2008—2011年北京協(xié)和醫(yī)院流感嗜血桿菌耐藥性監(jiān)測(cè)
2013-08-23 03:01:38楊啟文
中國(guó)感染與化療雜志 2013年6期
關(guān)鍵詞:耐藥
牛 翠,張 輝,楊啟文,李 娜
2.邢臺(tái)市第三醫(yī)院檢驗(yàn)科,河北邢臺(tái) 054000。
流感嗜血桿菌(Haemophilus influenzae)是呼吸道感染的重要病原菌之一,可通過(guò)呼吸道在人與人之間傳播。也可由鼻咽部侵入血流,導(dǎo)致多種感染性疾病如化膿性腦膜炎、小兒肺炎和血流感染、成人社區(qū)獲得性肺炎、中耳炎、慢性支氣管炎急性發(fā)作和急性鼻竇炎等。近年來(lái)由于抗菌藥物的廣泛應(yīng)用,流感嗜血桿菌對(duì)抗菌藥物的耐藥率呈上升趨勢(shì),給臨床抗感染治療帶來(lái)了許多困難。現(xiàn)對(duì)北京協(xié)和醫(yī)院2008—2011年臨床分離的流感嗜血桿菌的耐藥情況作一分析,以指導(dǎo)臨床采取有效的感染控制措施及合理用藥,報(bào)道如下。
材料與方法
一、材料
(一)菌株來(lái)源 收集2008年1月至2011年12月北京協(xié)和醫(yī)院臨床標(biāo)本分離的流感嗜血桿菌,剔除同一患者相同部位的重復(fù)分離菌株后共獲223株。
(二)抗菌藥物紙片 氨芐西林、氨芐西林-舒巴坦、阿莫西林-克拉維酸、頭孢呋辛、頭孢曲松、頭孢噻肟、頭孢克洛、氨曲南、環(huán)丙沙星、左氧氟沙星、莫西沙星、甲氧芐啶-磺胺甲口惡唑、氯霉素、阿奇霉素、四環(huán)素藥敏紙片和V、X因子、V+X因子紙片均為英國(guó)OXOID公司產(chǎn)品。測(cè)定β內(nèi)酰胺酶的頭孢硝噻吩紙片為美國(guó)BBL公司產(chǎn)品。
(三)培養(yǎng)基 流感嗜血桿菌選擇性培養(yǎng)基以哥倫比亞瓊脂(OXOID公司產(chǎn)品)為基礎(chǔ)制備巧克力平皿,澆注平皿前(約50℃)加入萬(wàn)古霉素(50 mg/L)。流感嗜血桿菌藥敏培養(yǎng)基(HTM)和營(yíng)養(yǎng)補(bǔ)充劑SR0158E為英國(guó)OXOID公司產(chǎn)品。
