因子反應的差異,對不同物種個體空間聯結性的測定分析,可以為研究群落動態演替、群落格局形成、種群進化等提供理論依據(黃世能等,2000)。物種的聯結性與種間相關性是植物群落重要數據與結構特征之一(張震云等,2011)。近期,許多科研工作者對不同物種種間的聯結性進行了研究(郭志華等,1997;歐祖蘭等,2002;李先琨等,1999;蔡冰玲等,2010),也有部分科研工作者對荒漠植物種間關聯性進行了研究(尹林克,2005),海草生態學領域研究主要涉及海草分布、食物鏈、能量流動和物質循環等方面(楊宗岱等,1981;黃道建等,2007;鄭杏雯,2007;韓秋影等,2008;張景平等,2008),海南島海草床群落種間相關性研究尚未見報道。
本文采用 X2檢驗、Ochiai(1957)指數和Spearman秩相關系數對海南海草群落的種間關系進行研究,以揭示不同海草種群之間的生態關系,進而探究群落的結構特征以及不同物種對環境的適應特點,為海草床生態系統的保護及恢復提供理論依據。
1 材料與方法
1.1 研究區域與樣地概況
研究區域為海南省文昌的高隆灣、港東村、長圮港、寶峙村、馮家灣沿岸海域,瓊海的青葛、龍灣、潭門沿岸海域,陵水的新村港、黎安港,三亞的后海灣、鐵爐港,澄邁的花場灣等13個區域,20°9′57.43″N-18°9′33.21 ″N,111°16′43.22″E-108°36′37.58″E,研究區域花場灣、鐵爐港、黎安港和新村港海草床主要分布在瀉湖沿岸,其中花場灣海草床沉積物組成類型是淤泥,鐵爐港、黎安港和新村港海草床沉積物組成類型是泥砂;港東村、寶峙村、青葛、后海灣、馮家灣、潭門、長圮、龍灣和高隆灣海草床主要分布在珊瑚礁港灣海岸,海草床沉積物組成類型是珊瑚屑、貝殼屑、礫石、粗砂、中砂和細砂,長圮港有紅樹林分布,有少量淤泥。研究區域海水鹽度在28.57~33.32之間,水溫在25.86~30.3℃之間,水質透明度在1~6m之間,pH變化范圍在7.9~8.25。

圖1 海南島海草資源調查區域
1.2 樣品采集與處理
采用樣方法進行野外調查,于2009年對海南省文昌的高隆灣、港東村、長圮港、寶峙村、馮家灣沿岸海域,瓊海的青葛、龍灣、潭門沿岸海域,陵水的新村港、黎安港,三亞的后海灣、鐵爐港,澄邁的花場灣13個區域,每個區域設定2個垂直岸線的斷面,斷面分布在潮間帶和潮下帶海草分布范圍,每個區域采用樣方法調查,每個斷面上垂直放置10~20個0.25 m×0.25 m的樣方,共計192個樣方,樣方放置盡量均勻且能覆蓋在不同的海草種類上。采齊樣方內的海草,并對海草的種類、蓋度、密度、生物量以及底質等指標進行分析。野外調查共調查到10個海草種(表1),采集的海草及生境(表2),將調查到的海草種組成10×192的原始數據矩陣并進行種間關聯性和相關性分析。

表1 10種海草優勢種的種名及種號
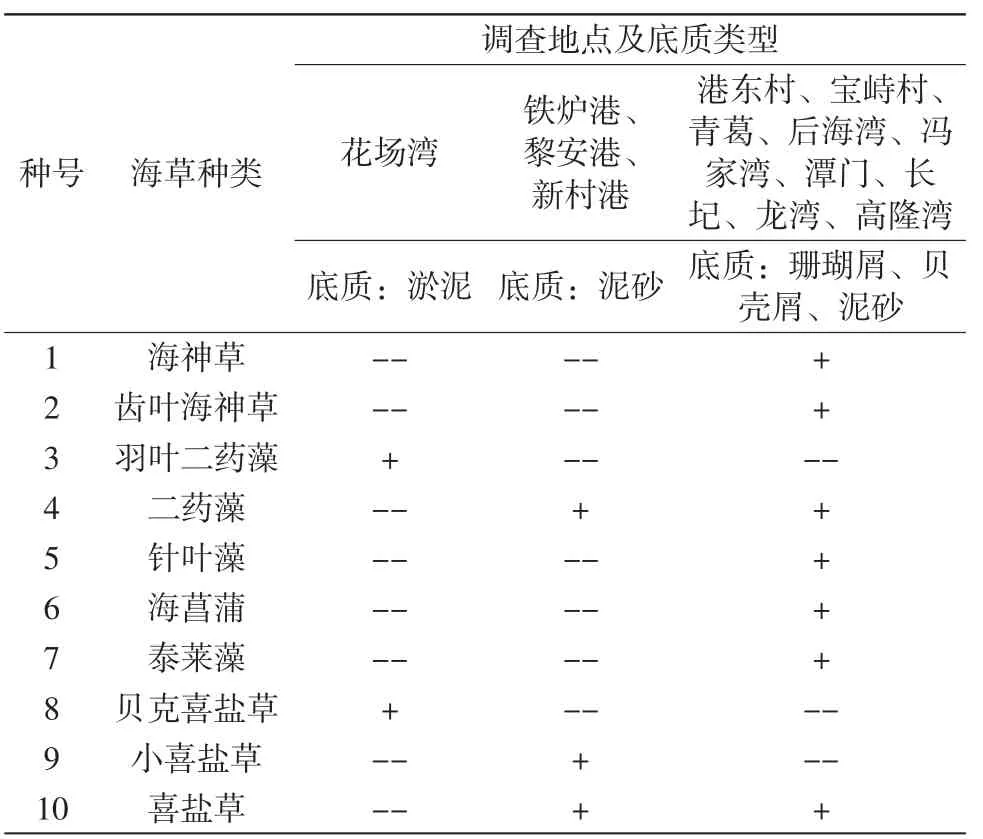
表2 10種海草與底質類型
1.3 數據分析
1.3.1 關聯性分析
