水稻萌發期耐Cu2+ 脅迫的QTL 定位*
2013-11-25 10:02:38饒玉春楊窯龍李曉靜馬伯軍曾大力
浙江師范大學學報(自然科學版) 2013年2期
饒玉春,楊窯龍,李曉靜,馬伯軍,曾大力
(1.浙江師范大學 化學與生命科學學院,浙江 金華 321004;2.中國水稻研究所 水稻生物學國家重點實驗室,浙江 杭州 310006)
20 世紀以來,隨著工農業、采礦、冶煉和制造等產業的快速發展,農用化學品的大量使用及城市污水的排放,植物受銅毒害的報道和研究逐漸增多[1-3].銅是水稻生長必須的微量元素,其對水稻的影響間接且較為復雜[4].過量的銅會導致水稻的銅毒害,阻礙水稻生長[5-6],甚至影響水稻產量[7-8].另外,銅在水稻體內大量累積,通過食物鏈傳遞,進而會威脅人類的健康[9].
銅傷害植物首先是對根系,傷害最嚴重的也是根系[10-11].銅對水稻幼苗根的生長有顯著的抑制作用,當銅質量濃度為50 mg/L 時,水稻根生長的抑制率達54%;銅質量濃度為600 mg/L 時,抑制率高達99%,根的長度與銅濃度呈顯著的負相關[12].水稻受銅毒害后通常表現為葉子細弱,葉片失綠發黃或紫紅乃至枯萎,返青延遲,心葉逐漸扭曲,植株瘦小,分蘗明顯減少,抽穗和成熟期延遲,且隨著污染程度加重,毒害癥狀相應加重[13].銅毒害主要是通過影響其他營養元素的吸收造成的.徐加寬等[14]研究發現,在土壤銅脅迫條件下,水稻吸磷能力明顯下降,影響植株的光合作用;邵登輝等[15]發現銅脅迫能顯著影響水稻對氮的吸收;Hiroki 等[16]在煙草細胞的研究中發現,銅能使質膜鈣通道的鈣通量增大,從而增加細胞溶質中的游離鈣.另外,銅脅迫對水稻光合作用、體內激素含量、細胞結構及質膜透性等均有影響[17].
目前,從分子和基因水平研究植物抗重金屬污染已引起國內外學者的高度重視[18],但在水稻上分離出來的耐銅脅迫基因或主效數量性狀基因座(QTL)少有報道.本研究嘗試應用一對典型的秈粳交(春江06/臺中本地1 號)F1 經花藥培養產生的雙單倍體(DH)群體及其構建的分子連鎖圖譜,系統考察了該DH 群體及其雙親耐銅脅迫的情況,進行了QTL 分析,并計算了QTL 對水稻耐銅性的貢獻率和加性效應,同時還進行了復合QTL 模型分析,以探討水稻耐銅脅迫的遺傳基礎.
1 材料與方法
1.1 遺傳群體的構建
本研究利用秈稻品種臺中本地1 號(TN1)和粳稻品種春江06(CJ06)為親本,對F1 代進行花藥離體培養,再經自然加倍或秋水仙素處理,共獲得純合二倍體(DH系)120個株系,組成DH 群體.
1.2 銅脅迫濃度的確定
為確定合適銅脅迫處理的濃度,對DH 群體親本CJ06 和TN1 進行了濃度梯度處理,設0,50,100,200,400,800,1 600 μmol/L 共7 個濃度的CuSO4溶液做梯度處理,每個處理3 次重復.考察雙親在各性狀上的差異程度,選取合適的濃度對DH 群體進行處理,用滅菌水作對照.
1.3 DH 群體耐銅脅迫的考察
種子萌發參照Tomar 等[19]的方法.取各株系的水稻種子500 粒,經0.50%次氯酸鈉表面消毒20 min,再用去離子水沖洗數次,28 ℃暗中浸種催芽48 h,挑選露白一致的種子播于墊有2 層濾紙的培養皿內培養,共6 皿,培養皿直徑約9 cm,其中3 皿用CuSO4溶液培養,另3 皿滅菌水培養.第l 天加入8 mL CuSO4溶液或滅菌水,每隔24 h換一次,連續4 d 后結束.以芽長超過1/2 種子長為發芽標準,以根長超過1/2 種子長為發根標準,記錄發芽數、發根數,計算發芽率、發根率;隨機取20 粒種子測定根長、芽長;并對每皿的總根和芽稱質量,以CuSO4溶液條件下對滅菌水條件下的百分率表示耐抑制程度.
