科爾沁沙地沙漠化恢復過程中冷蒿種群的擴散對策研究
2014-01-02 06:00:16孫英杰李衍青趙愛芬黃玉清周愛萍羅艾瀅徐廣平姚月鋒
草業學報 2014年1期
關鍵詞:生長
孫英杰,李衍青,趙愛芬,黃玉清,周愛萍,羅艾瀅,徐廣平,姚月鋒
(1.廣西壯族自治區中國科學院廣西植物研究所,廣西桂林541006;2.中國地質科學院巖溶地質研究所,廣西桂林541004;3.魯東大學生命科學學院,山東煙臺264000)
科爾沁沙地屬于我國半干旱半濕潤的農牧交錯帶脆弱區域,由于人類的過度放牧、開墾和樵采等,使得該地區成為我國沙漠化比較嚴重的地區之一。近年來,隨著退耕還林還草等一系列防治工程的實施,科爾沁沙地生態環境整體上呈現出逐漸恢復的趨勢。在退化生態系統恢復過程中,植被恢復是退化生態系統重建的重要部分,因此,對沙地沙漠化恢復過程中植物的適應生理生態方面研究顯得尤為重要。
冷蒿(Artemisia frigida)屬菊科小灌木,廣泛分布于半干旱草原和沙地[1-2]。是科爾沁沙地耐性較強的一種重要固沙植物種,它具有較強的補償性生長能力,其數量及繁殖方式隨著演替退化的各個階段變化而變化,這一特性對植物群落的結構與功能具有重要影響[3]。因此,冷蒿作為沙地植物群落演替后期的優勢種及草原指示種[4],在科爾沁沙地退化生態系統恢復過程中尤其是群落演替后期發揮著重要的作用。冷蒿具有旺盛的營養繁殖能力和一定的有性繁殖能力。沙埋、動物踩食、刈割等因素都會對其繁殖特性產生影響[5-8]。
國內外對冷蒿進行了大量的研究,這些研究主要關注于冷蒿個體形態[9]、冷蒿的形態學特性[10]、飼用價值、生態型、根系分布等生態學特性[11],冷蒿揮發性油的分離和鑒定[12],冷蒿的抗性生理與其他生理特性[13-16],冷蒿繁殖對放牧的響應[5-6,17-18]、生物量資源分配[7,19-20],冷蒿草原土壤種子庫[21]、冷蒿群落特性[22-24]等方面。而關于科爾沁沙地冷蒿種群擴散現象的研究少見報道。筆者在研究科爾沁沙地植被恢復的野外調查時發現,冷蒿有2種差異明顯的生長表現型,分別為枝條密集、木質化程度高且直立生長的冷蒿和枝條較柔軟且匍匐生長的冷蒿。本研究分析了這2種生長型不同的個體地上與地下部分的形態特征及繁殖特點,進而了解直立型冷蒿和匍匐型冷蒿在種群擴散上的地位及作用。旨在期望能為科爾沁沙地的植被恢復和土地荒漠化治理中植物篩選方面提供理論依據或可行性措施。
1 材料與方法
1.1 研究區概況
研究樣地位于內蒙古奈曼旗境內(120°55'E,42°41'N),平均海拔360 m。該區域屬溫帶半干旱大陸性氣候,干旱且多大風[25],年均氣溫6.5℃,1月份平均氣溫 -12.7℃,7月份平均氣溫 23.7℃,年均降水量 362.3 mm,主要集中在6,7,8月,年均蒸發量1 935.4 mm,雨熱同季。8級以上大風年均21次,年均風速達3.2~4.0 m/s,其中10~15 d沙塵暴天氣主要出現在春季。
植被種類主要以灌木、半灌木為主,由菊科、禾本科、豆科、藜科、蒺藜科、蓼科植物組成。近百年來由于人為的干擾,使該區的原生植被受到破壞,沙漠化日益嚴重,植被中的喬木層已基本消失,草本層退化,灌木層發育強烈。目前植被以灌木、多年生和一年生草本植物組成的沙生植物為主,主要植物種包括冷蒿、小葉錦雞兒(Caragana microphylla)、杠柳(Periploca sepium)、黃柳(Salix gordejevii)、差巴嘎蒿(Artemisia halodendron)、黃蒿(A.scoparia)、豬毛菜(Salsola collina)、糙隱子草(Cleistogenes squarrosa)、狗尾草(Setaria viridis)等。該區域土壤貧瘠,主要為風沙土、沙質栗鈣土、沙質草甸土和沼澤土[26]。