根據物種在各樣方內存在與否將10×192的數據矩陣轉化為0、1形式的二元數據矩陣,用于X2檢驗。X2檢驗公式為:
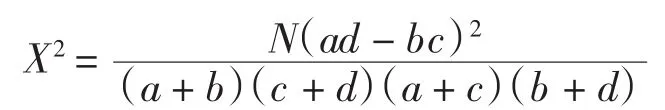
式中:N為樣方總數,a為2物種均出現的樣方數,b、c為僅有一個物種出現的樣方數,d為2物種均未出現的樣方數。當a d>b c時為正關聯,a d<b c時為負關聯。在下列2種情況下X2值被認為是有偏差:1)2×2列聯表中任一小格期望值<1;2)多于2個小格的期望值<5。這種偏差用Yates連續校正系數來糾正。Yates系數糾正公式為:
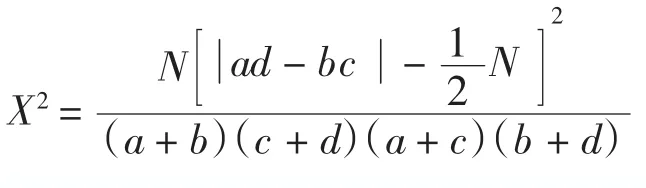
若 X2>3.841(0.01<P<0.05)表示種間聯結性顯著。若X2>6.635(P<0.01)表示種對間聯結性極顯著。本研究應用數據為Yates系數糾正后的X2。應注意的是,X2的計算結果與樣方大小關系較密切,應選擇適當大小的樣方。
1.3.2 種間關聯程度的分析
選用Ochiai(1957)指數來描述種間關聯程度,仍以0、1為基礎,a、b值含義同上。指數越接近1,表示種間的正關聯越顯著。公式為:

式中:I代表關聯指數。
1.3.3 Spearman秩相關分析
采用10個優勢種的多度值作為數量指標,進行Spearman秩相關分析。
Spearman秩相關系數:
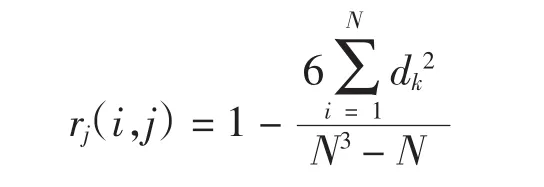
式中:N為樣方數,dk=(Xik-Xi)(Xjk-Xj),Xik和Xjk分別為種i和種j的在樣方k中的相對多度值,Xi和Xj是種i和種j的在所有樣方中多度值的平均值。
2 結果與分析
2.1 種間關聯
X2檢驗能比較準確的檢測出種對的關聯顯著程度,在10個海草種類組成的45對種對中,有16對種對表現出正關聯(a d>b c)(圖2),占種對總數的35.6%,其中有10對表現出極顯著和顯著關聯性(X2>3.841),占種對總數的22.2%;在16對正關聯的種對中,極顯著關聯(X2>6.635)的有8對,占正關聯對數總數50.0%,它所占比例較高,它們是種1-種2(海神草-齒葉海神草),種1-種4(海神草-二藥藻),種1-種5(海神草-針葉藻),種2-種5(齒葉海神草-針葉藻),種1-種7(海神草-泰萊藻),種2-種7(齒葉海神草-泰萊藻),種5-種7(針葉藻-泰萊藻),種4-種9(齒葉海神草-小喜鹽草),通過樣地分析發現(見表2),種5(針葉藻)主要分布在寶峙村與潭門,種2(齒葉海神草)主要分布在寶峙村與馮家灣,種4(二藥藻)分布在高隆灣與長圮港,種1(海神草)與種7(泰萊藻)除鐵爐港與花場灣外都有分布,這些地區分布在珊瑚礁港灣海岸,海草床沉積物組成類型是珊瑚屑、貝殼屑、礫石、粗砂、中砂和細砂;種1(海神草)、種4(二藥藻)、種7(泰萊藻)在潮間帶的低潮帶、潮下帶上部均有分布,種2(齒葉海神草)和種5(針葉藻)則主要分布在潮間帶的低潮區至潮下帶上部水深2m的范圍內;這幾種海草株高差不多,如:種1(海神草)平均株冠高度為12.97 cm、種2(齒葉海神草)為10.57 cm、種4(二藥藻)為6.82 cm、種5(針葉藻)為11.67cm、種7(泰萊藻)為13.55cm,這樣相互間就能合理充分利用光能。可見,這些種對對綜合環境的條件相似,在一定程度上體現了生態位的重疊。

