內分泌干擾物的非單一劑量-效應研究進展
2013-08-20 11:17:34樓欽欽田密秦占芬魏無際
生態毒理學報 2013年3期
樓欽欽,田密,秦占芬,* ,魏無際
1.南京工業大學環境學院,南京210009
2.中國科學院生態環境研究中心環境化學與生態毒理學國家重點實驗室,北京100085
劑量-效應關系是毒理學研究的核心內容之一。 經典毒理學理論認為,有毒物質在一定劑量下產生毒害作用,其劑量-效應曲線呈單調特征(monotonicity),即隨劑量的增加效應呈線性或非線性的增加或降低(圖1A-1、A-2和B-1、B-2)。近年來,隨著毒理學研究的進展,一些研究顯示除單調劑量-效應曲線外,還存在著另外一類可能更為普遍的非單調劑量-效應曲線(non-monotonic dose-response),即在一定的劑量范圍內,劑量-效應曲線的斜率可發生正負的轉換(圖1C-1、C-2和C-3),其中U型和倒U型劑量-效應曲線是最常見的2種非單調劑量-效應曲線形式。在U型劑量-效應曲線中,最大效應發生在高低劑量的兩端,隨劑量的增加斜率由負值轉為正值;倒U型劑量-效應曲線中,最大效應發生在中間劑量隨劑量的增加斜率由正值轉為負值;有的非單調劑量-效應曲線更為復雜,隨劑量的增加斜率發生多次正負的轉換。除以上這些類型的劑量-效應曲線外,還存在一種二元型(binary)的劑量-效應曲線,即在一定的閾值下不產生某一效應,當劑量到達閾值后才出現效應,但效應的大小不會隨著劑量的增加而增加。雖然毒物興奮效應(hormesis)也呈現U型或倒U型劑量-效應曲線,但一般認為毒物興奮效應是機體對壓力產生的適應性反應[1],且目前學術界對毒物興奮效應的認識還存在爭議[2-5],所以本文所討論的非單調劑量-效應不包括毒物興奮效應。
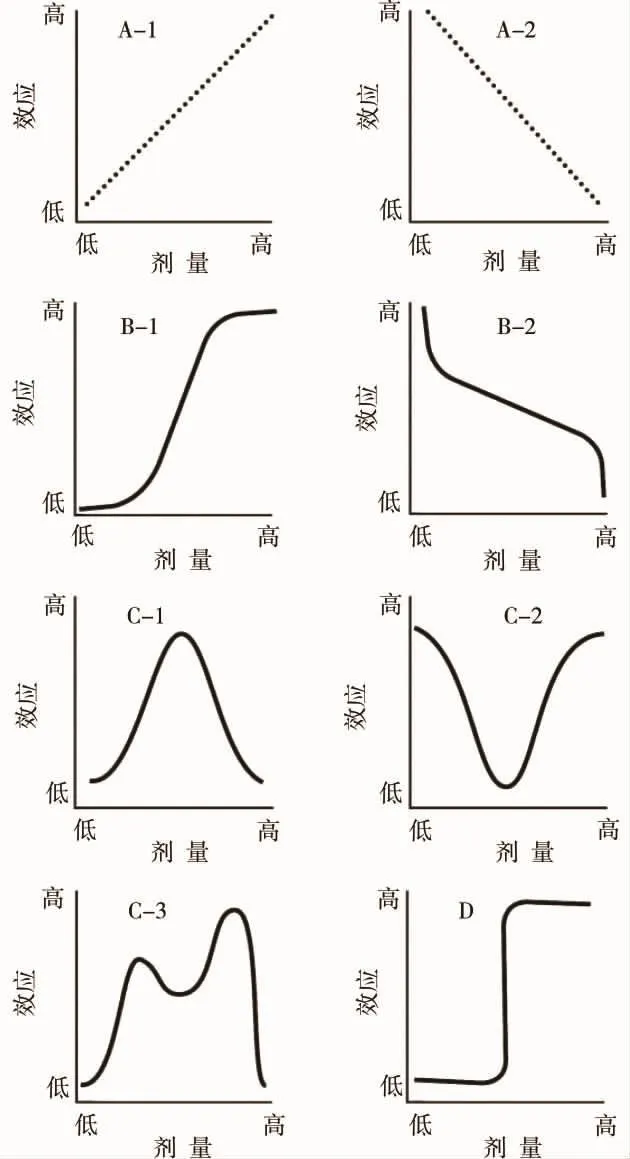
圖1 典型劑量-效應曲線[14]
雖然從20世紀60—70年代就已經有關于非單調劑量-效應的報道[6-7],但在接下來的30多年里,非單調劑量-效應并沒得到充分的關注。直到21世紀初,非單調劑量-效應才開始真正得到關注[8]。目前發現大多數激素的生物學效應都呈現非單調劑量-效應關系[9-10],相應的一些內分泌干擾物的活性也被證明具有非單調劑量-效應關系[11-12]。隨著非單調劑量-效應研究案例的不斷增加,一些科學家也開始致力于非單調劑量-效應機制的探索[13]。目前,細胞毒性、細胞和組織特異性的受體和共因子、受體選擇性、受體下調和脫敏、受體競爭、內分泌負反饋等機制可一定程度上解釋非單調劑量-效應關系[14]。非單調劑量-效應向以線性閾值劑量-效應模型為基礎的傳統毒理學理論以及現有的管理毒理學框架提出了挑戰。
本文就內分泌干擾物的非單調劑量-效應研究進展做一綜述,涉及典型的非單調劑量-效應研究案例、可能的機制以及給管理毒理學中的化學品風險評價帶來的挑戰3方面內容。
1 內分泌干擾物的非單一劑量-效應典型案例
激素對脊椎動物的生長發育以及生殖、神經、免疫等各系統結構和功能的維持起重要作用。目前已知雌激素、雄激素、孕激素、甲狀腺激素等多種激素的生物學效應具有非單調劑量-效應特征。同樣,一些具有類激素活性或干擾激素合成、代謝、轉運等過程的內分泌干擾物也具有非單調劑量-效應。下面就報道比較多的激素和內分泌干擾物的非單調劑量-效應的案例做一介紹。
1.1 激素的非單調劑量-效應案例
激素通過與細胞核、細胞質和細胞膜上的受體結合而發揮生物學效應。以雌激素為例,最早的認識是雌激素與細胞核內雌激素受體(ER)結合為復合體,復合體與雌激素靶基因上的受體反應元件(ERE)結合,從而誘導靶基因的表達,產生相應的生物學效應[15]。最近,激素的非基因活性理論也得到認可,即激素可通過細胞膜上受體而引發快速的作用模式產生生物學效應[16]。目前有越來越多的證據顯示激素對其靶細胞和靶組織的作用具有非單調劑量-效應特征。
雌激素與乳腺的生長發育有著密切的關系,Vandenberg等[17]報道雌激素在5 μg·kg-1·d-1以下的劑量可促進乳腺導管的延伸和末端形成,而在高劑量(5~50μg·kg-1·d-1)下抑制乳腺小管的延伸和末端形成。另外,雌激素也是乳腺癌發生的危險因素之一,早期研究顯示雌激素明顯誘導ER陽性的乳腺癌細胞MCF-7的增殖[18]。然而,進一步的研究證明,雌激素在低劑量(10-14~10-11mol·L-1)條件下促進MCF-7細胞增殖,在此劑量范圍內呈現單一的線性劑量-效應關系;隨著劑量的繼續增加(10-11~10-6mol·L-1),雌激素的增殖效應不再發生變化;但當劑量增加到10-6mol·L-1以上時,雌激素轉而抑制細胞增殖(圖 2)[9]。同樣,雄激素在低劑量時促進前列腺癌細胞增殖,而高劑量抑制細胞增殖,總體呈現倒U型劑量-效應曲線[19]。子宮也是雌激素作用的一個靶器官,低劑量的雌激素暴露可促進子宮增重,但當劑量增加到1mg·kg-1·d-1時,子宮增重趨勢會下降,呈現倒U型劑量效應曲線[20]。孕期低劑量的雌激素處理可促進鼠前列腺增生,而高劑量處理可抑制前列腺增生[21]。
瘦素(leptin)是參與糖代謝的一種激素,文獻顯示在不同生物學水平瘦素都可表現出非單調劑量-效應。如Zhang等[22]報道,中等劑量的瘦素會減少體外培養的脂肪細胞中胰島素介導的糖吸收,而高劑量和低劑量的瘦素能夠促進胰島素介導的糖吸收。在組織水平,瘦素對離體胰島分泌胰島素的效應影響呈倒 U性劑量-效應關系[23-24]。同樣,在個體水平瘦素也具有非單調劑量-效應特征,比如中等劑量的瘦素暴露可降低大鼠的食物吸收,而低劑量和高劑量的瘦素卻增加食物吸收[25]。以上這些證據顯示非單調劑量-效應是激素生物學效應的普遍特征。
1.2 激素相關藥物的非單一劑量-效應案例
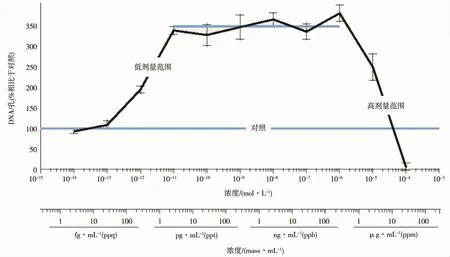
圖2 MCF-7乳腺癌細胞的增殖實驗[9]
激素類藥物是最早被關注的內分泌干擾物之一,如己烯雌酚、苯甲酸雌二醇、乙炔雌二醇、甲地孕酮等,它們通過模擬生理激素的作用達到治療的目的。三苯氧胺等藥物因為可通過與激素受體的作用而干擾生理激素的功能,也成為一類典型的內分泌干擾物。還有些藥物如甲巰咪唑,可通過影響生理激素的合成、代謝來調節體內激素的平衡以達到治療的目的。這些激素相關的藥物因為最終都要通過激素和受體的作用而產生效應,所以理論上,它們的內分泌干擾作用與生理激素類似,也具有非單調劑量-效應特征。目前有越來越多的研究結果證明確實如此。
己烯雌酚(DES)人工合成的非甾體雌激素類藥物,能產生與天然雌二醇相同的作用,主要用于雌激素低下癥、晚期乳腺癌的治療。因為能夠模擬生理雌激素的作用,己烯雌酚對乳腺、子宮、陰道等雌激素的靶器官有明顯的干擾作用,有些報道顯示這些干擾作用具有非單調劑量-效應特征。如 Shelby等[20]報道 DES 暴露未成年小鼠,10 μg·kg-1·d-1以下的劑量范圍內,隨劑量的增加促進子宮增重的效應增加,而當劑量增加到100 μg·kg-1·d-1時,子宮增重的效應明顯降低,整個劑量-效應呈現倒U型曲線。與此相似,Newbold等[26]的結果也顯示,低劑量的DES促進子宮發育相關的指標,而高劑量的DES抑制子宮發育相關的指標。另外,DES對實驗動物體質量、性別比、肛門-生殖器間距離、乳腺發育的影響也具有非單調劑量-效應關系[27-29]。
三苯氧胺是一種雌激素受體拮抗劑,最早用于晚期乳腺癌的治療,但其對乳腺癌的抑制作用嚴格依賴于在血液中濃度,一般認為,0.1~10μmol·L-1是有效的。但病人用藥開始的2周里,血液中三苯氧胺低于這個濃度,此時病人乳腺細胞的增殖非但不會被抑制反而會進一步增生;如果繼續用藥,血液中三苯氧胺升高到有效的濃度時,才會抑制乳腺癌的發展;當濃度繼續增加到10μmol·L-1以上,三苯氧胺將出現急性毒性(圖3)[14]。體外細胞研究也顯示,高劑量三苯氧胺抑制乳腺癌細胞生長,而低劑量促進乳腺癌細胞生長[30-31]。乳腺癌移植裸鼠模型研究同樣顯示,高劑量三苯氧胺抑制乳腺癌的發展,而低劑量促進乳腺癌的發展[32]。這些研究很好地證明了激素相關藥物的內分泌干擾作用具有非單調劑量-效應特征。