(四)VITEK2-NH 試卡 VITEK-2-NH 試卡由法國(guó)生物梅里埃公司提供。
二、方法
(一)菌株分離及鑒定 送檢標(biāo)本立即接種血平皿和加萬(wàn)古霉素巧克力平皿,置于35℃,5%~10%CO2培養(yǎng)24 h。挑取血平皿上不生長(zhǎng)、巧克力平皿上生長(zhǎng)的無(wú)色透明或灰白色半透明、圓形隆起、光滑、邊緣整齊,濕潤(rùn)不溶血、露滴狀菌落、并有特殊閃光,直徑0.5~1 mm的可疑菌落涂片。革蘭染色呈陰性短小桿菌、球桿菌、少數(shù)呈細(xì)絲狀等多形態(tài)性。同時(shí)做衛(wèi)星試驗(yàn),將可疑菌落同時(shí)接種于血平皿、普通瓊脂(MH)平皿上,密涂劃線,再用金葡菌點(diǎn)種,兩點(diǎn)相距2 mm以上。35℃,5%~10%CO2培養(yǎng)箱內(nèi)培養(yǎng)24 h。血平皿衛(wèi)星試驗(yàn)陽(yáng)性、MH平皿衛(wèi)星試驗(yàn)陰性者,鑒定為流感嗜血桿菌。同時(shí)進(jìn)行V、X、X+V因子需求試驗(yàn),只能在V+X紙片周?chē)L(zhǎng)者,鑒定為流感嗜血桿菌。疑難菌株用法國(guó)生物梅里埃公司提供的NH卡最終鑒定。
(二)藥敏試驗(yàn) 藥敏試驗(yàn)采用紙片擴(kuò)散法。以HTM瓊脂為培養(yǎng)基,按美國(guó)CLSI標(biāo)準(zhǔn)判斷結(jié)果。
(三)β內(nèi)酰胺酶測(cè)定 用頭孢硝噻酚顯色反應(yīng)法測(cè)定β內(nèi)酰胺酶。
(四)質(zhì)控菌株 以金葡菌ATCC25923為陰性對(duì)照,流感嗜血桿菌ATCC49247為陽(yáng)性對(duì)照。
(五)統(tǒng)計(jì)學(xué)方法 采用WHONET5.6對(duì)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)學(xué)分析,P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
結(jié) 果
一、流感嗜血桿菌的分布