圖2 2×2列聯表X2檢驗半矩陣圖
有29對種對表現出負關聯(a d<b c),占種對總數的64.4%,其中有10對表現出極顯著及顯著負關聯(X2>3.841),占負關聯對數總數的34.5%,它們是種1-種6(海神草-海菖蒲),種2-種6(齒葉海神草-海菖蒲),種3-種6(羽葉二藥藻-海菖蒲),種4-種6(二藥藻-海菖蒲),種5-種6(針葉藻-海菖蒲),種6-種7(海菖蒲-泰萊藻),種7-種8(泰萊藻-貝克喜鹽草),種1-種10(海神草-喜鹽草),種6-種10(海菖蒲-喜鹽草),種7-種10(泰萊藻-喜鹽草),通過樣地分析發現(表2),種6(羽葉二藥藻)與種8(貝克喜鹽草)只分布在花場灣,花場灣的底質類型為淤泥,其他種海草主要分布在在珊瑚屑、貝殼屑、礫石、粗砂、中砂、細砂等環境中,太多淤泥環境中生長不好;種6(海菖蒲)與種1(海神草)、種7(泰萊藻)分布潮間帶的低潮帶、潮下帶上部,種4(二藥藻)和種10(喜鹽草)則主要在潮間帶的低潮區分布,種9(小喜鹽草)、種6(羽葉二藥藻)和種8(貝克喜鹽草)分布范圍比較狹窄,主要分布在潮間帶的低潮區,種5(針葉藻)和種2(齒葉海神草)則主要分布在潮間帶的低潮區至潮下帶上部水深2 m的范圍內;種6(海菖蒲)平均株冠高度為44.11 cm、種6(羽葉二藥藻)為10.86 cm、種8(貝克喜鹽草)為1.70 cm、種9(小喜鹽草)2.35 cm、種10(喜鹽草)3.37 cm,它們株高存在巨大的差異,導致光合作用的巨大差異,存在很強對空間環境的相互競爭關系。可見,由于它們對資源空間環境的要求不同或種間存在一定的其他作用原因,使得種間表現出負聯結。
群落系統發育成熟度越高,群落結構及其種類組成越趨于穩定,種間關系也趨于正關聯,從而使得得物種間的穩定共存(朱利君等,2005)。總體上,海草種負關聯種對稍微多于正關聯的種對,顯著和極顯著關聯的種對較少,說明其群落處于較不穩定狀態,處在不穩定發展演替之中,容易受外界因素干擾而產生波動,如:2005年調查發現黎安港有針葉藻分布,因圍海造塘引起生態環境發生變化,導致針葉藻局域滅絕。
2.2 關聯度指數分析
X2檢驗僅得出了物種間聯結性是否顯著,對那些檢驗不是很顯著的種對,并不意味著它們之間不存在聯結性,X2檢驗不能區分聯結程度的大小,因此,X2檢驗只有與關聯度指數結合使用才能較好地反映種間關系(李軍玲等,2004)。關聯度指數是一種描述種間聯結程度的系數,數值越大說明兩物種間關聯程度越大,數值越低說明兩物種關聯性越小,根據10個海草種的Ochiai(1957)指數繪制半矩陣圖(圖3)顯示,I=0的有23對,占總對數的51.1%,全部為負關聯種對,表明這些種對由于對底質環境要求的差異而不可能同時出現;I>0的有22對,占總對數的48.9%,I>0.3的有種1-種2(海神草-齒葉海神草),種1-種5(海神草-針葉藻),種2-種5(齒葉海神草-針葉藻),種1-種7(海神草-泰萊藻),這幾對都成極正聯結,且聯結系數比較高,最高為2-種5(齒葉海神草-針葉藻)聯結系數為:0.500,其它正聯結的聯結系數在0.1<I<0.3,且顯著聯結的種對聯結值明顯高于其它正相關種對。