PLGF 參與Ang II 誘導的心臟成纖維細胞激活*
2013-12-23 06:26:56張曉歌陶小玲張賢銳
中國病理生理雜志 2013年4期
張曉歌, 陶小玲, 張賢銳, 蘇 立
(重慶醫科大學附屬第二醫院心內科,重慶400010)
血管緊張素Ⅱ(angiotensin II,AngⅡ)與其I 型受體(angiotensin II receptor 1,AT1)結合可通過多個環節刺激炎癥,促進細胞增殖及纖維化,導致心臟重構。研究提示,胎盤生長因子(placental growth factor,PLGF)與心血管疾病的病理過程有密切聯系。在人血管內皮細胞和平滑肌細胞,Ang II 可誘導PLGF 的表達[1],在缺血性心肌病患者中PLGF 隨心力衰竭的加重而升高[2]。心臟成纖維細胞(cardiac fibroblasts,CFs)在受到病理刺激后會發生增殖,其表型轉化為有分泌胞外基質功能的肌成纖維細胞,在心臟重構過程中具有重要作用[3]。心肌重構過程中,在血液動力學和神經體液激素的影響下,各種炎癥細胞被激活,分泌大量炎癥因子,加劇纖維化的發展。研究表明,可溶性血管內皮生長因子受體1(soluble vascular endothelial growth factor receptor-1,sFlt-1)能夠通過特異性的捕獲PLGF 而抑制其活性,減少腹膜炎癥過程中膠原的沉積,對腹膜纖維化有一定的抑制作用[4]。在心肌成纖維細胞中,阻斷PLGF 對Ang II 誘導的細胞增殖及纖維化是否具有抑制作用尚不清楚。本研究旨在探討PLGF 在Ang II 激活心臟成纖維細胞中的表達及其作用。
材 料 和 方 法
1 動物及主要試劑
出生2 ~3 d SD 大鼠,SPF 級,由重慶醫科大學動物中心提供。小鼠抗大鼠波形蛋白單克隆抗體和蛋白酶抑制劑試劑盒均購自Abcam;Ang II、胰蛋白酶和II 型膠原酶均購自Sigma-Aldrich;胎牛血清購自Gibco;重組人PLGF 購自Peprotech;α-平滑肌肌動蛋白(alpha-smooth muscle actin,α-SMA)和山羊單克隆抗體PLGF(anti-PLGF)購自Santa Cruz;兔抗大鼠ERK1/2 和p-ERK1/2 購自CST;DMEM/高糖培養基、兔抗山羊Ⅱ抗、山羊抗兔Ⅱ抗及山羊抗小鼠Ⅱ抗均購自北京鼎國公司;RIPA 蛋白裂解液、BCA 蛋白濃度測定試劑盒及小鼠抗大鼠α-tubulin 均購自碧云天公司;Trizol Reagent、逆轉錄、RT-PCR 試劑盒及引物均購自大連寶生生物公司。
2 主要方法