1.3 植物雌激素的非單一劑量-效應案例
一般認為植物雌激素是植物中具有弱雌激素/抗雌激素作用的化合物。這些化合物通過與ER以低親和度結合而發揮雌激素或抗雌激素的作用。目前有些植物雌激素被證明也可通過影響激素的合成代謝調節激素水平的平衡[33]。植物雌激素主要包括異黃酮、木酚素和香豆素3類。目前已知,雖然植物雌激素可導致某些動物不孕或雌性化[34],但對人還沒有此類不良影響,相反植物雌激素對乳腺癌、前列腺癌、心腦血管疾病、骨質疏松、絕經綜合癥等與雌激素相關的疾病有預防和治療作用[35-37]。在乳腺癌、前列腺癌等雌激素相關癌癥的防治方面,植物雌激素表現出的主要是抗雌激素作用;而在骨質疏松、絕經綜合癥等疾病的防治方面,植物雌激素表現出的主要是雌激素作用[38]。研究顯示,植物雌激素表現雌激素活性還是抗雌激素活性是與劑量相關的。
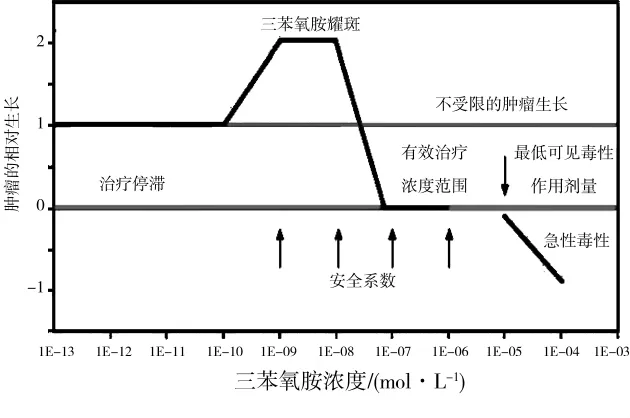
圖3 三苯氧胺在乳腺癌治療中的劑量-效應[14]Fig.3 Dose-response ranges for tamoxifen in breast cancer therapy[14]
染料木黃酮是異黃酮類植物雌激素,在不同劑量下可表現相反的效應。如Miodini等[39]曾研究染料木黃酮在0.5~20μmol·L-1對 MCF-7細胞增殖的劑量-效應關系,發現在1μmol·L-1時有最大促進值,當濃度大于5μmol·L-1時則抑制了細胞增殖。進一步研究發現,染料木黃酮在1和5μmol·L-1時抑制了ER介導的細胞生長,但卻促進了2種雌激素調節基因pS2和Cathepsin-D基因的轉錄。這些結果顯示,染料木黃酮既可表現出雌激素作用又可表現出抗雌激素作用,具體表現出什么作用取決濃度,另外與檢測的終點指標也相關。因此在一定劑量范圍內,染料木黃酮表現出非單調劑量-效應曲線[40-41]。
除染料木黃酮外,研究結果顯示玉米赤霉醇、柚皮素、雞豆黃素、香豆素、槲皮素等植物雌激素都可表現出非單調劑量-效應曲線[42-45],并且非單調劑量-效應關系可能是植物雌激素的普遍特征。
1.4 農藥類內分泌干擾物的非單一劑量-效應案例
目前為止,很多農藥被證明具有內分干擾作用,如滴滴涕/滴滴伊(DDT/DDE)、阿特拉津、西瑪津、毒死蜱、甲氧氯、乙稀菌核利、馬拉硫磷等等。農藥已經成為內分泌干擾物中不可被忽視的一類。而且,這些農藥類內分泌干擾物中的大部分已被證明具有非單一劑量-效應關系。如Wozniak等[46]報道DDE 在10-12~10-8mol·L-1的劑量范圍內,誘導垂體瘤細胞分泌催乳素的效應呈倒U型曲線,另一種農藥硫丹也表現出相似的劑量-效應曲線。Ohlsson等[47]研究了咪唑類抗真菌劑咪酰胺對腎上腺皮質瘤細胞H295R激素分泌的影響,結果發現在0.01~10μmol·L-1的濃度范圍內,除皮質醇的分泌隨濃度升高而下降外,醛固酮、皮質酮、孕酮、17α-羥基-孕酮的分泌隨劑量的變化而呈現倒U型曲線。在動物實驗中,目前積累的農藥類內分泌干擾物的非單調劑量-效應的案例也很多。
阿特拉津是一種倍受爭議的內分泌干擾物,以它對兩棲動物變態的影響為例,一些研究顯示對變態發育沒有影響[48-49],而另外一些結果顯示對變態有影響[50-52],在這些有影響的結果中,有的顯示抑制作用[52],有的顯示促進作用[51]。另外,也有研究顯示阿特拉津對兩棲動物變態的影響可能是非單調劑量-效應,如 Larson 等[53]報道75 μg·L-1的阿特拉津處理抑制虎皮蠑螈變態,而250μg·L-1時卻促進變態。Brodeur等[54]報道阿特拉津在1 mg·L-1時促進南美蟾蜍變態,而5 mg·L-1時抑制變態,其他劑量時效應不明顯。最近Rohr等[55]用變態分析的方法對文獻中阿特拉津對兩棲動物變態影響的結果進行了分析,結果認為阿特拉津可以影響兩棲動物變態,根據劑量的不同,既可表現為促進作用也可變現為抑制作用,也就是說阿特拉津對兩棲動物變態的影響具有的非單調劑量-效應關系。以上阿特拉津的研究證明了農藥類內分泌干擾物劑量-效應關系的復雜性。
1.5 工業化學品類內分泌干擾物的非單一劑量-效應案例
大量研究顯示雙酚A、壬基酚、酞酸酯、多氯聯苯(PCBs)、多溴二苯醚(PBDEs)等工業生產或生活中廣泛應用的一些化學品具有內分泌干擾作用,而其中一些內分泌干擾作用表現為非單調劑量-效應關系。
雙酚A是最早被關注也是目前研究最多的一種內分泌干擾物,它的內分泌干擾作用涉及生殖內分泌、甲狀腺以及相關的神經發育等多個系統[56-57]。這些內分泌干擾作用都不同程度上表現出非單調劑量-效應。如Markey等[58]報道,高劑量(100 mg·kg-1·d-1)和低劑量(0.1 mg·kg-1·d-1)的雙酚A處理都能導致小鼠陰道開放,而中間劑量(0.5~75 mg·kg-1·d-1)雙酚A不影響鼠的陰道開放。與此一致,Cabaton 等[59]報道高劑量(25 μg·kg-1·d-1)和低劑量(25 ng·kg-1·d-1)的雙酚A降低雌鼠的生殖能力,而中間劑量250 ng·kg-1·d-1對雌鼠的生殖能力沒有明顯影響。Jones等[60]的報道則相反,雙酚A對雄性鼠性行為的影響僅發生在中間劑量(50μg·kg-1·d-1),而高劑量和低劑量的雙酚A處理沒有效應。最近這個研究組又報道,雙酚A對鼠神經行為的影響也呈非單調劑量-效應特征[61]。
PCBs是另外一種被研究較多的內分泌干擾物,其內分泌干擾作用的非單調劑量效應關系也在很多研究中得到證明。如Muto等[62]報道二甲基苯蒽誘導大鼠乳腺癌模型中,以250 ng·kg-1的劑量注射PCB126可明顯促進乳腺癌的發生,而高劑量(7.5 μg·kg-1)處理明顯抑制乳腺癌的發生。進一步的在分子水平上的研究也證明了PCB126對乳腺癌發生的非單調劑量-效應關系[62-64]。值得注意的是,PCBs內分泌干擾作用的非單調劑量-效應關系不僅在動物實驗中得到證明,一些流行病學的數據也支持PCBs對人體健康作用也存在非單調的劑量-效應關系。如Lee等[65-66]發現一些高氯代PCB同類物與糖尿病具有非線性的劑量-效應相關性。有報道顯示,一些PCB同類物與DNA甲基化也有類似的相關性[67]。
除雙酚A和PCBs外,壬基酚、辛基酚、酞酸酯和三氯生等很多內分泌干擾物都被證實具有非單調劑量-效應關系,越來越多的證據顯示內分泌干擾物的非單調劑量-效應具有普遍性。
2 內分泌干擾物的非單調劑量-效應關系機制
隨著內分泌干擾物非單調劑量-效應研究案例的不斷增加,這一與經典的線性劑量-效應關系不一致的劑量-效應曲線已逐漸被接受,并認為非單調劑量-效應關系可能是內分泌干擾效應更為普遍的劑量-效應關系。因此,一些科學家開始致力于內分泌干擾物非單調劑量-效應關系機制的研究。
2.1 細胞毒性
一些環境物質的內分泌干擾活性是基于體外細胞實驗獲得的,如評價雌激素活性的乳腺癌MCF-7(或TD47)細胞增殖實驗、評價雄激素活性的前列腺癌LNCaP-FGC細胞增殖實驗等。在這些體外細胞實驗中,一些內分泌干擾活性表現出的非單調劑量-效應關系可部分地用細胞毒性解釋,即低劑量下表現出的對細胞增殖的促進作用是激素受體介導的作用,是真正的內分泌干擾活性的表現;而高劑量時表現的對細胞的抑制作用則是細胞毒性的結果,與內分泌干擾活性無關。但在整個測試劑量范圍內,內分泌干擾物對細胞增殖的影響整體上呈現倒U型劑量-效應曲線。如Welshons等[9]報道的雌激素在10-14~10-11mol·L-1時促進 MCF-7細胞增殖,劑量-效應呈單一線性關系;隨著劑量增加到10-11~10-6mol·L-1時,雌激素的增殖效應不再變化;當劑量增加到10-6mol·L-1以上,雌激素轉而抑制細胞增殖。