而部分負聯結的種對聯結系數也有I>0,但一般在0.1以下。總體來看I<0.1的種對占多數。可知,負關聯種對多于正關聯,這與X2檢驗結果一致,表明海南省海草床種間聯結性還是比較不穩定。

圖3 Ochiai(1957)相似性系數半矩陣圖
2.3 種間相關
秩相關系數是反映2個物種線形關系的重要指標,其處理的是定量可靠數據,反映2個物種同時出現的可能性程度。海南省海草床Spearman秩相關分析結果顯示呈極顯著和顯著相關的種對有13對,占總種對數的28.9%,其中6對呈正相關,9對呈負相關,包含所有X2檢驗中呈現出的顯著與極顯著關聯的種對,不顯著相關的種對有32對,占總種對數的71.1%,且負相關種對數多于正相關種對數(圖4)。種間秩相關系數高說明兩物種間存在互利關系,或者這2個物種對環境條件的需求相似,相反,系數低或負值說明這2個物種對需求的環境條件不同或者相互間具有一定的排斥性。總體上看,呈極顯著負相關的種對比X2檢驗多2對,它們是種1-種3(海神草-羽葉二藥藻),種3-種7(羽葉二藥藻-泰萊藻),通過樣地分析發現(見表2),種3(羽葉二藥藻)只分布在花場灣。花場灣的底質類型為淤泥,種1(海神草)與種7(泰萊藻)分布在珊瑚屑、貝殼屑、礫石、粗砂、中砂、細砂等環境中,淤泥質環境中未發現分布;呈極正相關的種對只有4對,為種1-種2(海神草-齒葉海神草),種1-種4(海神草-二藥藻),種1-種5(海神草-針葉藻),種2-種5(齒葉海神草-針葉藻),比X2檢驗少了4對,種4(二藥藻)分布在馮家港和寶峙村,底質主要珊瑚屑、貝殼屑及沙礫,種5(針葉藻)分布環境與種4(二藥藻)類似,在馮家港和潭門,種1(海神草)與種2(齒葉海神)分布較廣,分布區主要類型除了珊瑚屑、貝殼屑、礫石、粗砂、中砂、細砂等外,少量淤泥中也能生長。可見,X2檢驗與秩相關分析的結果非常一致,但秩相關分析的靈敏度要遠遠高于X2檢驗。
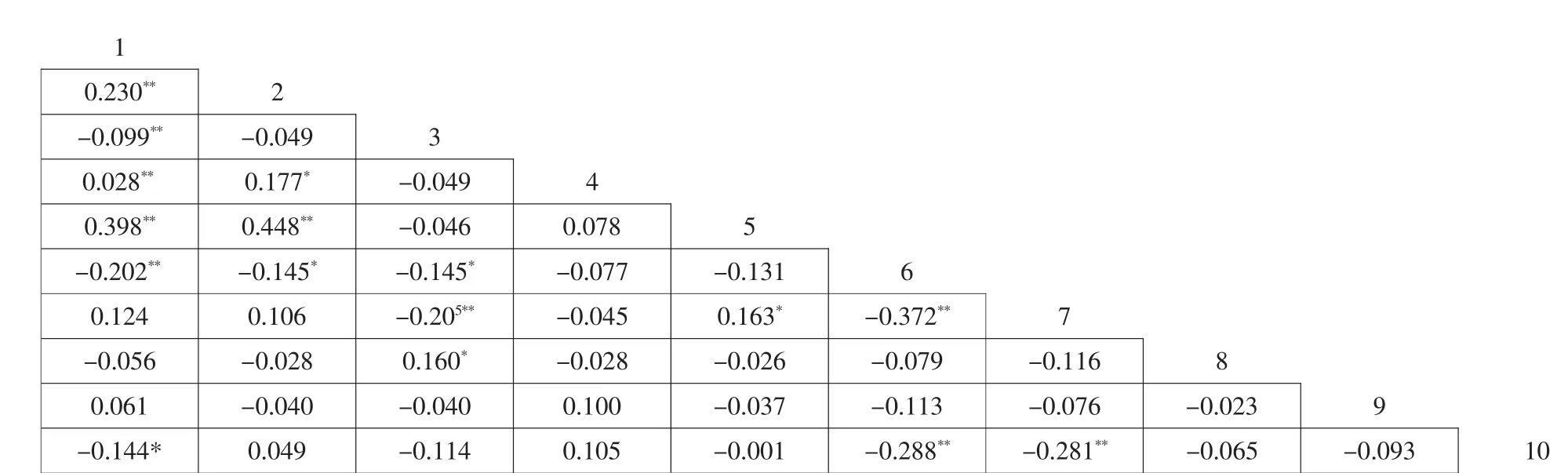
圖4 Spearman相關系數半矩陣圖
2.4 生態種組劃分
海草由于分布環境相同或相似,造成各種海草分布有一定聯系;但由于海草種類和分布環境有一定區別,又造成各種海草分布有一定差異(閆美芳等,2006)。海南島各種海草分布關系有單種海草單獨分布特性,也有兩種或兩種以上共同生長特性。植物之間的正聯結體現了植物之間對資源利用的相似性和生態位的重疊性,植物之間的負聯結體現了植物之間的相互排斥,這是不同生活環境和空間資源對植物選擇的結果,也是生態位分離的原因。依據 X2檢驗、Ochiai(1957)指數分析和Spearman秩相關分析,以負聯結作為劃分種組的分界限,同一種組內兩個種之間盡可能大的正聯結性為原則,并結合10個海草種對綜合生態因子的適應程度和種間聯結差別,將其劃分為2個生態種組。