2.1 心臟成纖維細胞培養 無菌開胸剪取SD 大鼠心尖部(心室肌)組織,移入離心管后各加入3 mL濃度為0.08%胰蛋白酶和0.08%II 型膠原酶,37 ℃水浴中消化組織碎塊,重復3 ~4 次。收集各次消化所得細胞,用含10%胎牛血清的DMEM 高糖培養基在5%CO2、37 ℃培養箱中培養,差速貼壁60 min 后棄去培養液,剩余貼壁細胞即為心臟成纖維細胞。將傳代2 次的心臟成纖維細胞用于實驗研究。
2.2 WST-1 檢測細胞增殖 消化后用培養基吹散的細胞以4 000 cells/well 接種在96 孔板中,培養12 h 后無血清培養基饑餓24 h,藥物干預24 h 后,每孔加入WST-1 10 μL,5%CO2、37 ℃培養箱繼續孵育4 h,終止培養,振蕩混勻15 min 后在酶聯免疫檢測儀上470 nm 波長處測定吸光度值,用600 nm 的波長作為參考波長。
2.3 免疫印跡 常規提取心肌組織總蛋白,BCA 法測定各組蛋白濃度。SDS-PAGE 凝膠電泳,蛋白電轉移至PVDF 膜,封閉后分別稀釋Ⅰ抗[ERK1/2(1∶1 000)、p-ERK1/2(1 ∶1 000)、α-SMA(1 ∶200)、PLGF(1∶200)和α-tubulin(1∶1 000)]孵育,4 ℃過夜,根據Ⅰ抗來源選用相應HRP 標記的Ⅱ抗孵育,ECL 顯影,凝膠圖像分析系統進行圖像分析,以特異性目的蛋白條帶光密度值與對應α-tubulin 蛋白條帶吸光度值間的比值對蛋白表達量進行半定量分析。
2.4 免疫熒光檢測α-SMA 的表達 將細胞種在24孔板的載玻片上,培養12 h,饑餓24 h,給予100 ng/L PLGF 干預24 h,4%多聚甲醛固定15 min,加1%BSA 室溫下封閉非特異抗原1 h,沖洗后加入α-SMA Ⅰ抗4 ℃過夜,加入抗大鼠熒光標記Ⅱ抗37 ℃孵育1 h,熒光顯微鏡下觀察分析。
2.5 實時熒光定量PCR 收集細胞,用Trizol 提取總RNA,紫外分光法確定RNA 的濃度和純度,將其逆轉錄為cDNA。實時熒光定量反應條件:95 ℃預變性90 s 后進入PCR 循環:95 ℃變性10 s,60 ℃退火30 s,共40 個循環;擴增反應結束后,從65 ℃緩慢加熱到95 ℃,以建立PCR 產物的熔解曲線。每個標本均設置復管PCR 反應。反應體系:10 μL。