細胞毒性除部分地解釋一些細胞實驗中的非單調劑量-效應關系外,可能還能夠在一定程度上外推到一些體內實驗中,即高劑量的抑制結果是急性毒性的反映,而不是內分泌干擾效應。這一推測還需要進一步驗證。
2.2 兩個(或以上)相反單調效應的疊加
某些非單調劑量-效應曲線可能是兩個(或以上)相反的單調效應共同作用的結果。如有研究顯示,某些激素對細胞增殖的非單調劑量-效應曲線是因為細胞中存在對激素具有不同響應的兩種類型的細胞,一種細胞表現為接受激素而增殖,而另一種細胞表現為接受激素而凋亡,兩種效應疊加而出現非單調劑量-效應曲線[68]。如體外培養的乳腺癌MCF-7細胞中有一類細胞可接受雌激素的誘導而增殖,另外一類細胞可在雌激素的作用下凋亡[69]。于是,在一定劑量范圍內,兩種效應疊加時可能出現非單調劑量-效應曲線。
2.3 受體多樣性和選擇性
細胞或組織中通常存在與配體親和能力不同的兩種或更多的受體亞型,而不同的受體亞型介導的效應通常相反[70-71]。低濃度時具有高親和力的受體亞型發揮作用,高濃度時低親和力的受體亞型發揮作用,并且這時低親和力受體的高容量將變得相當重要。當在一個較寬的劑量范圍內評價內分泌干擾物活性時,就會出現典型的非單一劑量-效應。以子宮組織中ER為例,ERα介導細胞增殖反應,而ERβ介導細胞凋亡[72]。由于內分泌干擾物與這兩種ER的結合能力存在差異,所以由兩種受體介導的相反的單調效應共同作用而產生非單調劑量-效應。
另外,一些內分泌干擾物可以與多種激素受體結合,但是與各種受體的親和力存在差異,所以在不同劑量下會表現為與受體的選擇性結合。比如雙酚A,與ER的親和力最強,在低劑量時只能和ER結合;但在高劑量時,雙酚 A也能與 AR、TR等受體結合[73-74]。也就是說,一些內分泌干擾物在高劑量時表現出的某一活性可能是由多種受體介導的,而在低劑量下的活性可能是由一種受體介導的。因此,在比較寬泛的劑量范圍內,內分泌干擾物對某一終點指標的影響可能呈現復雜的非單調劑量-效應關系。
2.4 受體競爭
有觀點認為,內分泌干擾物在非單一劑量-效應中,高劑量效應和低劑量效應形成的機制是不同的[9]。低劑量時,內分泌干擾物對機體的擾動處于激素的調控活性范圍之內,劑量上升能夠通過正常的受體介導途徑使得效應增加。當內分泌干擾物濃度遠大于生理激素濃度時,受體結合出現飽和,較大的劑量改變僅能引起較小的生物學效應的變化或不能引起生物學效應的變化。而數學模型顯示,當有內源性激素存在時,內分泌干擾物可與非占位的激素受體結合,導致生物學效應的增加[75]。在低劑量時,內分泌干擾物和內源性激素都能和激素受體結合而激發生物學效應;而在高劑量時,內分泌干擾物與受體的結合超過內源性激素與受體的結合。因為內分泌干擾物的活性低于生理激素的活性,所以當內分泌干擾物的劑量升高時,總體的效應會下降,因而呈現倒U型劑量-效應曲線。
2.5 受體下調和受體脫敏
激素配體與受體的結合最終導致靶基因轉錄的變化,之后受體通過某些途徑降解或脫敏,從而調節生物學效應的變化[74,76]。在一些受體降解過程中,激素配體本身起重要作用。如雌激素、孕激素和糖皮質激素介導相應受體的降解,而雄激素的結合阻止雄激素受體的降解[77-78],另外,一些激素受體的降解依賴于其他激素配體的參與[79]。也就是說,隨激素或內分泌干擾物劑量的變化,其生物學的效應也在發生變化,反過來又影響激素受體數量的變化。脫敏是另外一條調節激素配體與受體結合導致生物學效應的途徑,即不是通過受體降解減少受體數目而是通過減小受體對激素配體的反應性來調節生物學效應。目前有研究發現,人絨毛膜促性腺激素、促卵泡激素、前列腺素等激素的受體都存在脫敏的現象[80]。生物學的研究顯示,激素濃度與結合受體的數目之間、結合受體的數目與生物學效應之間的關系呈非線性關系[9]。所以,受體下調和脫敏可能部分參與了某些內分泌干擾物的非單調劑量-效應。
2.6 其他機制
以上僅是幾種導致內分泌干擾物非單調劑量-效應可能的機制。由于內分泌系統及其調節的復雜性,可能還存在其他的導致非單調劑量-效應的機制。如激素的反饋調節,很多激素的合成分泌受一系列的正負反饋調節,通過這些調節激素被控制在一個相對穩定的水平。在反饋調節的過程中,可能會出現不同時間內的生物學效應,因此出現非單調劑量-效應曲線。
總之,內分泌干擾物的非單調-劑量效應是普遍的現象,導致這一復雜的劑量-效應曲線的機制還需進一步探索。
3 非單調劑量-效應對傳統管理毒理學的挑戰
在傳統管理毒理學中,所謂化學品的安全劑量是基于線性閾值劑量-效應曲線獲得的,即毒性實驗從高劑量做起,獲得最低可觀察效應劑量(LOAEL)和無可觀察效應劑量(NOAEL),NOAEL除以一個安全系數(3~1000),即為安全劑量(參考劑量)。但目前存在的化學品的安全劑量是否真的安全,很少有實驗驗證過。與目前發現的內分泌干擾物的非單調劑量-效應曲線比較來看,這種基于線性閾值劑量-效應曲線獲得的安全劑量存在很大問題(圖4),在所謂的安全劑量下會漏掉潛在的風險。另外,有觀點認為從內分泌干擾效應是相對體內穩定的內分泌系統而言,即使很低劑量的內分泌干擾物也能對生理的內分泌系統產生干擾作用,所以內分泌干擾物像致癌物一樣不存在安全閾值[81-82]。因此,內分泌干擾物的非單調劑量-效應對傳統的管理毒理學提出了空前的挑戰。
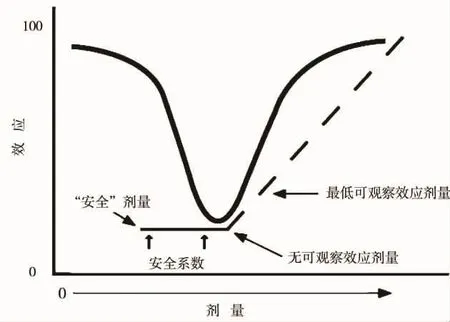
圖4 非單調劑量-效應曲線和作為計算安全劑量理論基礎的線性劑量-效應曲線[14]Fig.4 Non-monotonic dose-response curve and linear dose-response curve as the theoretical base for calculating the safe dose[14]
為應對非單調劑量-效應關系的挑戰,一種觀點認為今后的內分泌干擾研究應設計包含環境相關劑量在內的更為寬泛的劑量范圍,探索更為精細的終點評價指標,選擇更為合理的暴露時間和檢查終點指標的時間,而直接揭示相對低劑量下的內分泌干擾效應,而不是從高劑量的結果推導低劑量的效應[14,83]。盡管這種觀點的呼聲很高,但目前包括內分泌干擾在內的毒理學研究仍以高劑量為主,因為一般認為低劑量暴露不容易產生明顯的毒理學效應。另一種觀點認為,盡管在比較寬泛的劑量范圍內存在非單調劑量-效應關系,但不能否定高劑量在毒理學研究中的作用,建立在科學基礎上的由高劑量到低劑量的外推是必要的[13]。Conolly 和 Lutz[13]主張結合系統生物學研究的成果,探索新的適合于非單調劑量-效應關系的外推模型。
盡管有關內分泌干擾物非單調劑量-效應關系的報道越來越多,非單調劑量-效應關系也引起學術界的關注,但與線性的劑量-效應研究相比,目前對非單調劑量-效應的認識還很膚淺。所以,無論是從優化實驗設計直接揭示從低劑量到高劑量的非單調劑量-效應的角度,還是從建立由高劑量到低劑量的新的外推模型的角度,開展非單調劑量-效應這一嶄新領域的研究都是必要的。
[1] Calabrese E J,Baldwin L A.Defining hormesis[J].Human& Experimental Toxicology,2002,21(2):91-97
[2] Calabrese E J,Baldwin L A,Kostecki P T,et al.A toxicologically based weight-of-evidencemethodology for the relative ranking of chemicals of endocrine disruption potential[J].Regulatory Toxicology and Pharmacology,1997,26(1):36-40