(一)年齡分布 2008年1月1日至2011年12月31日共收集到223株流感嗜血桿菌,其中分離自成人(≥18歲)的菌株為208株,占93.3%;分離自未成年人(<18歲)的菌株為15株,占6.7% 。
(二)科室分布 223株流感嗜血桿菌分離自門(mén)診101株(45.3%);分離自住院患者122株(54.7%),其中呼吸內(nèi)科17株(7.6%),其他內(nèi)科共90株(40.4%),外科10株(4.5%),兒科5株(2.2%)。
(三)標(biāo)本分布 223株流感嗜血桿菌分離自痰液194株(87.0%),咽拭子、支氣管肺泡灌洗液等呼吸道標(biāo)本28株(12.6%),血培養(yǎng)1株(0.4%)。
二、223株流感嗜血桿菌對(duì)各種抗菌藥物的耐藥性
本次檢測(cè)的流感嗜血桿對(duì)單環(huán)β內(nèi)酰胺類(氨曲南),喹諾酮類(環(huán)丙沙星、左氧氟沙星、莫西沙星),加酶抑制劑(阿莫西林-克拉維酸、氨芐西林-舒巴坦),大環(huán)內(nèi)酯類(阿奇霉素)抗菌藥物的敏感率都在90.0%以上;對(duì)第二代頭孢菌素(頭孢呋辛、頭孢克洛),第三代頭孢菌素(頭孢噻肟、頭孢曲松)的敏感率在80.0%以上,而對(duì)氨芐西林、甲氧芐啶-磺胺甲口惡唑和四環(huán)素的敏感率較低,分別為78.1%、41.9%和34.3%。
三、連續(xù)4年流感嗜血桿菌對(duì)抗菌藥物的敏感性分析
連續(xù)4年流感嗜血桿菌對(duì)大多數(shù)測(cè)試抗菌藥物的耐藥率差異無(wú)統(tǒng)計(jì)學(xué)意義,見(jiàn)表1。
四、連續(xù)4年分離自門(mén)診和住院患者的流感嗜血桿菌對(duì)抗菌藥物的敏感性分析
連續(xù)4年分離自門(mén)診和住院患者的流感嗜血桿菌對(duì)大多數(shù)測(cè)試抗菌藥物的耐藥率差異無(wú)統(tǒng)計(jì)學(xué)意義,見(jiàn)表2。
五、β內(nèi)酰胺酶檢測(cè)結(jié)果
(一)β內(nèi)酰胺酶陽(yáng)性菌 223株流感嗜血桿菌中產(chǎn)β內(nèi)酰胺酶陽(yáng)性氨芐西林耐藥菌株(BLPAR)為35株,產(chǎn)酶檢出率為15.7%。