引物序列如下:I 型膠原蛋白上游引物5'-TCTGACTGGAAGAGCGGAGAG-3',下游引物5'-GAGTGGGGAACACACAGGTCT-3';III 型膠原蛋白上游引物5'-ACAGATGCTGGTGCTGAGAAGA-3',下 游 引 物 5'-GCTGGAAAGAAGTCTGAGGAAGG-3';PLGF 上游引物5'-CTGCTGGGAACAACTCAACAGA -3',下游引物5'-CTACAGCGACTCAGAAGGACACA-3';內參 照 βactin 上游引物5'-ACGGTCACAGG TCATCACTATCG-3',下游引物5'-GGCATAGAGGTCT TTACGGATG-3'。
數據的分析采用下述表達公式:目的mRNA 表達水平=2-ΔΔCt。
3 統計學處理
計量資料以均值±標準差(mean ±SD)表示,多組均數比較采用單因素ANOVA 法進行統計分析,組間兩兩比較采用最小顯著性差異法(LSD 法),以P<0.05 為差異有統計學意義,統計軟件采用SPSS 17.0。
結 果
1 Ang II 對PLGF 表達的影響
據預實驗結果,Ang II(10-6mol/L)干預SD 大鼠的心臟成纖維細胞24 h,PLGF mRNA 表達水平最高,Ang II 可以增加PLGF 表達4.90 倍(P <0.01)。AT1受體阻斷劑telmisartan(10-6mol/L)和AngII(10-6mol/L)聯用干預后,PLGF mRNA 表達被抑制(P <0.05),見圖1。圖2 示Ang II 可以誘導PLGF蛋白水平的表達上調(P <0.05)。上述結果表明,Ang II 可能通過AT1受體調節PLGF 在心臟成纖維細胞內表達。
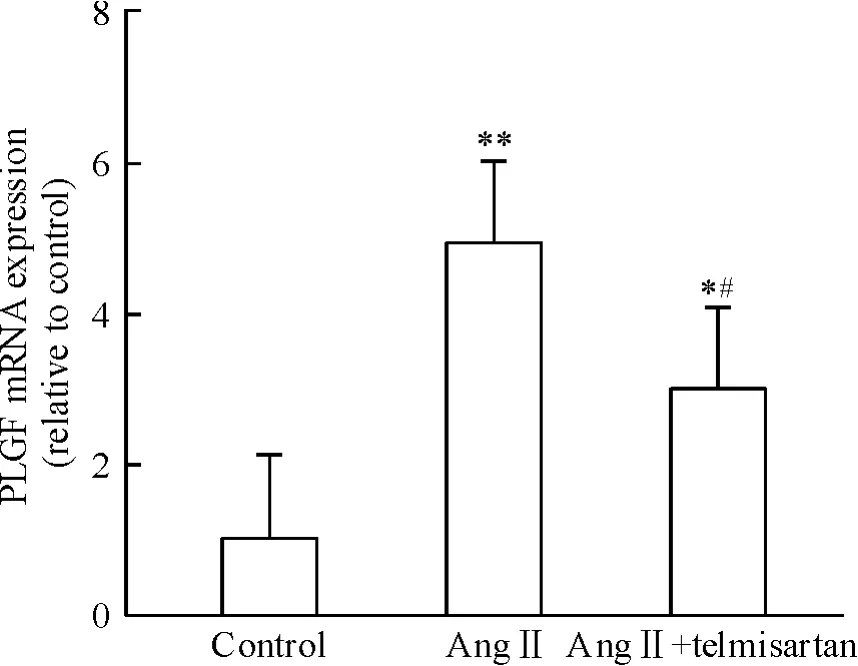
Figure 1. Effect of Ang II on the mRNA expression of PLGF.Mean ±SD.n =5. * P <0.05,**P <0.01 vs control;#P <0.05 vs Ang II.圖1 Ang II 對PLGF mRNA 表達的影響
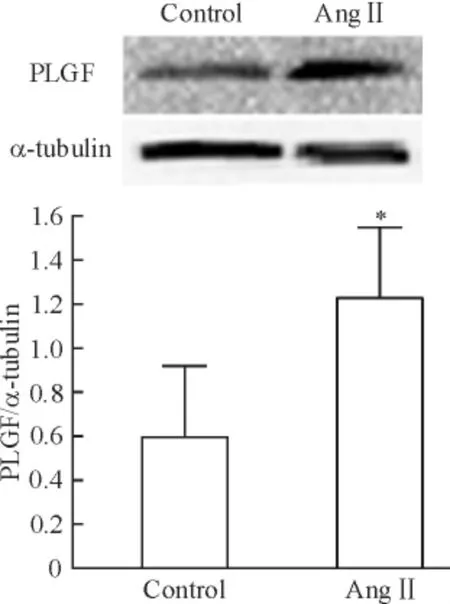
Figure 2. Effect of Ang II on the protein expression of PLGF in CFs.Mean±SD.n=4. * P <0.05 vs control.圖2 Ang II 對PLGF 蛋白表達的影響
2 PLGF 對心臟成纖維細胞的影響