[3] Calabrese E J,Baldwin L A.Hormesis as a biological hypothesis [J].Environmental Health Perspectives,1998,106(1):357-362
[4] Mushak P.Hormesis and its place in nonmonotonic dose-response relationships:Some scientific reality checks[J].Environmental Health Perspectives,2007,115(4):500-506
[5] Kendig E L,Le H H,Belcher SM.Defining hormesis:Evaluation of a complex concentration response phenomenon[J].International Journal of Toxicology,2010,29(3):235-246
[6] Young E D,Brownell W E.Responses to tones and noise of single cells in dorsal cochlear nucleus of unanesthetized cats [J].Journal of Neurophysiology,1976,39(2):282-300
[7] Bliznakov EG.Immunostimulation or immunodepression[J].Biomedicine,1977,26(2):73-76
[8] Melnick R,Lucier G,Wolfe M,et al.Summary of the National Toxicology Program's report of the endocrine disruptors low-dose peer review [J].Environmental Health Perspectives,2002,110(4):427-431
[9] Welshons W V,Thayer K A,Judy B M,et al.Large effects from small exposures.I.Mechanisms for endocrine-disrupting chemicals with estrogenic activity [J].Environmental Health Perspectives,2003,111(8):994-1006
[10] Fallah N,Mohammad K,Nourijelyani K,etal.Nonlinear association between serum testosterone levels and coronary artery disease in Iranian men[J].European Journal of Epidemiology,2009,24(6):297-306
[11] Endo F,Monsees TK,Akaza H,etal.Effects of single non-ortho,mono-ortho,and di-ortho chlorinated biphenyls on cell functions and proliferation of the human prostatic carcinoma cell line,LNCaP [J].Reproductive Toxicology,2003,17(2):229-236
[12] Jenkins S,Wang J,Eltoum I,et al.Chronic oral exposure to bisphenol A results in a nonmonotonic dose response in mammary carcinogenesis and metastasis in MMTV-erbB2 mice[J].Environmental Health Perspectives,2011,119(11):1604-1609
[13] Conolly R B,Lutz W K.Nonmonotonic dose-response relationships:Mechanistic basis,kinetic modeling,and implications for risk assessment[J].Toxicological Sciences,2004,77(1):151-157
[14] Vandenberg L N,Colborn T,Hayes T B,et al.Hormones and endocrine-disrupting chemicals:Low-dose effects and nonmonotonic dose responses[J].Endocrine Reviews,2012,33(3):378
[15] Brown M,Sharp P A.Human estrogen receptor forms multiple protein-DNA complexes[J].The Journal of Biological Chemistry,1990,265(19):11238-11243
[16] Watson C S,Alyea R A,Jeng Y J,et al.Nongenomic actions of low concentration estrogens and xenoestrogens on multiple tissues[J].Molecular and Cellular Endocrinology,2007,274(1-2):1-7
[17] Vandenberg L N,Wadia P R,Schaeberle C M,et al.Themammary gland response to estradiol:Monotonic at the cellular level,non-monotonic at the tissue-level of organization?[J].Journal of Steroid Biochemistry and Molecular Biology,2006,101(4-5):263-274
[18] Soto A M,Sonnenschein C,Chung K L,et al.The EScreen assay as a tool to identify estrogens—An update on estrogenic environmental-pollutants[J].Environmental Health Perspectives,1995,103(s7):113-122
[19] Geck P,Szelei J,Jimenez J,et al.Expression of novel genes linked to the androgen-induced,proliferative shutoff in prostate cancer cells[J].Journal of Steroid Biochemistry and Molecular Biology,1997,63(4-6):211-218