1.4 連鎖圖譜的構建和QTL 定位
利用微衛星DNA(SSR)和序列標志位點(STS)標記,構建了該DH 群體的高密度分子標記連鎖圖譜,從中選取178 個均勻分布在12 條染色體上的標記構建連鎖圖譜,該連鎖圖譜總共覆蓋1 674.80 cM,平均圖距為9.44 cM,它們均適合QTL 區 間 作 圖[20-21].采 用 作 圖 分 析 軟 件Mapmaker/Exp3.0b,用QTL 分析軟件mapmaker/QTL1.1B 對控制水稻種子萌發時期的相關性狀進行QTL 分析.以檢出限(LOD)為2.50 作為閾值判斷QTL 的存在與否,若標記區間檢出限>2.50,則認為該區間檢出限最高處所對應的位點為一個QTL.同時,計算出每個QTL 對各性狀表型方差的貢獻率和加性效應,QTL 的命名原則遵循McCouch 等[22-23]的方法.
2 結果與分析
2.1 銅脅迫濃度的確定
本實驗采用0,50,100,200,400,800,1 600 μmol/L 共7 個濃度的CuSO4溶液做梯度處理,對DH 群體雙親CJ06 和TN1 進行脅迫處理,并通過統計方法方差分析.結果顯示:雙親在100 μmol/L CuSO4溶液的處理下差異顯著(見表1),此時TN1 的發芽率和發根率都在97%以上,幾乎不受銅脅迫的影響;而CJ06 在該濃度下的發芽率為83%,發根率僅為61%,受脅迫的表現明顯;同時,雙親的平均芽長、平均根長、芽質量和根質量差異度很明顯.當CuSO4溶液濃度超過200 μmol/L 以后,雙親各性狀受脅迫的影響都很大,銅濃度越高,受脅迫的表型越明顯,但雙親之間的差異難以區分.
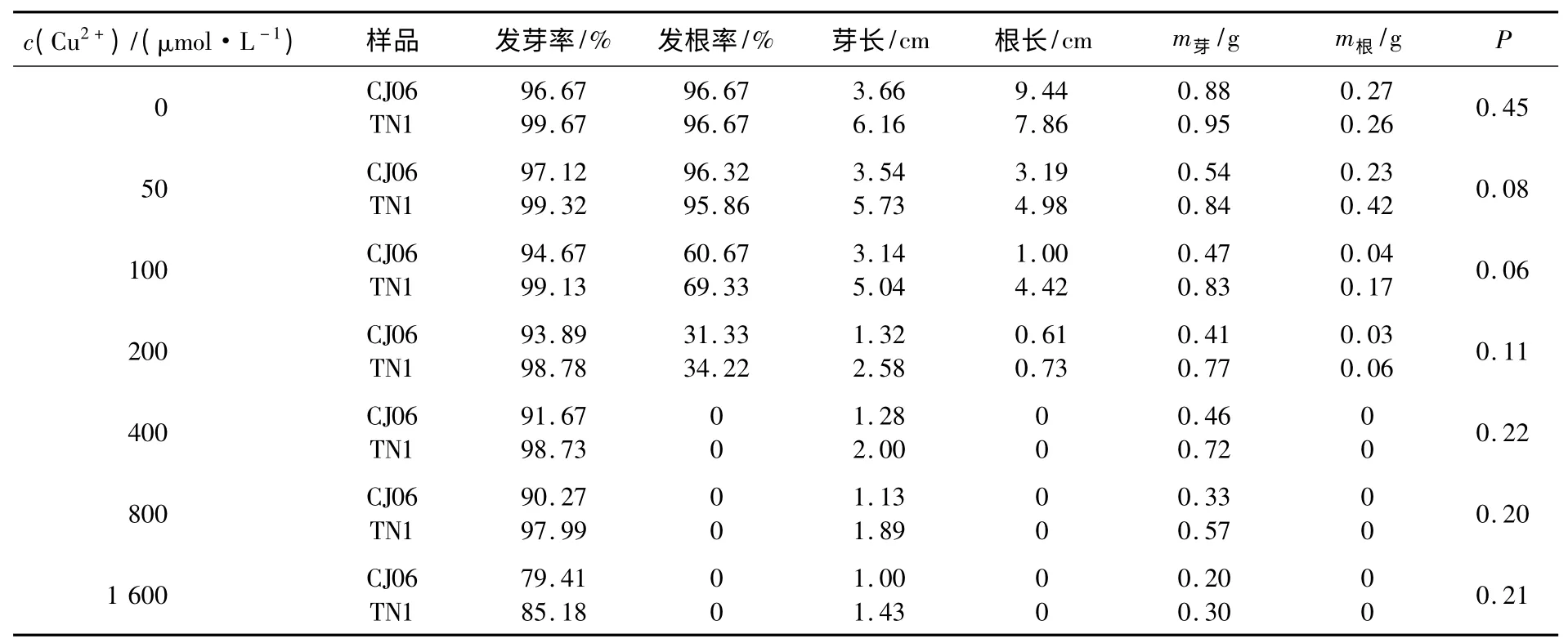
表1 不同濃度銅處理下雙親萌發期的性狀表現
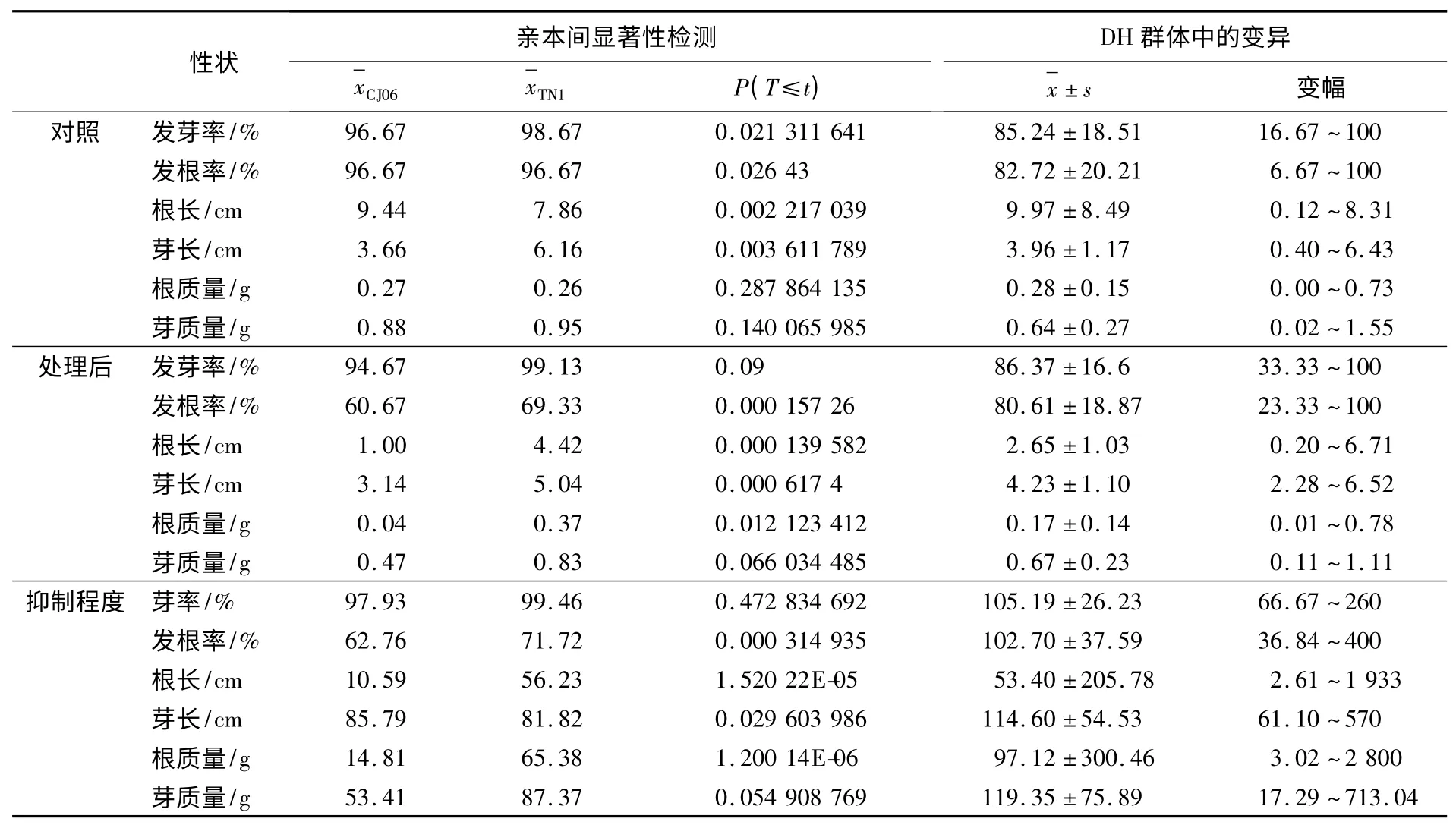
表2 銅脅迫下水稻萌發期相關性狀在親本間的顯著性檢測及在DH 群體中的表現
2.2 雙親和群體在各性狀上的表現
用100 μmol/L CuSO4溶液處理DH 群體中的后代各株系,并考察了萌發期相關性狀的數據,同時用滅菌水處理作對照.結果發現,親本CJ06在發芽率、發根率、平均芽長、根質量和芽質量等性狀上對銅脅迫比TN1 敏感(見表2),尤其發根率和平均芽長最為明顯.且120 個DH 株系對銅脅迫的耐抑制表現為連續正態分布,且有一定數量的超親類型存在,符合QTL 區間作圖的要求.