運用不同濃度(10、50 和100 ng/L)PLGF 干預成纖維細胞24 h 后細胞增殖如圖3 所示,100 ng/L PLGF 明顯誘導細胞增殖達2.21 倍(P <0.05)。PLGF(100 ng/L)干預細胞24 h 后,免疫熒光檢測α-SMA,結果如圖4 所示,與對照組相比,細胞熒光強度增加。相同濃度的PLGF 分別干預36 h 和48 h 后檢測α-SMA 蛋白水平表達,如圖5 所示,PLGF 干預48 h 后誘導α-SMA 表達明顯增加(P <0.05)。以上結果提示,PLGF可以誘導心臟成纖維細胞增殖及向肌成纖維細胞轉化。PLGF 干預細胞0 min、5 min、15 min、30 min 和60 min 后,檢測p-ERK1/2 蛋白水平表達,如圖6 所示,60 min 時p-ERK1/2 蛋白水平明顯高于對照組(P <0.01)。
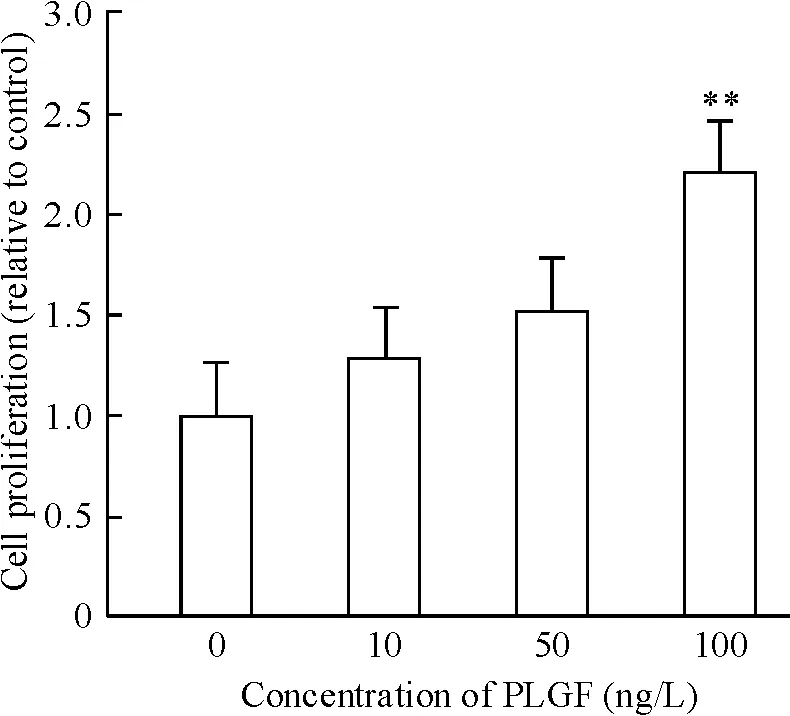
Figure 3. The proliferation of CFs treated with PLGF. Mean ±SD.n=5. **P <0.01 vs control (0 ng/L).圖3 PLGF 對CFs 增殖的影響
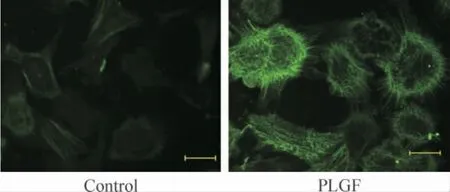
Figure 4. Effect of PLGF on α-SMA protein expression detected by immunofluorescence (×200).Bar:5 μm.圖4 免疫熒光評價PLGF 對α-SMA 蛋白表達的影響
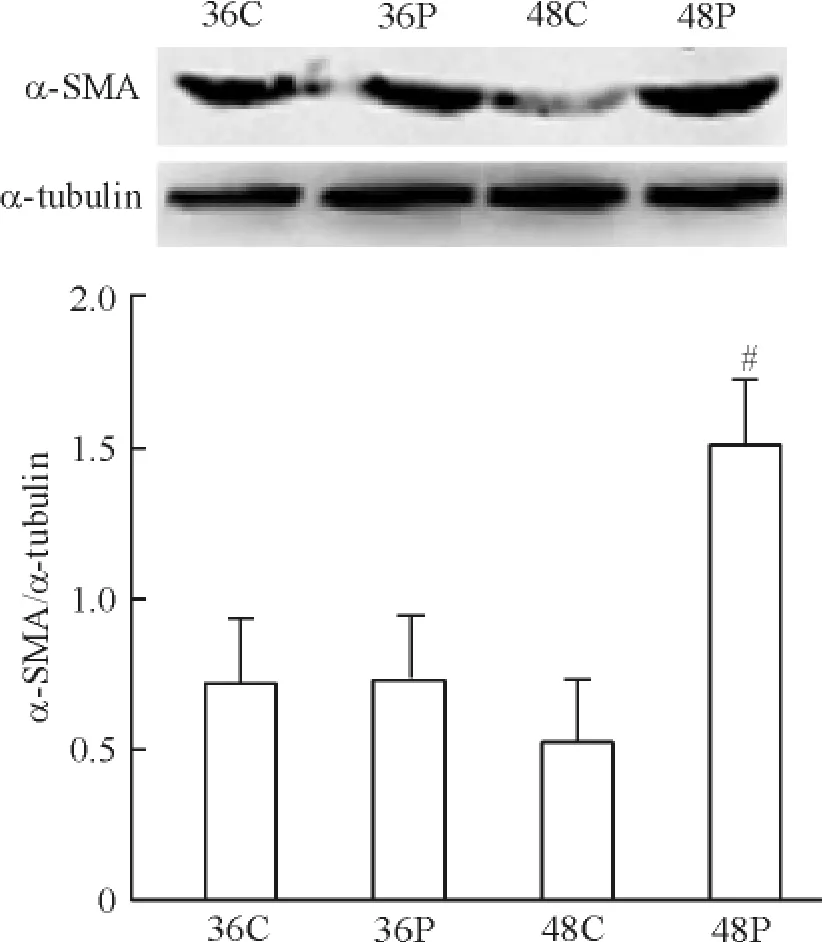
Figure 5. Effect of PLGF on α-SMA protein expression detected by Western blotting. 36C:36 h in control group;36P:36 h in PLGF group;48C:48 h in control group;48P:48 h in PLGF group.Mean ±SD. n =5.#P <0.05 vs 48C.圖5 PLGF 對α-SMA 蛋白表達的影響
3 Anti-PLGF 對Ang II 誘導心臟成纖維細胞增殖的影響
用不同濃度(5、10、20 和30 μg/L)PLGF 抗體(anti-PLGF)干預成纖維細胞,結果顯示濃度為20 μg/L 的anti-PLGF 抑制作用最明顯,干預24 h 后,結果如圖7 所示,Ang II 可以誘導細胞的增殖達2.23倍(P <0.05),Ang II 和anti-PLGF 聯用后可明顯抑制Ang II 誘導的細胞增殖(P <0.05)。上述結果提示anti-PLGF 可拮抗Ang II 誘導的成纖維細胞增殖效應。
4 Anti-PLGF 對Ang II 誘導α-SMA 表達影響