[20] Shelby M D,Newbold R R,Tully D B,etal.Assessing environmental chemicals for estrogenicity using a combination of in vitro and in vivo assays[J].Environmental Health Perspectives,1996,104(12):1296-1300
[21] vomSaal F S,Timms B G,Montano M M,et al.Prostate enlargement inmice due to fetal exposure to low doses of estradiol or diethylstilbestrol and opposite effects at high doses[J].Proceedings of the National Academy of Sciences,1997,94(5):2056-2061
[22] Zhang H H,Kumar S,Barnett A H,et al.Intrinsic site-specific differences in the expression of leptin in human adipocytes and its autocrine effects on glucose uptake[J].Journal of Clinical Endocrinology& Metabolism,1999,84(7):2550-2556
[23] Pallett A L,Morton N M,Cawthorne M A,etal.Leptin inhibits insulin secretion and reduces insulinmRNA levels in rat isolated pancreatic islets[J].Biochemical and Biophysical Research Communications,1997,238(1):267-270
[24] Haddad N,Howland R,Baroody G,etal.Themodulatory effect of leptin on the overall insulin production in ex-vivo normal rat pancreas[J].Canadian Journal of Physiology and Pharmacology,2006,84(2):157-162
[25] Thorburn AW,Holdsworth A,Proietto J,etal.Differential and genetically separable associations of leptin with obesity-related traits[J].International Journal of Obesity,2000,24(6):742-750
[26] Newbold R R,Jefferson W N,Padilla-Banks E,et al.Developmental exposure to diethylstilbestrol(DES)alters uterine response to estrogens in prepubescentmice:Low versus high dose effects[J].Reproductive Toxicology,2004,18(3):399-406
[27] Palanza P,Parmigiani S,vom Saal F S.Effects of prenatal exposure to low doses of diethylstilbestrol,o,p'DDT,and methoxychlor on postnatal growth and neurobehavioral development inmale and femalemice[J].Hormones and Behavior,2001,40(2):252-265
[28] Utsuyama M,Kanno J,Inoue T,et al.Age/sex dependent and non-monotonous dose-response effect of diethylstilbestrol on the immune functions in mice[J].Toxicology Letters,2002,135(1-2):145-153
[29] Kohlerova E,Skarda J.Mouse bioassay to assess oestrogenic and anti-oestrogenic compounds:Hydroxytamoxifen,diethylstilbestrol and genistein [J].Journal of Veterinary Medicine Series A-Physiology Pathology Clinical Medicine,2004,51(5):209-217
[30] Reddel R R,Sutherland R L.Tamoxifen stimulation of human breast cancer cell proliferation in vitro:A possible model for tamoxifen tumour flare[J].European Journal of Cancer& Clinical Oncology,1984,20(11):1419-1424
[31] Berthois Y,Pons M,Dussert C,et al.Agonist-antagonistactivity of anti-estrogens in the human breast cancer cell line MCF-7:An hypothesis for the interaction with a site distinct from the estrogen binding site[J].Molecular and Cellular Endocrinology,1994,99(2):259-268
[32] Osborne C K,Hobbs K,Clark GM.Effectof estrogens and antiestrogens on growth of human-breast cancer-cells in athymic nude-mice[J].Cancer Research,1985,45(2):584-590
[33] Lacey M,Bohday J,Fonseka SM,etal.Dose-response effects of phytoestrogens on the activity and expression of 3beta-hydroxysteroid dehydrogenase and aromatase in human granulosa-luteal cells[J].The Journal of Steroid Biochemistry and Molecular Biology,2005,96(3-4):279-286
[34] 從琳,秦占芬,周景明,等.植物雌激素對動物和人體健康的影響[J].環境與健康雜志,2006,23(2):176-178 Cong L,Qin Z F,Zhou JM,et al.Effects of phytoestrogens on the health of human and animals[J].Journal of Environment and Health,2006,23(2):176-178(in Chinese)