2.3 性狀間的相關性分析
表3 是各性狀的相關性分析結果,表明:水稻發芽率與發根率、根質量與芽質量之間均呈現顯著的正相關,尤其是水稻種子的發芽率與發根率之間,在處理前后其相關系數都達到了0.90,而根質量與芽質量的相關系數也達到了0.50 附近.由此可以說明,水稻發芽期間,根與芽之間有著密切的相關性.在這些性狀中,根長與其他的性狀并不存在相關性,說明根長與其他性狀關系并不密切.同時,相關性分析也表明:未用CuSO4溶液處理的水稻種子,其平均芽長與平均根長并無顯著相關性;而用100 μmol/L CuSO4溶液處理后,其平均芽長與平均根長呈現顯著正相關.這可能是由于用100 μmol/L CuSO4溶液處理對水稻種子的平均芽長與平均根長影響較大.
2.4 QTL 定位
利用該DH 群體對水稻萌發期相關性狀進行了QTL 分析,有11 個性狀檢測到了相關的QTL(見圖1 和表4),共有30 個QTLs,它們分布于除8,11 和12 染色體外的其他9 條染色體上.其中,用滅菌水處理種子后檢測到萌發期相關性狀的QTL 共22 個,用100 μmol/L CuSO4溶液處理后檢測到萌發期相關性狀的QTL 共6 個,而抑制程度共檢測到1 個QTL.