為探討anti-PLGF(20 μg/L)對拮抗Ang II 誘導的成纖維細胞表型轉化的影響,以Ang II 及anti-PLGF 同時干預成纖維細胞48 h,結果如圖8 所示。Ang II 可增加α-SMA 的表達1.93 倍(P <0.05),anti-PLGF 可 拮抗AngII 誘 導 的α-SMA 表達(P <0.05)。結果提示anti-PLGF 可拮抗Ang II 誘導的成纖維細胞表型轉化。
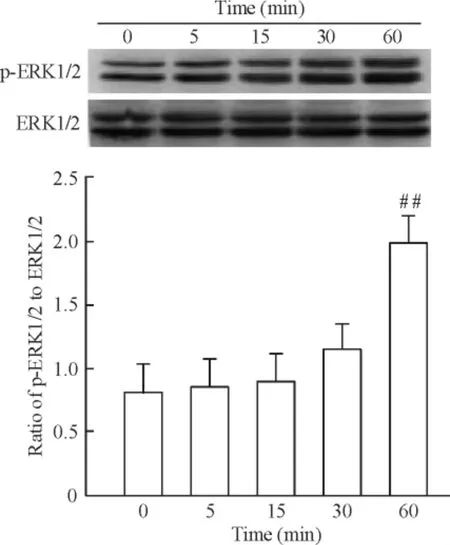
Figure 6. Effect of PLGF (100 μg/L)on the protein expression of p-ERK1/2 and ERK1/2 detected by Western blotting.Mean±SD.n=5. ##P <0.01 vs control (0 min).圖6 PLGF 對p-ERK1/2 表達的影響

Figure 7. Effects of anti-PLGF and Ang II on the proliferation of CFs.Mean ± SD. n =5. * P <0.05 vs control;#P <0.05 vs Ang II.圖7 Anti-PLGF 對Ang II 誘導細胞增殖的影響
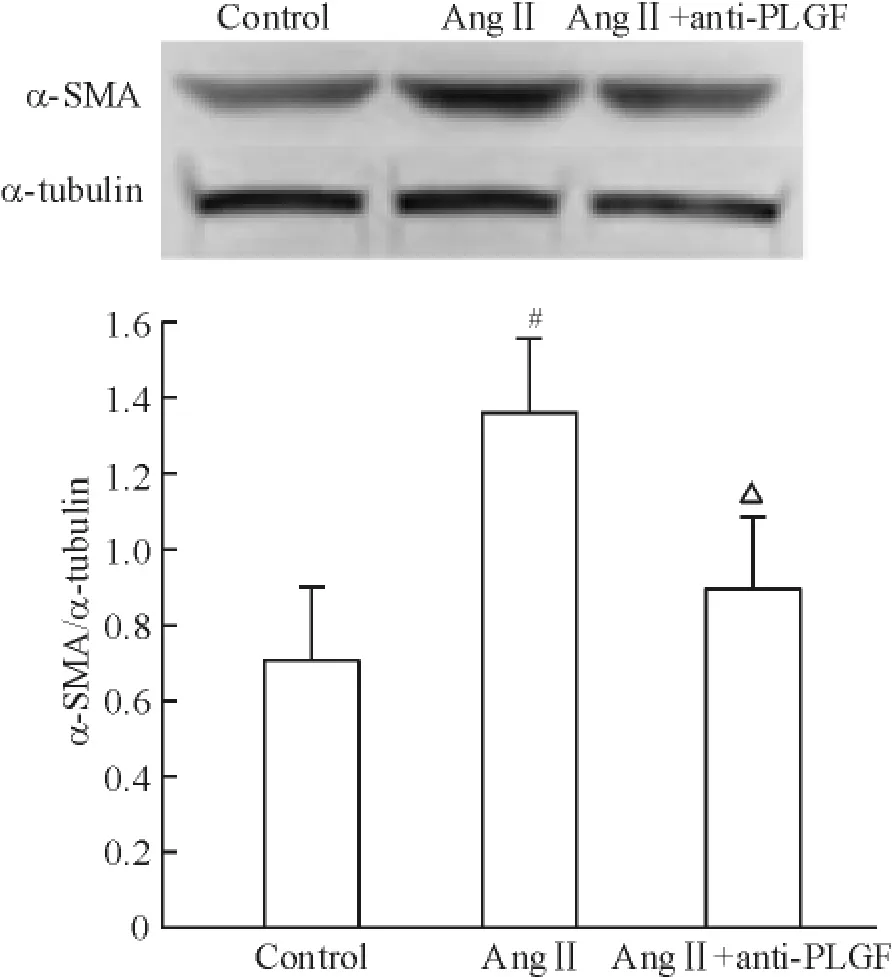
Figure 8. Effects of anti-PLGF on Ang II-induced α-SMA protein expression. Mean ± SD. n = 5. # P <0.05 vs control;△P <0.05 vs Ang II.圖8 Anti-PLGF 對Ang II 誘導α-SMA 蛋白表達的影響
5 Anti-PLGF 對Ang II 誘導I 型和III 型膠原蛋白mRNA 表達的影響
Anti-PLGF(20 μg/L)和Ang II 共同干預SD 大鼠的心臟成纖維細胞30 h,結果如圖9 所示。Ang II可增加I 型和III 型膠原蛋白的mRNA 表達分別為2.33 倍和2.47 倍(均P <0.05)。Anti-PLGF 可拮抗Ang II 誘導I 型和III 型膠原蛋白mRNA 表達上調(P <0.05)。
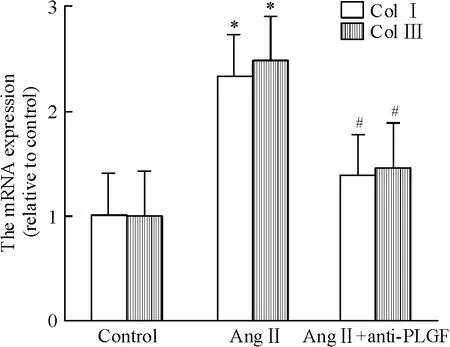