[35] Ganry O.Phytoestrogen and breast cancer prevention[J].European Journal of Cancer Prevention,2002,11(6):519-522
[36] Altavilla D,Crisafulli A,MariniH,etal.Cardiovascular effects of the phytoestrogen genistein[J].Current Medicinal Chemistry-Cardiovascular&Hematological Agents,2004,2(2):179-186
[37] Szkutnik-Fiedler D,Jedrzejczyk M,Grzeskowiak E,et al.The role of phytoestrogen therapy in relieving postmenopausal symptoms[J].Ginekologia Polska,2010,81(12):929-934
[38] Bingham SA,Atkinson C,Liggins J,et al.Phyto-oestrogens:Where are we now?[J].British Journal of Nutrition,1998,79(5):393-406
[39] Miodini P,Fioravanti L,Di Fronzo G,et al.The two phyto-oestrogens genistein and quercetin exert different effects on oestrogen receptor function[J].British Journal of Cancer,1999,80(8):1150-1155
[40] Chen A C,Donovan SM.Genistein at a concentration present in soy infant formula inhibits Caco-2BBe cell proliferation by causing G2/M cell cycle arrest[J].Journal of Nutrition,2004,134(6):1303-1308
[41] El Touny L H,Banerjee P P.Identification of a biphasic role for genistein in the regulation of prostate cancer growth andmetastasis[J].Cancer Research,2009,69(8):3695-3703
[42] Collins B M,McLachlan J A,Arnold S F.The estrogenic and antiestrogenic activities of phytochemicalswith human estrogen receptor expressed in yeast[J].Steroids,1997,62(4):365-372
[43] Almstrup K,Fernandez M F,Petersen JH,et al.Dual effects of phytoestrogens result in u-shaped dose-response curves[J].Environmental Health Perspectives,2002,110(8):743-748
[44] Yuri T,Tsukamoto R,Miki K,etal.Biphasic effects of zeranol on the growth of estrogen receptor-positive human breast carcinoma cells [J].Oncology Reports,2006,16(6):1307-1312
[45] Liu M,Yanagihara N,Toyohira Y,et al.Dual effects of daidzein,a soy isoflavone,on catecholamine synthesis and secretion in cultured bovine adrenal medullary cells[J].Endocrinology,2007,148(11):5348-5354
[46] Wozniak A L,Bulayeva N N,Watson C S.Xenoestrogens at picomolar to nanomolar concentrations trigger membrane estrogen receptor-alpha-mediated Ca2+fluxes and prolactin release in GH3/B6 pituitary tumor cells[J].Environmental Health Perspectives,2005,113(4):431-439
[47] Ohlsson A,Ulleras E,Oskarsson A.A biphasic effectof the fungicide prochloraz on aldosterone,but not cortisol,secretion in human adrenal H295R cells—Underlyingmechanisms[J].Toxicology Letters,2009,191(2-3):174-180
[48] Du Y,Shi X,Liu C,et al.Chronic effects of waterborne PFOSexposure on growth,survival and hepatotoxicity in zebrafish:A partial life-cycle test[J].Chemosphere,2009,74(5):723-729
[49] Oka T,Tooi O,Mitsui N,et al.Effect of atrazine on metamorphosis and sexual differentiation in Xenopus laevis[J].Aquatic Toxicology,2008,87(4):215-226
[50] Freeman J L,Rayburn A L.Developmental impact of atrazine on metamorphing Xenopus laevis as revealed by nuclear analysis and morphology[J].Environmental Toxicology and Chemistry,2005,24(7):1648-1653
[51] Forson D,Storfer A.Effects of atrazine and iridovirus infection on survival and life-history traits of the long-toed salamander(Ambystoma macrodactylum)[J].Environmental Toxicology and Chemistry,2006,25(1):168-173
[52] Langlois V S,Carew A C,Pauli B D,et al.Low levels of the herbicide atrazine alter sex ratios and reducemetamorphic success in Rana pipiens tadpoles raised in outdoor mesocosms[J].Environmental Health Perspectives,2010,118(4):552-557