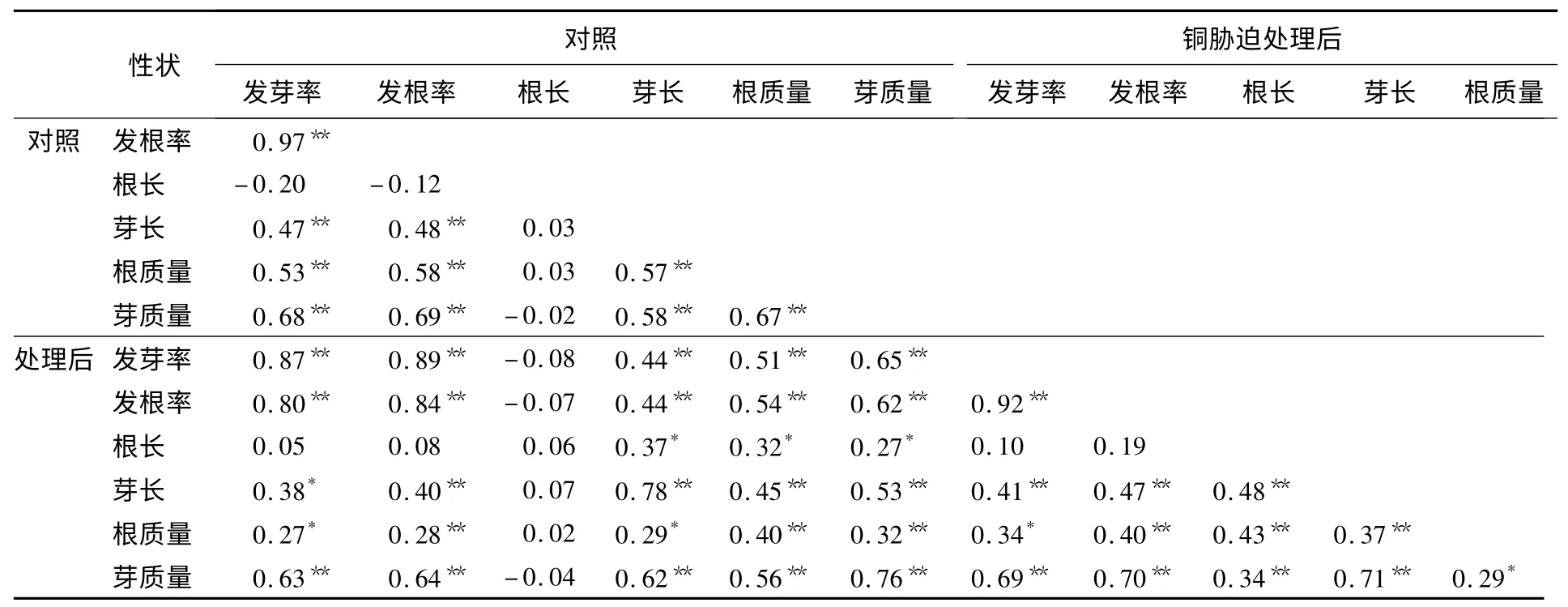
表3 DH 群體中水稻萌發期各性狀的相關性分析
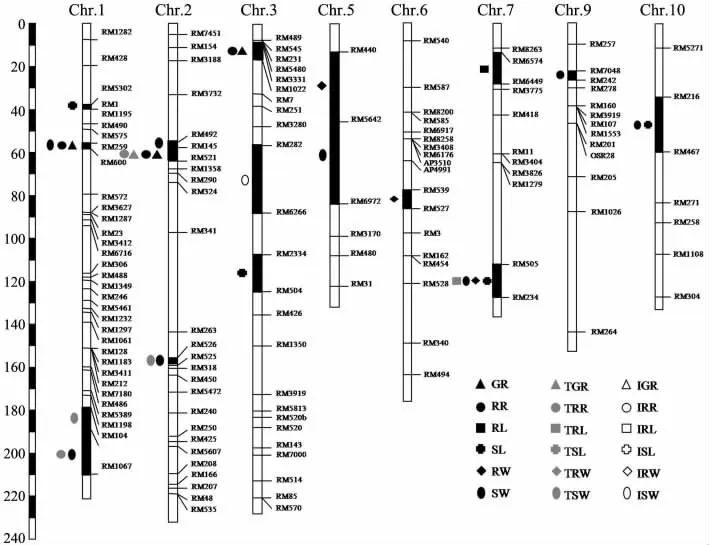
圖1 CJ06/TN1 DH 群體水稻萌發期相關性狀的QTL 定位圖譜
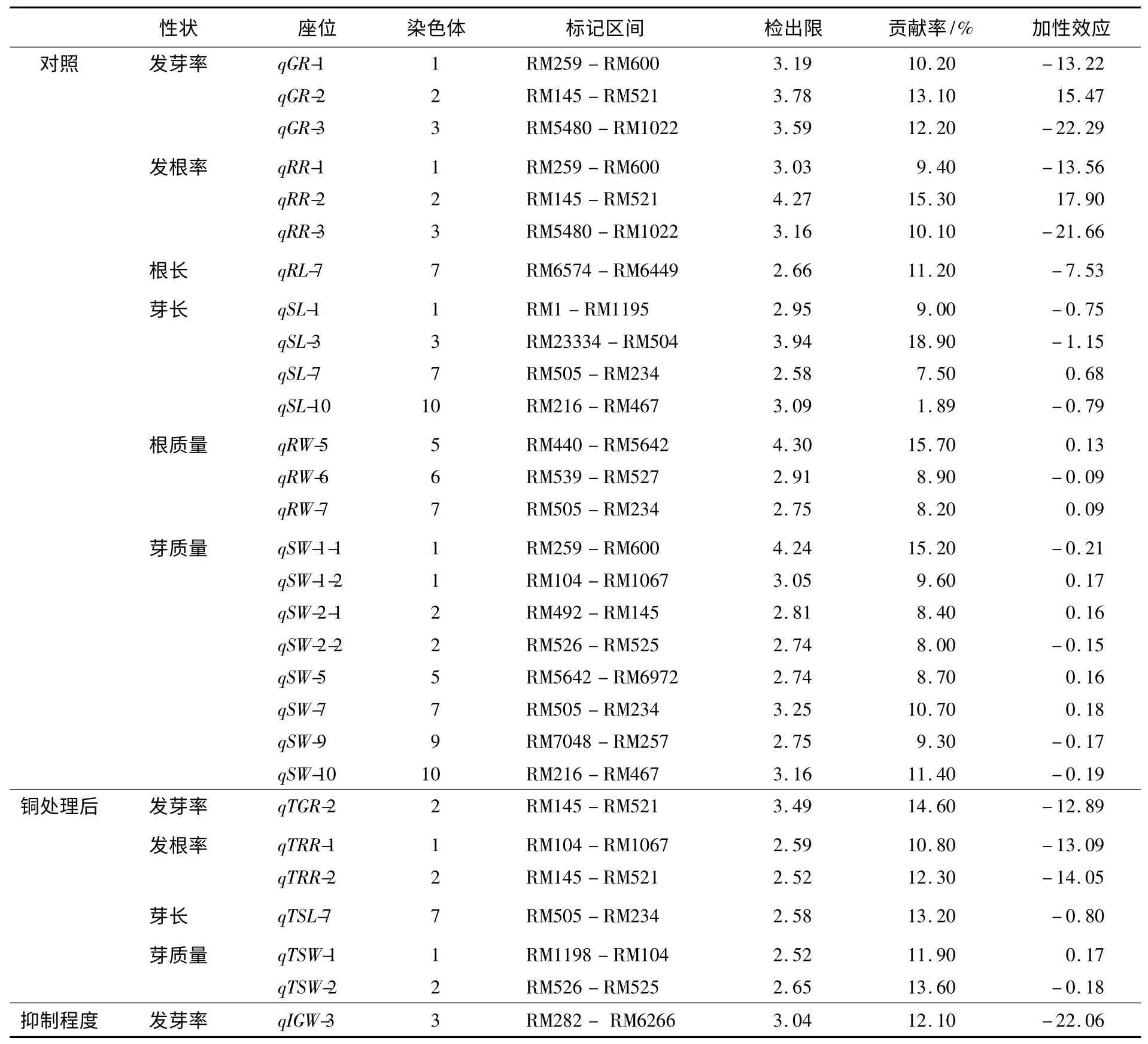
表4 DH 群體中水稻萌發期相關性狀的QTL 定位
在用滅菌水處理種子后檢測到的22 個QTLs中,與發芽率、發根率、根長、芽長、根質量和芽質量相關的QTL 分別有3,3,1,4,3 和8 個,它們分別位于除4,11 和12 染色體之外的所有染色體上,其檢出限介于2.52~4.30,可以解釋性狀變異的8.00%~15.70%.在這些QTL 中,與發芽率相關的QTL 有3 個,位于1,2 和3 染色體上,與發根率檢測到的QTL 區間重合,說明水稻種子發芽與發根是緊密相關的,受同一類QTL 控制.在7 染色體上檢測到1 個與根長相關的QTL.