Figure 9. Collagen I and collagen III mRNA expression in various groups.Mean±SD.n=5. * P <0.05 vs control;#P <0.05 vs Ang II.圖9 Anti-PLGF 對Ang II 誘導膠原蛋白mRNA 表達的影響
討 論
本研究通過觀察Ang II 對心臟成纖維細胞PLGF 表達的影響及后者對細胞表型轉化、增殖和纖維化促進作用,以及給予anti-PLGF 阻斷PLGF 后對Ang II 介導的成纖維細胞激活的抑制效應,探討PLGF 參與Ang II 介導的心肌重構的可能機制。主要發現:(1)PLGF 可促進成纖維細胞增殖和表型轉化,激活ERK1/2;(2)Ang II 可通過AT1受體誘導成纖維細胞PLGF 表達升高;(3)anti-PLGF 對Ang II 誘導的成纖維細胞增殖、表型轉化及I 型和III 型膠原蛋白基因表達效應具有拮抗作用。
PLGF 是VEGF 家族成員之一,其堿基序列與血管內皮生長因子(vascular endothelial growth factor,VEGF)有高度的同源性。有研究表明,PLGF 在炎癥反應和心肌重構等病理過程中可能具有重要作用。在主動脈縮窄誘導心臟負荷增加的小鼠模型中,心肌細胞內PLGF 的表達升高[5],同時PLGF 在心臟超負荷狀態下,是腫瘤壞死因子α(tumor necrosis factor α,TNF-α)激活和巨噬細胞聚集的必需因子[6]。既往已有研究提示Ang II 可誘導VEGF 表達上調,同時激活血管炎癥反應[7],并通過AT1受體調節細胞增殖、細胞因子的產生及一些疾病的病理過程如高血壓、心臟的肥大及心臟纖維化等[1]。但有關PLGF在上述病理過程中的作用研究尚少,本研究結果提示,Ang II 可能通過AT1受體后水平,上調PLGF 在細胞中的表達,參與Ang II 介導的心血管疾病的病理過程。
PLGF 參與心血管重構的機制尚未完全闡明。研究表明,PLGF 可能通過破壞基質金屬蛋白酶抑制因子3/腫瘤壞死因子α 轉化酶軸的平衡等多種途徑激活心臟的炎癥反應,誘發或促進心肌重構[6,8]。本研究中,以PLGF 干預成纖維細胞后,細胞增殖明顯,且成纖維細胞向肌成纖維細胞表型轉化;同時短期給予PLGF 處理后,纖維化通路中的磷酸化ERK1/2 蛋白表達水平增高,而總ERK1/2 的表達水平無明顯改變,提示PLGF 可能通過影響細胞ERK1/2 蛋白的磷酸化水平產生生物學效應。既往研究證實,磷酸化ERK1/2 水平增高后,可啟動核內纖維化、增殖性相關基因的轉錄與表達,并成為心臟成纖維細胞趨化和增殖、膠原蛋白合成增加、心肌纖維化改變等效應的共同通路之一[9-10]。