[53] Larson D L,McDonald S,Fivizzani A J,et al.Effects of the herbicide atrazine on Ambystoma tigrinummetamorphosis:Duration,larval growth,and hormonal response[J].Physiological Zoology,1998,71(6):671-679
[54] Brodeur JC,Svartz G,Perez-Coll CS,etal.Comparative susceptibility to atrazine of three developmental stages of Rhinella arenarum and influence on metamorphosis:Non-monotonous acceleration of the time to climax and delayed tail resorption [J].Aquatic Toxicology,2009,91(2):161-170
[55] Rohr JR,McCoy K A.A qualitativemeta-analysis reveals consistent effects of atrazine on freshwater fish and amphibians [J].Environmental Health Perspectives,2010,118(1):20-32
[56] Golub M S,Wu K L,Kaufman F L,et al.Bisphenol A:Developmental toxicity from early prenatal exposure[J].Birth Defects Research Part B-Developmental and Reproductive Toxicology,2010,89(6):441-466
[57] Rubin B S.Bisphenol A:An endocrine disruptor with widespread exposure and multiple effects[J].The Journal of Steroid Biochemistry and Molecular Biology,2011,127(1-2):27-34
[58] Markey C M,Michaelson C L,Veson E C,et al.The mouse uterotrophic assay:A reevaluation of its validity in assessing the estrogenicity of bisphenol A[J].Environmental Health Perspectives,2001,109(1):55-60
[59] Cabaton N J,Wadia PR,Rubin B S,et al.Perinatal exposure to environmentally relevant levels of bisphenol A decreases fertility and fecundity in CD-1 mice[J].Environmental Health Perspectives,2011,119(4):547-552
[60] Jones B A,Shimell JJ,Watson N V.Pre-and postnatalbisphenol A treatment results in persistent deficits in the sexual behavior ofmale rats,but not female rats,in adulthood[J].Hormones and Behavior,2011,59(2):246-251
[61] Jones B A,Watson N V.Perinatal BPA exposure demasculinizes males in measures of affect but has no effect on water maze learning in adulthood[J].Hormones and Behavior,2012,61(4):605-610
[62] Muto T,Wakui S,Imano N,etal.Mammary gland differentiation in female rats after prenatal exposure to 3,3',4,4',5-pentachlorobiphenyl[J].Toxicology,2002,177(2-3):197-205
[63] Wakui S,Yokoo K,TakahashiH,etal.CYP1 and AhR expression in 7,12-dimethylbenz[a]anthracene-induced mammary carcinoma of rats prenatally exposed to3,3,4,4',5-pentachlorobiphenyl[J].Toxicology,2005,211(3):231-241
[64] Wakui S,Yokoo K,TakahashiH,etal.Prenatal3,3',4,4 ',5-pentachlorobiphenyl exposure modulates induction of rat hepatic CYP1A1,1B1,and AhR by 7,12-dimethylbenz[a]anthracene [J].Toxicology and Applied Pharmacology,2006,210(3):200-211
[65] Lee D H,Steffes M W,Sjodin A,et al.Low dose of some persistent organic pollutants predicts type 2 diabetes:A nested case-control study[J].Environmental Health Perspectives,2010,118(9):1235-1242
[66] Lee D H,Steffes MW,Sj?din A,et al.Low dose organochlorine pesticides and polychlorinated biphenyls predict obesity,dyslipidemia,and insulin resistance among people free of diabetes[J].PLoSONE,2011,6(1):e15997
[67] Kim H,Jeong JH,Hwang JH,et al.Enhancement of immunostimulation and anti-metastasis in submerged culture of bearded tooth mushroom(hericium erinaceum)mycelia by addition of ginseng extract[J].Food Science and Biotechnology,2010,19(5):1259-1266