而與芽長相關的QTL 有4 個,分別位于1,3,7 和10 染色體上,其中有3 個增效等位基因來自于TN1,1 個來自于CJ06,這可以間接解釋TN1 的芽長比CJ06長的原因.另外,檢測到3 個與根質量有關的QTL,其中在5 染色體上有一個檢出限達到了4.30,可解釋性狀變異的15.70%.檢測到與芽質量有關的QTL 有8 個,其檢出限介于2.74~4.24之間,可解釋性狀變異的8.00%~15.20%,其QTL 大部分都和發芽率、發根率、根長、芽長、根質量的QTL 區間重合,這也充分說明水稻萌發期各類性狀的調控都存在著相互聯系.
用100 μmol/L CuSO4溶液處理種子后檢測到6 個萌發期相關性狀的QTL,分布在1,2 和7染色體上,其中發芽率與發根率具有重合的QTL區間,進一步說明了發芽率與發根率存在著密切的關系.但沒有檢測到根長與根質量的相關QTL.而芽長與芽質量相關性狀,不管是經過100 μmol/L CuSO4溶液處理后,還是沒有經過CuSO4溶液處理,在2 染色體上的RM526-RM525 區間與7 染色體上的RM505-RM234 區間都檢測到了相關QTL,說明2 個QTL 區間對芽長與芽質量的調控是密切相關的,且對銅脅迫有一定的耐性.