細胞的炎癥反應可誘導或加重心肌纖維化過程[11-12],大量研究證實,PLGF 可激活機體內巨噬細胞,促使白細胞介素6、TNF-α 和單核細胞趨化蛋白1(monocyte chemotactic protein 1,MCP-1)表達升高,抑制其活性后可通過減少炎癥反應,阻斷疾病的進展[13-15]。
值得注意的是,盡管給予PLGF 干預細胞后,PLGF 本身對TGF-β、I 型和III 型膠原蛋白的mRNA 表達水平無顯著影響,但用anti-PLGF 阻斷PLGF 后可拮抗Ang II 誘導的成纖維細胞增殖和α-SMA 蛋白水平表達增加,同時降低Ang II 誘導I 型和III 型膠原蛋白mRNA 表達水平。最近,Van Steenkiste 等[16]關于肝纖維化的研究亦表明,PLGF 雖不能直接引起纖維化指標的顯著改變,但抑制PLGF 活性可顯著減少肝硬化小鼠肝纖維化及炎癥反應嚴重程度。另一項有關肝纖維化的研究顯示,anti-PLGF 干預或PLGF 基因敲除同樣可拮抗二乙基亞硝胺或四氯化碳誘導的肝纖維化[17]。以上2 項在肝細胞中觀察到的現象與本研究在心肌成纖維細胞中所觀察的結果近似。產生這一現象的具體機制仍不清楚。有作者推測,在心臟處于超負荷狀態下,機體內PLGF 與其受體Flt-1 可能通過激活心臟的炎癥反應,使心肌纖維化信號轉導通路中的ERK 或Akt 活性增加,最終導致心肌纖維化,這一效應可能需要病理狀態下其它周圍環境因素的協同作用[8]。我們推測Ang II 促細胞增殖及纖維化效應,可能部分需通過誘導PLGF 在心肌成纖維細胞中的表達而實現,這一過程發生于AT1 受體后水平。anti-PLGF 通過與PLGF 結合,抑制高濃度PLGF 對ERK1/2 的磷酸化效應,拮抗Ang II 的部分生物學效應。
綜上所述,本研究提示Ang II 促進心臟成纖維細胞內PLGF 表達,PLGF 可能為Ang II 下游調節因子,可通過ERK 途徑誘導細胞增殖及表型轉化,anti-PLGF 可拮抗Ang II 誘導的細胞增殖、表型轉化及I型和III 型膠原蛋白mRNA 的上調。PLGF 參與了Ang II 誘導的心肌重構過程。
[1] Pan P,Fu H,Zhang L,et al. Angiotensin II upregulates the expression of placental growth factor in human vascular endothelial cells and smooth muscle cells[J]. BMC Cell Biol,2010,11:36.