[68] Soto A M,Sonnenschein C.The two faces of Janus:Sex steroids as mediators of both cell proliferation and cell death[J].Journal of the National Cancer Institute,2001,93(22):1673-1675
[69] Song R X D,Mor G,Naftolin F,et al.Effect of longterm estrogen deprivation on apoptotic responses of breast cancer cells to17 beta-estradiol[J].Journalof the National Cancer Institute,2001,93(22):1714-1723
[70] Heldring N,Pike A,Andersson S,et al.Estrogen receptors:How do they signal and what are their targets[J].Physiological Reviews,2007,87(3):905-931
[71] Li L,Andersen M E,Heber S,et al.Non-monotonic dose-response relationship in steroid hormone receptormediated gene expression[J].Journal ofMolecular Endocrinology,2007,38(5):569-585
[72] Morani A,Warner M,Gustafsson JA.Biological functions and clinical implications of oestrogen receptors alfa and beta in epithelial tissues[J].Journal of Internal Medicine,2008,264(2):128-142
[73] Moriyama K,Tagami T,Akamizu T,etal.Thyroid hor-mone action is disrupted by bisphenol A as an antagonist[J].Journal of Clinical Endocrinology& Metabolism,2002,87(11):5185-5190
[74] Ismail A,Nawaz Z.Nuclear hormone receptor degradation and gene transcription:An update[J].International Union of Biochemistry and Molecular Biology Life,2005,57(7):483-490
[75] Kohn M C,Melnick R L.Biochemical origins of the nonmonotonic receptor-mediated dose-response[J].Journal of Molecular Endocrinology,2002,29(1):113-123
[76] Lonard DM,O'Malley BW.Emerging roles of the ubiquitin proteasome system in nuclear hormone receptor signaling[J].Progress in Molecular Biology and Translational Science,2009,87:117-135
[77] Nawaz Z,Lonard D M,Dennis A P,etal.Proteasomedependent degradation of the human estrogen receptor[J].Proceedings of the National Academy of Sciences,1999,96(5):1858-1862
[78] Lin H K,Altuwaijri S,Lin W J,et al.Proteasome activity is required for androgen receptor transcriptional activity via regulation of androgen receptor nuclear translocation and interaction with coregulators in prostate cancer cells [J].Journal of Biological Chemistry,2002,277(39):36570-36576
[79] Kinyamu H K,Archer T K.Estrogen receptor-dependent proteasomal degradation of the glucocorticoid receptor is coupled to an increase in Mdm2 protein expression[J].Molecular and Cellular Biology,2003,23(16):5867-5881
[80] Shankaran H,Wiley H S,ResatH.Receptor downregulation and desensitization enhance the information processing ability of signalling receptors[J].BMC Systems Biology,2007,1(9):48
[81] Sakaue M,Ohsako S,Ishimura R,et al.Bisphenol-A affects spermatogenesis in the adult rat even at a low dose[J].Journal of Occupational Health,2001,43:185-190
[82] Gupta C.Reproductive malformation of the male offspring following maternal exposure to estrogenic chemicals[J].Proceedings of the Society for Experimental Biology and Medicine,2000,224(2):61-68
[83] Myers JP,Zoeller R T,vom Saal F S.A clash of old and new scientific concepts in toxicity,with important implications for public health [J].Environmental Health Perspectives,2009,117(11):1652-1655 ◆
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中老年保健(2022年6期)2022-08-19 01:41:48
核科學與工程(2021年4期)2022-01-12 06:30:26
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
今日農業(2020年19期)2020-12-14 14:16:52
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
祝您健康(2018年5期)2018-05-16 17:10:16
中學物理·高中(2016年12期)2017-04-22 11:53:03