(二)β內(nèi)酰胺酶陰性菌 β內(nèi)酰胺酶陰性氨芐西林耐藥菌株(BLNAR)為20株,占9.0%,見(jiàn)表3。
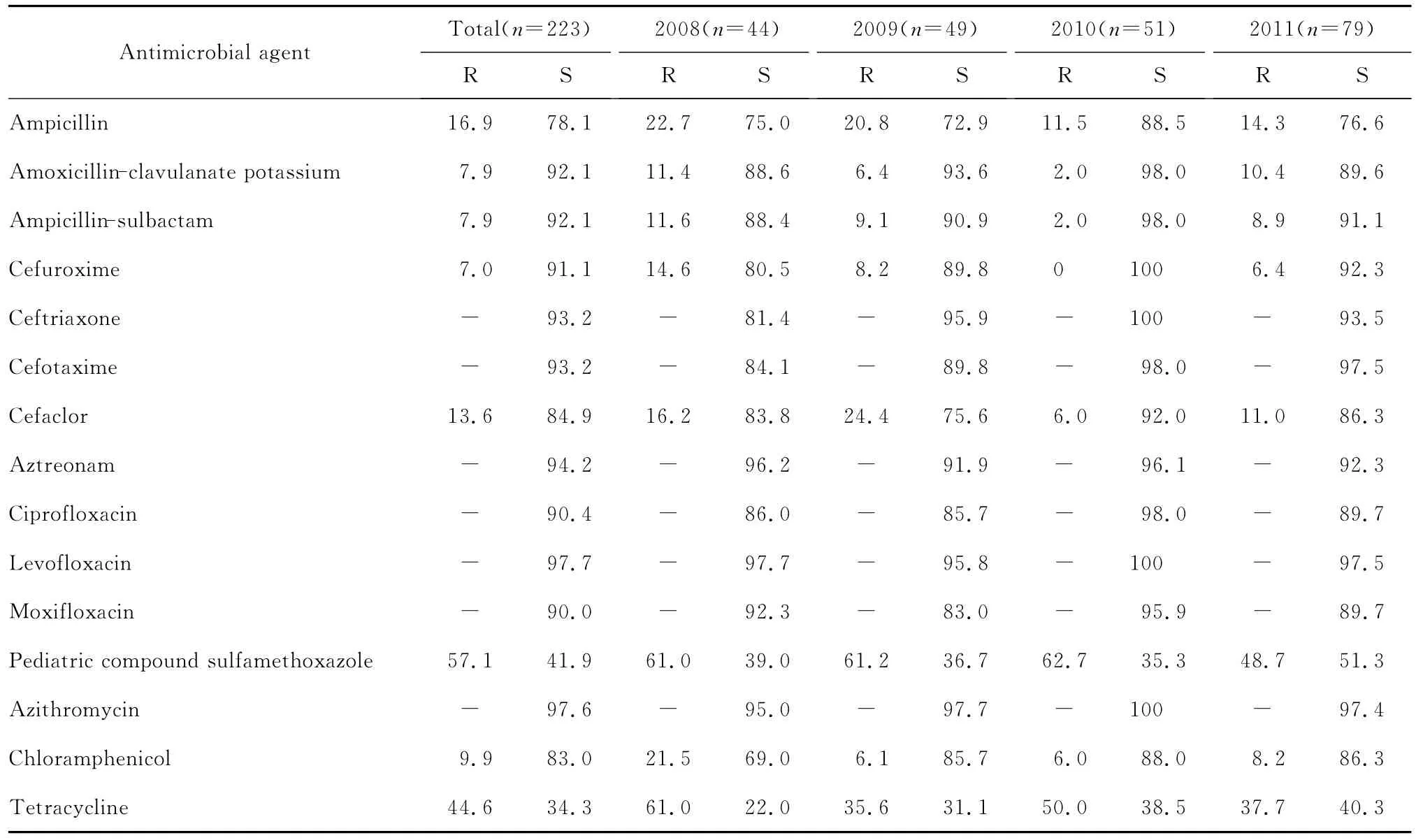
表1 2008—2011年流感嗜血桿菌對(duì)抗菌藥物的耐藥率和敏感率(%)。Table 1 The susceptibility of Haemophilus influenzae strains to various antimicrobial agents during 2008-2011(%)
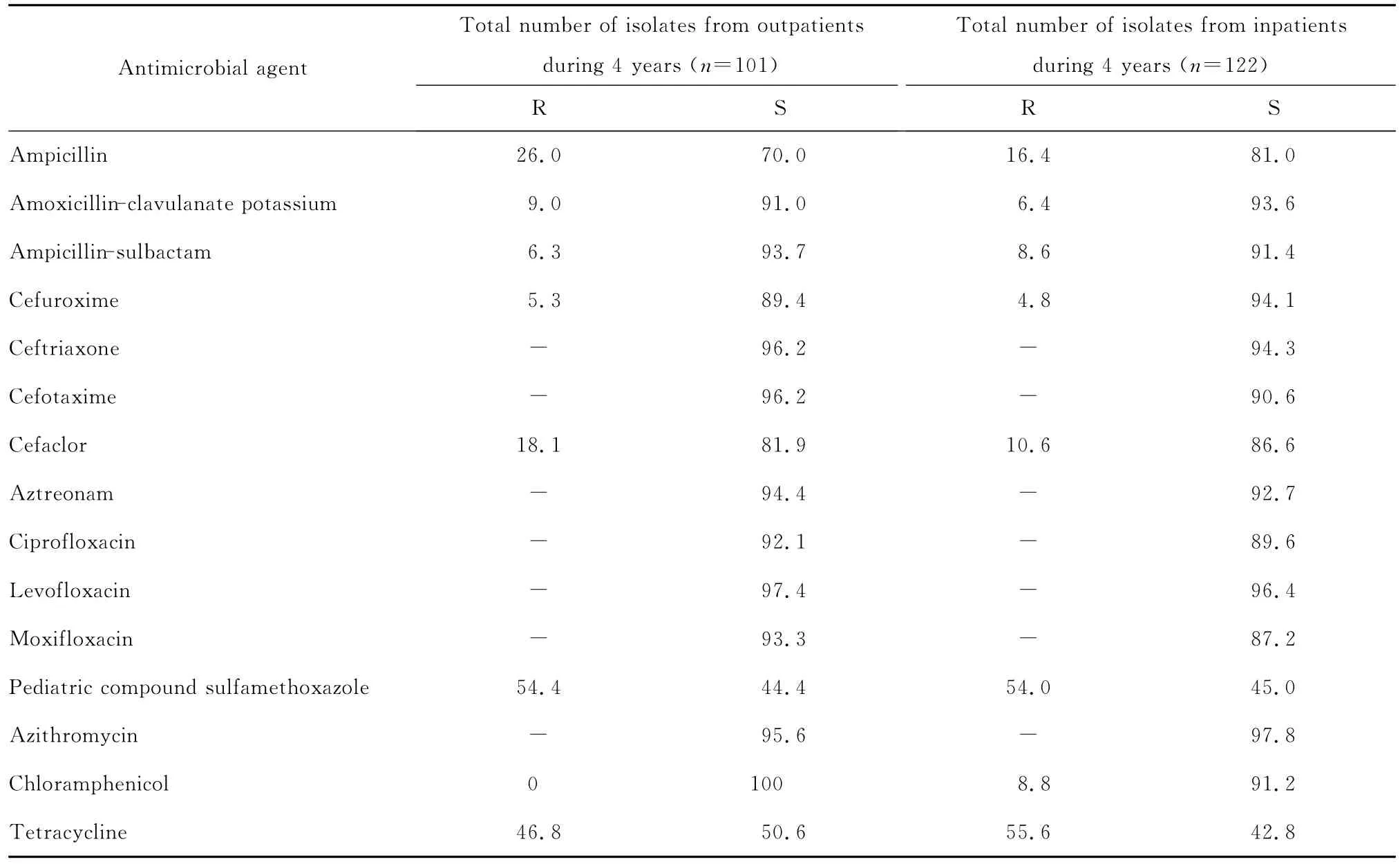
表2 4年間分離自門(mén)診和住院患者的流感嗜血桿菌對(duì)抗菌藥物的耐藥率和敏感率(%)。Table 2 The suscetibility of Haemophilus influenzae isolated from outpatients and inpatients to antimicrobial agents during 4 consecutive years(%)
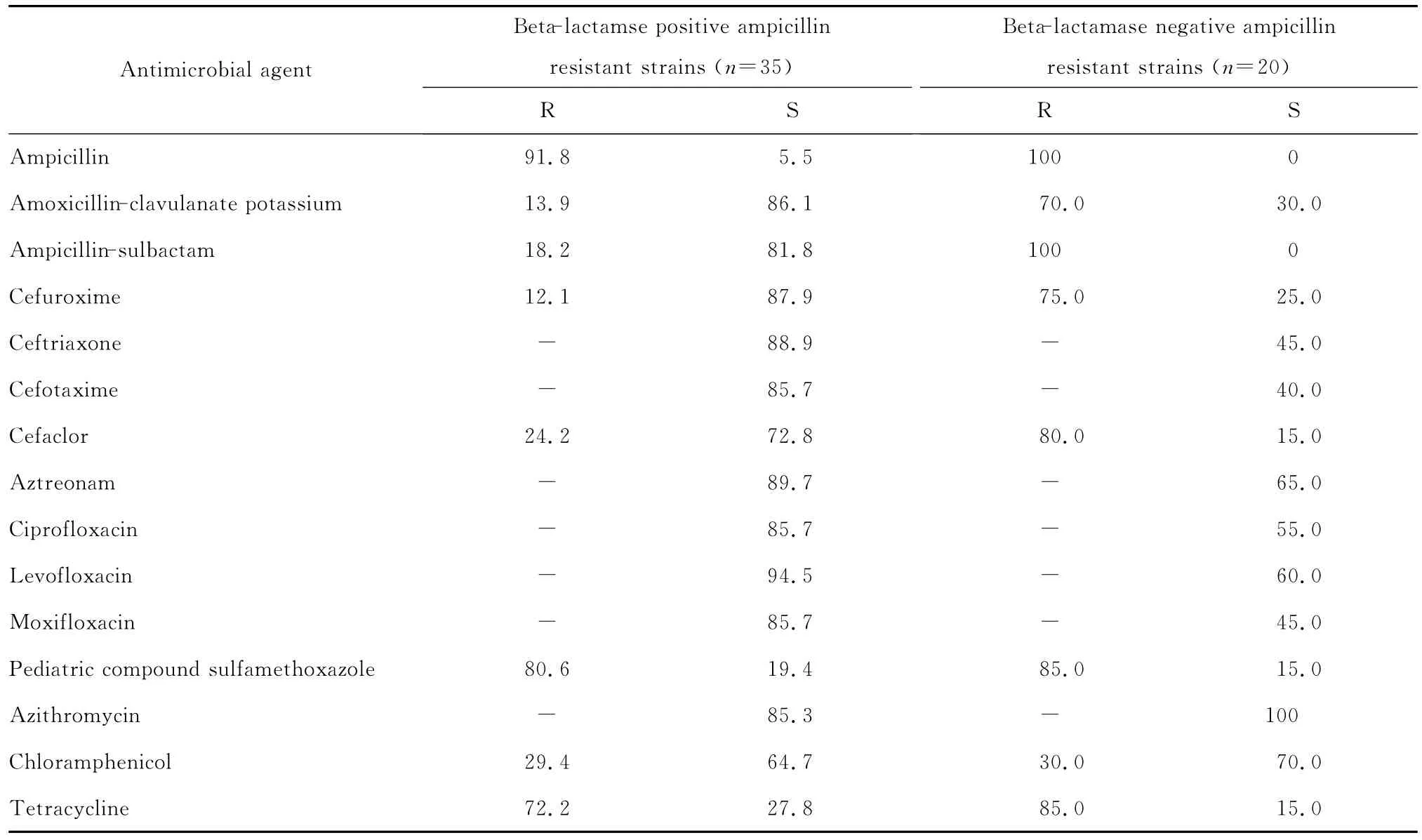
表3 4年間β內(nèi)酰胺酶陽(yáng)性和β內(nèi)酰胺酶陰性氨芐西林耐藥的流感嗜血桿菌對(duì)抗菌藥物的耐藥率和敏感率(%)Table 3 The susceptibility ofβ-lactamase-positive andβ-lactamase-negative ampicillin resistant Haemophilus influenzae strains to antimicrobial agents during 4 consecutive years(%)
討 論
本次統(tǒng)計(jì)中流感嗜血桿菌對(duì)氨芐西林不敏感率為21.9%,4年間差異無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05),略低于2010年中國(guó)CHINET細(xì)菌耐藥監(jiān)測(cè)數(shù)據(jù)(35.7%)[1]。