[2] Nakamura T,Funayama H,Kubo N,et al. Elevation of plasma placental growth factor in the patients with ischemic cardiomyopathy[J]. Int J Cardiol,2009,131(2):186-191.
[3] Turner NA. Therapeutic regulation of cardiac fibroblast function:targeting stress-activated protein kinase pathways[J]. Future Cardiol,2011,7(5):673-691.
[4] Motomura Y,Kanbayashi H,Khan WI,et al. The gene transfer of soluble VEGF type I receptor (Flt-1)attenuates peritoneal fibrosis formation in mice but not soluble TGF-β type II receptor gene transfer[J]. Am J Physiol Gastrointest Liver Physiol,2005,288(1):G143-G150.
[5] Accornero F,van Berlo JH,Benard MJ,et al. Placental growth factor regulates cardiac adaptation and hypertrophy through a paracrine mechanism[J]. Circ Res,2011,109(3):272-280.
[6] Carnevale D,Cifelli G,Mascio G,et al. Placental growth factor regulates cardiac inflammation through the tissue inhibitor of metalloproteinases-3/tumor necrosis factor-αconverting enzyme axis:crucial role for adaptive cardiac remodeling during cardiac pressure overload[J]. Circulation,2011,124(12):1337-1350.
[7] Zhao Q,Ishibashi M,Hiasa K,et al. Essential role of vascular endothelial growth factor in angiotensin II-induced vascular inflammation and remodeling[J]. Hypertension,2004,44(3):264-270.
[8] Carnevale D,Lembo G. Placental growth factor and cardiac inflammation[J]. Trends Cardiovasc Med,2012,22(8):209-212.
[9] Li L,Fan D,Wang C,et al. Angiotensin II increases periostin expression via Ras/p38 MAPK/CREB and ERK1/2/TGF-β1 pathways in cardiac fibroblasts[J].Cardiovasc Res,2011,91(1):80-89.
[10]Eliceiri BP,Klemke R,Str?mblad S,et al. Integrin αvβ3 requirement for sustained mitogen-activated protein kinase activity during angiogenesis[J]. J Cell Biol,1998,140(5):1255-1263.
[11]敖杰男,馮慧娟,吳 迪,等.炎癥細胞因子的心力衰竭發病過程中的作用[J]. 中國病理生理雜志2005,21(10):2062-2064.
[12]Liu Y,Liao Y,Cheng X,et al. TGF-β1of cardiac tissue and ventricular remodeling in rats with acute myocardial infarction[J]. Chin J Pathophysiol,2005,21(12):2305-2309.
[13]Yoo SA,Yoon HJ,Kim HS,et al. Role of placenta growth factor and its receptor flt-1 in rheumatoid inflammation:a link between angiogenesis and inflammation[J].Arthritis Rheum,2009,60(2):345-354.
[14]Luttun A,Tjwa M,Moons L,et al. Revascularization of ischemic tissues by PlGF treatment,and inhibition of tumor angiogenesis,arthritis and atherosclerosis by anti-Flt1[J]. Nat Med,2002,8(8):831-840.
[15] Markovic' M,Ignjatovic' S,Dajak M,et al. Placental growth factor as short-term predicting biomarker in acute coronary syndrome patients with non-ST elevation myocardial infarction[J]. South Med J,2010,103(10):982-987.
[16]Van Steenkiste C,Ribera J,Geerts A,et al. Inhibition of placental growth factor activity reduces the severity of fibrosis,inflammation,and portal hypertension in cirrhotic mice[J]. Hepatology,2011,53(5):1629-1640.
[17] Heindryckx F,Bogaerts E,Coulon SH,et al. Inhibition of the placental growth factor decreases burden of cholangiocarcinoma and hepatocellular carcinoma in a transgenic mouse model[J]. Eur J Gastroenterol Hepatol,2012,24(9):1020-1032.