對水稻萌發期100 μmol/L CuSO4溶液處理后各性狀的抑制程度進行QTL 分析,發根率、根長、芽長、根質量和芽質量均沒有檢測到相關的QTL,只有發芽率的抑制程度在3 染色體上RM282-RM6266 區間檢測到1 個QTL,該增效基因來自于TN1,可解釋表型變異的12.10%.
3 討論
目前,對于水稻萌發期耐銅脅迫的研究相對較少,也沒有明確的耐銅脅迫基因的相關報道.應用銅脅迫實驗進行群體的耐銅性遺傳研究,選擇適當的培養方式和合適的銅脅迫濃度是實驗成敗的關鍵[24].本實驗利用不同濃度的硫酸銅溶液對DH 群體的雙親進行處理,發現在100 μmol/L CuSO4溶液處理下,雙親萌發期的各種性狀差異較為顯著;之后,用該濃度銅脅迫處理整個群體,同時用滅菌水處理作為對照,進行QTL 作圖;最后,將100 μmol/L CuSO4溶液處理下各種性狀的值和用滅菌水處理下對應性狀的值的比值作為一種抑制程度的指標進行QTL 分析.在檢測到的QTLs 中,大部分是在滅菌水處理下檢測到的,用100 μmol/L CuSO4溶液處理后共檢測到萌發期相關性狀的6 個QTLs,而抑制程度共檢測到1 個QTL,這可能是由于經過銅處理后普遍抑制了控制各性狀的QTLs 的作用,且其抑制程度相當,消除了個別QTL 的主效作用.其中在1 染色體上RM104-RM1067 區間和2 染色體RM526-RM525區間檢測到了芽質量的QTL 與滅菌水處理下檢測到的芽質量QTL 區間一致,說明在這兩區間內存在著控制芽質量的主效QTL,不受CuSO4溶液處理的影響,證明這兩位點對銅脅迫有很好的耐性.同樣,發芽率和發根率2 個性狀在CuSO4溶液處理前后都能在2 染色體的RM145-RM521 區間上檢測到相應的QTL,證明這個區間內也存在著對銅脅迫有很好耐性的位點.
對比其他學者的研究結果,有些耐銅脅迫的區間與本實驗檢測到的區間有重疊.姚盟成等[25]利用栽培稻優良品種“特青”與普通野生稻“元江普野”構建的DH 群體139 個家系構建連鎖圖譜,用硫酸銅(150 mg/L)脅迫處理2 葉期的水稻苗,在水稻2 染色體RM318 標記處檢測到了耐銅脅迫的QTL,與本實驗檢測到的耐銅脅迫的區間類似.同時,藏金萍[26]以伊朗粳稻品種Binam 為供體,以秈稻品種特青為輪回親本,構建了BC2F8定位群體,并在水稻3 染色體上定位到了耐鹽的QTL(QRkc3),該區間在本實驗檢測到的耐銅脅迫的區間附近,說明在水稻2 染色體RM526-RM525 區間周圍存在一個主效的位點,對高濃度的金屬脅迫有一定的耐性,由于所用的親本品種和考察的性狀不一致而導致了區間略有偏移.另外,在3 染色體上檢測到關于發芽率抑制程度的QTL 區間RM282-RM6266 以前未曾見報到;CuSO4溶液處理下,在7 染色體上檢測到關于芽長的QTL 區間RM505-RM234 與前人的研究區間也不重疊.
整體上看,CuSO4溶液處理后檢測到一些耐銅的QTL,但QTL 與環境互作是影響數量性狀表達的重要因素[27],要從遺傳上改良水稻品種的耐銅性并非易事,通過常規有性雜交技術難于將這些分散的有利基因集中起來.下一步我們將通過分子標記輔助選育技術,構建關于這幾個耐銅脅迫位點的單片段置換系,以期為選育抗性品種提供耐銅脅迫的基因.
[1]Holmgren G G S,Meyer M W,Cahney R L,et al.Cadmium,lead,zinc,copper and nickel in agricultural soils of the United States of America[J].Journal of Environmental Quality,1993,22(2):335-348.
[2]張秀梅,唐以劍,章申.白洋淀地區土壤-植物系統污染物含量與變化規律研究[J].地理科學進展,1997,16(2):62-69.
[3]Xu Jiakuan,Yang Lianxin,Wang Ziqiang,et al.Toxicity of copper on rice growth and accumulation of copper in rice grain in copper contaminated soil[J].Chemosphere,2006,62(4):602-607.