第1生態種組為種3(羽葉二藥藻)和種8(貝克喜鹽草),這兩種海草只分布于花場灣,花場灣海草分布區主要是瀉湖,沿岸有紅樹林分布,底質類型為淤泥。可知,羽葉二藥和貝克喜鹽草為喜生淤泥物種。
第2生態種組為種1(海神草)、種2(齒葉海神)、種4(二藥藻)、種5(針葉藻)、種7(泰萊藻)、種9(小喜鹽草)及種10(喜鹽草),這幾種海草分布較廣,主要在底質為碎屑、砂質環境中,種9(小喜鹽草)及種10(喜鹽草)在砂質與少量淤泥中也有分布。
3 討論
臺灣柯智仁等通過對臺灣海草分類與分布的研究中認為熱帶、亞熱帶地區的海草和珊瑚在分布上多有關系,如:喜鹽草、泰萊藻、海神草、齒葉海神草、針葉二藥藻及二藥藻等均能在珊瑚沙、珊瑚碎屑及珊瑚礁平臺上生長,喜鹽草羽葉二藥藻及二藥藻能在泥質灘地上生長,臺南及嘉義縣軟泥質的鹽田中有貝克喜鹽草及喜鹽草的分布(柯智,2004),這與本次實地調查一致(表2)。
本文主要采用X2檢驗和 Spearman秩相關分析,從種間關聯和種間相關2個方面探討了海南島13個區域海草生態床群落種間關系。Spearman秩相關分析得出的呈顯著的種對數與X2檢驗對數相當,Spearman秩相關分析得到的極顯著相關種對明顯低于X2檢驗得到的極顯著相關種對,而顯著相關性種對高于X2檢驗得到的顯著相關種對,表明種間相關檢驗靈敏度較高,反映的信息要比種間關聯分析更全面更準確。在海草群落種間關系的研究過程中,種對間呈負相關聯結的數量明顯大于正相關數量,總體上呈現負聯結趨勢,表明海南島海草群落尚未達到穩定狀態,仍處于發展演替之中,較容易受外界因素干擾而發生波動,這與研究區域海草群落環境條件處于逐漸退化的事實相吻合。根據X2檢驗和Spearman研究結果,現將10個優勢種劃分為2個生態種組。第1組為生長在花場灣的種3(羽葉二藥)和種8(貝克喜鹽),這兩種海草存在顯著的正聯結,且都只在淤泥底質生長,碎屑砂質底質中未見分布;第2組為廣布種的種1(海神草)、種2(齒葉海神)、種4(二藥藻)、種5(針葉藻)、種7(泰萊藻)、種9(小喜鹽草)及種10(喜鹽草),這些物種相互間存在顯著的聯結與相關性,并且在淤泥底質的花場灣沒有分布。另外,種6(海菖蒲)與其他海草種類表現出很強的負相關性,可見種6(海菖蒲)具有很強的排斥其他海草種的特性,通過樣地分析發現,種6(海菖蒲)平均株冠高度為44.11 cm,是其他海草平均株高的3.5-20倍,故與種6(海菖蒲)一起生長的海草分布面積及數量都不多,可以推測,通過群落的演替,與種6(海菖蒲)一起生長的海草將會隨著種6(海菖蒲)的進一步擴增而退出該海草群落。通過對海南島海草群落種對種間種間關系的研究,從而更加了解海草群落穩定狀態,為今后開展海草的保護及修復工作提供理論依據。
Den H C,1970.The seagrasses of the world.Amsterdam:North Holland Publication,234-275.