對(duì)氨芐西林耐藥的流感嗜血桿菌在1973年被首次報(bào)道,產(chǎn)β內(nèi)酰胺酶是其主要耐藥機(jī)制。編碼β內(nèi)酰胺酶的基因主要在質(zhì)粒上,但也有些基因整合于細(xì)菌的染色體上,產(chǎn)生了2種β內(nèi)酰胺酶:TEM-1和 ROB-1,以 TEM-1比較常見(jiàn),而幾乎所有產(chǎn)ROB-1酶的菌株出現(xiàn)在北美[2]。產(chǎn)β內(nèi)酰胺酶的比率因地域分布而不同,如俄羅斯為4.2%,美國(guó)則為29.6%[3]。
β內(nèi)酰胺酶陰性氨芐西林耐藥,或者β內(nèi)酰胺酶陽(yáng)性,阿莫西林-克拉維酸耐藥是由于青霉素結(jié)合蛋白(PBP)的改變,二型和未分型的菌株是由PBP3介導(dǎo)的,PBP3由ftsI基因整合[4]。根據(jù)2012 CLSI的規(guī)定,BLNAR應(yīng)評(píng)價(jià)為對(duì)于這些菌株:阿莫西林-克拉維酸、氨芐西林-舒巴坦、頭孢克洛、頭孢他美、頭孢尼西、頭孢丙烯、頭孢呋辛、氯碳頭孢,頭孢孟多以及哌拉西林-他唑巴坦均視為耐藥,即使一些BLNAR株體外試驗(yàn)顯示對(duì)這些藥物敏感也應(yīng)視為耐藥。
超廣譜β內(nèi)酰胺酶(ESBL),是由TEM-1變異而來(lái)的,但是在流感嗜血桿菌中還未發(fā)現(xiàn),盡管已經(jīng)在克隆株上有表達(dá)[5],現(xiàn)已報(bào)道由2例南非患者體內(nèi)分離的副流感嗜血桿菌是產(chǎn)ESBL的[6]。
流感嗜血桿菌對(duì)大環(huán)內(nèi)酯類抗生素的耐藥率在過(guò)去30年里一直沒(méi)有改變,盡管已經(jīng)發(fā)現(xiàn)了少量高耐藥株[7-8]。
流感嗜血桿菌對(duì)喹諾酮類抗菌藥物耐藥非常罕見(jiàn),廣譜喹諾酮類抗菌藥物通過(guò)干擾DNA復(fù)制而發(fā)揮抗菌作用,編碼DNA解螺旋酶或者拓?fù)洚悩?gòu)酶Ⅳ的耐藥決定區(qū)的改變導(dǎo)致流感嗜血桿菌對(duì)喹諾酮藥物耐藥。喹諾酮類藥物由于其不良反應(yīng)目前不推薦給兒童使用。