1.2 研究方法
1.2.1 樣地選擇與實驗設計 1)調查樣地:中國科學院奈曼沙漠化研究站1994年建立的禁牧的放牧圍欄(樣地A)、與放牧圍欄相鄰的東南方向的固定沙丘(樣地B)。
樣地A處冷蒿種群基本為直立生長,個體界限清楚,在封育圍欄內成斑塊狀分布,約200株左右,主要集中生長在相對低洼的區域。斑塊中心少有其他植物生長而靠近斑塊邊緣則有較多的地錦(Euphorbia humifusa)、糙隱子草、灰綠藜(Chenopodium glaucum)等植物分布。
樣地B處于典型恢復過程中的半固定沙丘。土壤有風蝕,地表形成淺風蝕坑。屬于冷蒿+杠柳群落。該樣地與玉米地、人工楊樹林、放牧圍欄毗鄰;土壤為沙土質土壤;小半灌木為冷蒿、差巴嘎蒿;草本植物以黃蒿、霧冰藜(Bassia dasyphylla)、灰綠藜為主;以及灌木植物——杠柳。冷蒿為匍匐生長,呈現大面積斑塊狀分布,從群落中心到群落邊緣,冷蒿密度逐漸變小并呈現向外分散生長的態勢。
2)野外沙埋試驗地:微氣象站東面的固定沙丘(樣地C)。此區域冷蒿植株由微氣象站的固定沙丘頂部的冷蒿種群擴散而來。該樣地內冷蒿為優勢植物種,差巴嘎蒿逐漸退出僅余少量幾株。此處冷蒿個體以直立型和匍匐型2種表現型生長,且生長旺盛,種群密度較高。
1.2.2 調查時間與方法 1)野外調查:2009年6月29日,在樣地A處的冷蒿斑塊內,以種群中最原始的冷蒿植株(調查時植株已經死亡,但根系仍在土壤中,大部分枯枝完好保留)為原點,向該株冷蒿周圍種群邊緣的10個冷蒿植株做樣線,用經緯儀測量每條樣線間的角度,并記錄下每條樣線上冷蒿個體在樣線上的投影長度,以代表其蓋度[27]。
2009年6月30日,在樣地B中選擇30株匍匐型冷蒿作為研究對象。調查每株冷蒿產生的分株的數量和分株與基株間的距離。
2009年7月24日,在樣地B處,選擇30株直立型冷蒿和30株匍匐型冷蒿標記,記錄每株冷蒿樣株的平均高度、最高枝高度、最低枝高度、叢輻、營養枝數量、生殖枝數量及花序數。然后,分別將標記的冷蒿的地上部分從莖基部剪掉,裝入袋中標記后帶回實驗室。用分層法[20,28-29]從冷蒿莖的底端向最頂端將植株分為0~10 cm、10~20 cm、20~30 cm、30~40 cm、40~50 cm、50 cm以上6層,把每層生物量放入編號信封中,然后放入80℃的烘箱中烘24 h,24 h后取出稱量,記錄下每株冷蒿的各層生物量干重。
選擇典型的直立型冷蒿和匍匐型冷蒿個體,將其地下部分小心挖出,去掉根系部分的沙土和地上部分,用掃描儀掃描2株冷蒿的根系。
2)野外沙埋實驗:2009年分別在6和7月選擇22株352個枝條做沙埋處理實驗,以確定不同沙埋時間對冷蒿生長和分株的影響。6月2日,在樣地C中選擇自然狀態下生長良好的直立型冷蒿和匍匐型冷蒿各4株作為實驗對象。從每株冷蒿中各選取6枝健康且生長態勢一致的枝條做沙埋處理。7月2日,試驗地點與6月2日相同,方法類似,在樣地中選擇生長良好的直立型和匍匐型冷蒿各7株,每株沙埋處理10個枝條。
2009年8月22日用小鐵鍬挖出2次處理的所有枝條,從枝條基部與母株分離并小心去掉枝條上的沙土標記裝袋。小心去掉枝條附著的沙土及其他植物的細根。分別測量出每次沙埋處理的每個枝條生根枝條數,不定根的數量,不定根的長度。