Dennison W C,Orth R J,Moore K A,et al,1993.Assessing water quality with submersed aquatic vegetation.Bioscience,43:86-94.
蔡冰玲,范海蘭,洪偉,等,2010.梅花山擬赤楊群落樹種種群間聯結關系的研究.江西農業大學學報,32(3):0517-0522.
范航清,石雅君,邱廣龍,2009.中國海草植物.北京:海洋出版社,1-50.
郭志華,卓正大,1997.廬山常綠闊葉、落葉闊葉混交林喬木種群種間聯結性研究.植物生態學報,21(5):424-432.
韓秋影,黃小平,施平,等,2008.廣西合浦海草示范區的生態補償機制.海洋環境科學,27(3):283-286.
黃道建,黃小平,黃良民,2007.海南島新村灣營養負荷對海菖蒲的影響研究.海洋科學進展,25(2):200-207.
黃世能,李章德,駱土壽,等,2000.海南島尖峰嶺次生熱帶山地雨林樹種間的聯結動態.植物生態學報,24(5):569-574.
柯智仁,2004.臺灣海草分類與分布之研究.碩士學位論文,臺灣:國立中山大學,45-48.
李軍玲,張金屯,袁建英,2004.關帝山亞高山灌叢群落和草甸群落優勢種的種間關系.草地學報,12(2):113-119.
李先琨,黃玉清,蘇宗明,1999.南方紅豆杉群落主要樹木種群間聯結關系初步研究.生態學雜志,18(3):1-14.
歐祖蘭,李先琨,蘇宗明,等,2002.元寶山冷杉群落主要樹木種群間聯結關系的研究.生態學雜志,21(1):14-18.
閆美芳,上官鐵梁,張金屯,等,2006.五臺山藍花棘豆群落優勢種群的種間關系分析.生態學雜志,25(4):354-358.
楊宗岱,1979.中國海草植物地理學的研究.海洋湖沼通報,(2):41-46.
楊宗岱,吳寶鈴,1981.中國海草場的分布、生產力及其結構與功能的初步探討.生態學報,1(1):84-89.
尹林克,2005.塔里木河中下游地區荒漠河岸林群落種間關系分析.植物生態學報,29(2):226-234.
張金屯,2004.數量生態學.北京:科學出版社,98-99.
張景平,黃小平,2008.海草與其附生藻類之間的相互作用.生態學雜志,27(10):1785-1790.
張震云,周伶,上官鐵梁,等,2011.半干旱區錦雞兒群落優勢種種間關系.生態學雜志,30(9):1868-1874.
鄭杏雯,2007.廣西防城海草光合作用研究.南寧:碩士學位論文,廣西大學.
中國科學院中國植物志編輯委員會,1992.中國植物志.北京:科學出版社,8:84-94.
朱利君,蘇智先,王偉偉,等,2005.臥龍自然保護區三江珙桐群落種間關系的數量分析.生態學雜志,24(10):1167-1171.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:30
當代陜西(2021年17期)2021-11-06 03:21:36
當代陜西(2019年15期)2019-09-02 01:52:00
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
學苑創造·A版(2018年11期)2018-02-01 06:29:20
讀者(2017年5期)2017-02-15 18:04:18