但是一項(xiàng)監(jiān)測(cè)表明,一些喹諾酮類藥物MIC值在增加,并且在一個(gè)長(zhǎng)期護(hù)理的設(shè)備上發(fā)現(xiàn)了1株高耐藥的克隆株[9]。相反,其對(duì)甲氧芐啶-磺胺甲口惡唑的耐藥率在過(guò)去的20年間不斷上升,2003年比利時(shí)為8.5%,肯尼亞為55.2%[3],2010年我國(guó)為64.5%[1],我院為57.1%。
同樣,流感嗜血桿菌對(duì)四環(huán)素的耐藥率較高,四環(huán)素類抗菌作用是藥物結(jié)合到細(xì)菌核糖體的30S并阻止tRNA接合到A和P位點(diǎn)上[10]。對(duì)四環(huán)素的耐藥性是由于流感嗜血桿菌的細(xì)胞膜上的tet(B)基因產(chǎn)生的外排機(jī)制,這種基因位于接合質(zhì)粒上[11-12]。tet(B)基因編碼的外排蛋白可以對(duì)四環(huán)素和米諾環(huán)素耐藥。由氨芐西林、卡那霉素的耐藥基因介導(dǎo)的質(zhì)粒經(jīng)常轉(zhuǎn)移到四環(huán)素上,導(dǎo)致其耐藥,這一現(xiàn)象已在比利時(shí)、西班牙和古巴分離的B型流感嗜血桿菌中報(bào)道[13-14]。流感嗜血桿菌對(duì)四環(huán)素和氯霉素的耐藥性增高,與質(zhì)粒介導(dǎo)的氨芐西林、氯霉素、四環(huán)素、卡那霉素耐藥基因有關(guān)。根據(jù)2012 CLSI標(biāo)準(zhǔn),如果細(xì)菌對(duì)四環(huán)素耐藥,同時(shí)也對(duì)強(qiáng)力霉素和米諾環(huán)素耐藥。
從我院分離的流感嗜血桿菌4年來(lái)對(duì)頭孢噻肟、氯霉素和四環(huán)素的耐藥率有所降低,說(shuō)明醫(yī)師應(yīng)用抗菌藥物合理。但由于我國(guó)流感嗜血桿菌分離率較低,尚未對(duì)該菌造成的感染及其耐藥性引起重視,如果不合理使用抗菌藥物的現(xiàn)象持續(xù)下去,將會(huì)有高耐藥株出現(xiàn)[15]。
[1] 朱德妹,汪復(fù),胡付品,等.2010年中國(guó)CHINET細(xì)菌耐藥性監(jiān)測(cè)[J].中國(guó)感染與化療雜志,2011,9(5):321-329.