1.3 數據分析與處理
1.3.1 封育圍欄內冷蒿種群擴散圖的繪制 1994年在因過度放牧引起的沙漠化區域進行了圍欄封育,樣地本底調查時冷蒿種群按照Drude多度等級應為“個別種”(Un),1998年調查時即形成冷蒿小斑塊,所發現植株均為直立型生長。種群擴散調查圖以樣地內最大的1株冷蒿為中心,向各個方向輻射,每條線間隔30°,圖上以黑點代表冷蒿植株在樣線上的位置,2009年則以1998年調查地內最原始的冷蒿植株O為中心,根據經緯儀測量出來的各個樣線的角度畫圖,圖上每小格代表實際距離1 m。

1.3.2 匍匐型冷蒿調查及2次沙埋處理枝條的處理 利用Microsoft Excel 2003對30株匍匐型冷蒿分株調查結果進行統計分析,并計算出每株冷蒿母株產生的平均分株數量,以及其分株擴散的平均最遠和最近距離。
2 結果與分析
2.1 冷蒿個體地上部分的形態及生物量特征
直立型冷蒿與匍匐型冷蒿的地上形態差異主要表現在個體高度、枝條數、叢輻等方面(表1)。從總體上來看,直立型冷蒿的高度、枝條數、每株花序數均大于匍匐型冷蒿。但直立型冷蒿的平均叢輻(61.1 cm×51.0 cm)小于匍匐型冷蒿(63.5 cm×57.8 cm),兩者之間無較大差異。由于調查樣地冷蒿種群大小的限制,30株冷蒿個體間生長差異的變異性較大。

表1 2種表現型冷蒿個體地上生長形態比較Table 1 The comparison of growth form aboveground of two phenotypes of A.frigida
由t-檢驗結果可知,直立型和匍匐型冷蒿的最低枝高度、平均枝高、營養枝條數和平均每株每枝的花序數均存在顯著差異(P<0.05),直立型冷蒿植株的生殖枝一般多于營養枝。但最高枝高度及生殖枝條數均無顯著差異。雖然花序數與種子產量有很大關系,但種子形成又受到天氣條件、傳粉方式等因素的影響,所以花序數不能用于預測冷蒿的種子產量。
通過對冷蒿個體調查結果分析可知枝條密集生長的直立型有較高的地上生物量,直立型冷蒿植株有76.67%的單株生物量大于20 g,而匍匐型冷蒿中有50%。從構建的生物量金字塔(圖1)可以看出,在地上部分的高度及平均單株地上生物總量上,直立型冷蒿均優于匍匐型冷蒿。在0~10 cm即靠近植株基部,匍匐型冷蒿的生物量大于直立型冷蒿,而且基部的生物量大于10~50 cm以上部分的任一層。直立型冷蒿10~50 cm以上這5層的生物量大于匍匐型冷蒿,其株高達50 cm以上,但匍匐型冷蒿卻在50 cm以內。以上結果可能與冷蒿的枝條木質化及匍匐型冷蒿生長得較低矮直接相關。
2.2 冷蒿地下根系形態生長特性
調查發現直立型冷蒿是由種子萌發的有性生殖產生,因此植株的根系類似于直根系(圖2左圖),這種形態有利于冷蒿植株從更深的土壤層次中吸取水分以滿足整個植物的生長發育所需。而匍匐型冷蒿的根系都有與其他植株相連的老枝條,形態如圖2右圖所示,o點為冷蒿基株的基部,a~e分別為基株產生的分株處。隨著時間的推移,各分株與基株脫離形成一個新的冷蒿植株個體。冷蒿枝條被沙埋后,在適宜環境下會生長出不定根進而在基株上形成不同大小的分株,從而形成了叢輻較大的匍匐型冷蒿個體。因此,匍匐型冷蒿個體比直立型冷蒿對地上空間利用的更充分,而直立型冷蒿個體則在土壤縱向的資源利用上占優勢。
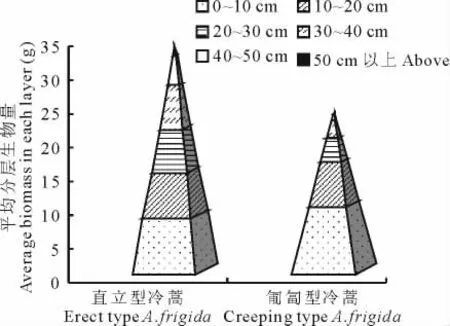