[4]李海華,劉建武,李樹人,等.土壤-植物系統中重金屬污染及作物富集研究進展[J].河南農業大學學報,2000,34(1):30-34.
[5]Ouzounidou G.Copper-induced changes on growth,methal content and photosynthetic function of Alyssum montanum L.plants[J].Environment Experiment Botany,1994,34(2):165-172.
[6]Nagalakshmi N,Prasad M N V.Copper-induced oxidative stress in Scenedesmus bijugatus:protective role of free radical scavenger[J].Bull Environ Contam Toxical,1998,61(5):623-628.
[7]江行玉,趙可夫.植物重金屬傷害及其抗性機理[J].應用與環境生物學報,2001,7(1):92-99.
[8]袁玲花,徐加寬,顏士敏,等.土壤銅脅迫對不同秈型水稻品種產量和品質的影響[J].農業環境科學學報,2008,27(2):435-441.
[9]丁園,宗良綱.不同土壤重金屬復合污染有效態離子沖量表征[J].環境污染與防治,2003,25(3):173-175.
[10]宋玉芳,許華夏,任麗萍,等.重金屬對西紅柿種子發芽與根伸長的抑制效應[J].中國環境科學,2001,21(5):390-394.
[11]羅亞平,劉杰,蔡湘文,等.銅對水稻種子萌發和生長的影響[J].廣西農業科學,2005,36(4):316-318.
[12]胡筑兵,陳亞華,王桂萍,等.銅脅迫對玉米幼苗生長、葉綠素熒光參數和抗氧化酶活性的影響[J].植物學通報,2006,23(2):129-137.
[13]崔德杰,張玉龍.土壤重金屬污染現狀與修復技術研究進展[J].土壤通報,2004,35(3):366-370.
[14]徐加寬,楊連新,王志強,等.土壤銅脅迫對水稻磷素吸收利用及產量的影響[J].江蘇農業科學,2008,35(2):213-217.
[15]邵登輝,儲玲.銅脅迫下氮肥對水稻幼苗生長的影響[J].南京曉莊學院學報,2008(3):59-63.
[16]Hiroki I,Tomoko K,Hiroshi K,et al.Copper elicits an increase in cytosolic free calcium in cultured tobacco cells[J].Plant Physiology and Biochemistry,2005,43(12):1089-1094.
[17]王狄,李鋒民,熊治廷,等.銅的植物毒性與植物蓄積的關系[J].土壤與環境,2000,9(2):146-148.
[18]徐加寬,楊連新,王余龍,等.水稻對重金屬元素的吸收與分配機理的研究進展[J].植物學通報,2005,22(5):614-622.
[19]Tomar R P S,Singh B P,Chaudhary B S.Effect of modes application of growth regulators on wheat[J].Indian J Agr Res,1977,5:41-44.
[20]Zeng Dali,Hu Jiang,Dong Guojun,et al.Quantitative trait loci mapping of flaf-leaf ligule length in rice and alignment with ZmLG1 gene[J].JIPB,2009,51(4):360-366.
[21]Rao Yuchun,Dong Guojun,Zeng Dali,et al.Genetic analysis of leaffolder resistance in rice[J].Genet Genomics,2010,37(5):325-331.
[22]Khush G S.Prospects of and approaches to increasing the genetic yield potential of rice[M]//Evenson R E,Herdt R W,Hossain M.Rice research in Asia:progress and priorities.Wallingford:CAB International,1996:57-71.
[23]McCouch S R,Cho Y G,Yano M,et al.Report on QTL nomenclature[J].Rice Genet Newsl,1997,14(11):11-131.
[24]沈圣泉,莊杰云,舒慶堯,等.水稻苗期耐高Cu2+脅迫的QTL 定位和上位性分析[J].植物營養與肥料學報,2006,12(3):352-357.
[25]姚盟成,張文會,張曉杰,等.水稻苗期耐Cu2+脅迫QTL 遺傳分析[J].廣西農業科學,2009,40(9):1119-1123.
[26]藏金萍.水稻抗旱、耐鹽QTL 表達的遺傳背景效應及抗旱、耐鹽的遺傳重疊研究[D].北京:中國農業科學院,2008:17-18.
[27]樊葉楊,莊杰云,李強,等.水稻株高QTL 分析及其與產量QTL 的關系[J].作物學報,2001,27(6):915-922.
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
軍事文摘(2021年22期)2021-11-26 00:43:51
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
中國生殖健康(2019年2期)2019-08-23 08:12:08