[2] Farrell DJ,Morrissey I,Bakker S,et al.Global distribution of TEM-1 and ROB-1 beta-lactamases in Haemophilus influenzae [J].J Antimicrob Chemother,2005,56(4):773.
[3] Jacobs MR,F(xiàn)elmingham D,Appelbaum PC,et al.The Alexander Project 1998-2000:susceptibility of pathogens isolated from community-acquired respiratory tract infection to commonly used antimicrobial agents[J].J Antimicrob Chemother,2003,52(1):229-246.
[4] Tristram S,Jacobs MR,Appelbaum PC.Antimicrobial resistance Haemophilus influenzae [J].Clin Microbiol Rev,2007,20(2):368-389.
[5] Bozdogan B,Tristram S,Appelbaum PC.Combination of altered PBPs and expression of cloned extended-spectrum betalactamases confers cefotaxime resistance in Haemophilus influenzae[J].J Antimicrob Chemother,2006,57(4):747-749.
[6] Pitout M,MacDonald K,Musgrave H,et al.Characterization of extended spectrum beta-lactamase(ESBL)activity in Haemophilus influenzae[M]//Program and Abstracts of the 42nd Interscience Conference on Antimicrobials and Chemotherapy,San Diego,CA.American Society for Microbiology,Washington,DC,USA.2002:96.
[7] Kim IS,Ki CS,Kim S,et al.Diversity of ampicillin resistance genes and antimicrobial susceptibility patterns in Haemophilus influenzae strains isolated in Korea[J].Antimicrob Agents Chemother,2007,51(2):453-460.
[8] Garcia-Cobos S,Campos J,Lazaro E,et al.Ampicillin-resistant non-beta-lactamase-producing Haemophilus influenzae in Spain:recent emergence of clonal isolates with increased resistance to cefotaxime and cefixime[J].Antimicrob Agents Chemother,2007,51(7):2564-2573.
[9] Nazir J,Urban C,Mariano N,et al.Quinolone-resistant Haemophilus influenzae in a long-term care facility:clinical and molecular epidemiology[J].Clin Infect Dis,2004,38(11):1564-1569.
[10] Chopra I,Hawkey PM,Hinton M.Tetracyclines,molecular and clinical aspects[J].J Antimicrob Chemother,1992,29(3):245-277.
[11] Marshall B,Roberts M,Smith A,et al.Homogeneity of transferable tetracycline-resistance determinants in Haemophilus species[J].J Infect Dis,1984,149(6):1028-1029.
[12] Chopra I,Roberts M.Tetracycline antibiotics:mode of action,applications,molecular biology,and epidemiology of bacterial resistance[J].Microbiol Mol Biol Rev,2001,65(2):232-260.
[13] Campos J,Chanyangam M,deGroot R,et al.Genetic relatedness of antibiotic resistance determinants in multiply resistant Hemophilus influenzae[J].J Infect Dis,1989,160(5):810-817.
[14] Levy J,Verhaegen G,De Mol P,et al.Molecular characterization of resistance plasmids in epidemiologi-cally unrelated strains of multiresistant Haemophilus influenzae [J].J Infect Dis,1993,168(1):177-187.
[15] 楚文瑛.臨床微生物實(shí)驗(yàn)室需重視嗜血桿菌的分離培養(yǎng)[J].國(guó)際檢驗(yàn)醫(yī)學(xué)雜志,2011,32(18):2160-2162.
猜你喜歡
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
現(xiàn)代臨床醫(yī)學(xué)(2022年3期)2022-06-06 07:59:40
昆明醫(yī)科大學(xué)學(xué)報(bào)(2022年1期)2022-02-28 07:43:40
天津醫(yī)科大學(xué)學(xué)報(bào)(2021年3期)2021-07-21 09:04:02
科學(xué)大眾(2020年12期)2020-08-13 03:22:22
云南醫(yī)藥(2019年3期)2019-07-25 07:25:10
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2016年1期)2016-11-12 13:19:40
國(guó)外醫(yī)藥(抗生素分冊(cè))(2016年6期)2016-07-10 11:34:45
中國(guó)衛(wèi)生標(biāo)準(zhǔn)管理(2015年14期)2016-01-15 02:58:37
中國(guó)當(dāng)代醫(yī)藥(2015年17期)2015-03-01 02:03:58