圖1 2種表現型冷蒿的地上生物量比較Fig.1 The comparison of aboveground biomass of two phenotypes of A.frigida
2.3 匍匐型冷蒿的分株能力

圖2 直立型冷蒿根系(左)和匍匐型冷蒿(右)根系形態圖Fig.2 The morphology of erect type’roots(left)and creeping type’roots(right)of A.frigida
在適宜的水分條件下,冷蒿被沙埋的枝條發展成匍匐莖并向上生長出不定芽,向下產生不定根。匍匐莖是冷蒿為其克隆生長增繁地上枝條、地下不定根的營養器官。冷蒿通過這種分枝方式實現其在生境內獲取必需資源的策略。隨著時間的推移,不定芽會慢慢長成枝條,不定根則汲取土壤中的養分并向土壤深處活動,形成冷蒿個體。分株最終脫離基株,成為單個的冷蒿植株,完成了冷蒿數量上的擴張。冷蒿對環境的適應特性是通過其生長特征(生物量、分株數)和形態特征(匍匐莖、節間距和不定根間距)來表現的,生長型主要包括分株高、枝條長、枝條數量等[30]。
通過對30株匍匐型冷蒿的調查發現,產生的分株數多數在10株以下,最少的產生3株,最多的可達16株,擴散的距離為4~35 cm,大部分分株與基株距離在10~20 cm之間。這與匍匐型冷蒿的枝條長度相關。這種繁殖特性是在沙漠化地區其不斷被沙埋、被放牧牲畜啃食等環境干擾下種群能夠生存下去的重要因素。
2.4 不同沙埋時間對冷蒿枝條生根的影響
被沙埋的冷蒿枝條產生不定根與否以及數量的多少與沙埋時間有關,分析認為與生長期間的氣溫、降水量有很大關系。實驗地區6,7,8月月平均氣溫分別為20.28,23.44,23.25℃。降水主要集中在4-7月,雖然8月份降水較少,但從整體上來看,適度的氣溫和降水量都是冷蒿枝條生根的有利條件。
由表2可知,在不同沙埋時間處理下,冷蒿生根枝條數在每株冷蒿處理枝條中所占比例、每株的平均最長根長度及每株生根數總量上均表現為匍匐型冷蒿>直立型冷蒿,平均每株冷蒿的生根數量在0~5 cm根長度中占大部分。而且第1次處理下2種表現型冷蒿根的長度在5~10 cm與10 cm以上所占比例大致相近,第2次處理下卻表現出根長為5~10 cm的根數多于10 cm以上的趨勢。可以推測出在適宜環境下,處理時間越長越利于冷蒿枝條生根。此外,同一處理時間下,2種表現型冷蒿的平均最長根長度及各級根長間的數量均接近,這說明同一條件下2種表現型冷蒿的枝條生根能力無差異。

表2 2種表現型冷蒿枝條的沙埋處理結果比較Table 2 The comparison of sand buried branches of two phenotypes of A.frigida
2.5 封育圍欄內冷蒿種群擴散特征
自1994年沙地圍欄封育后,在無人為干擾及牲畜采食、踐踏的自然條件下,冷蒿種群從最初的僅幾株發展到1998年的119株以上,以選定的測量植株為基點,擴散最遠的冷蒿植株與基株灌叢間距離達到2.7 m(圖3),位于東偏南30°方向。并且冷蒿在各條樣線上成斑塊狀聚集分布。基株的西方無冷蒿生長,大部分冷蒿主要集中在東北方向、東方而且與基株間的距離也較遠,西南方向有少數冷蒿分布但西方和西偏北30°方向的冷蒿卻更少。
2009年的調查結果顯示:在十多年的無干擾條件下,冷蒿種群無論在數量上還是規模上都遠遠超過了1998年。在冷蒿基株周圍,擴散最遠的冷蒿植株距離基株17.1 m,位于東偏南52°(圖4)。從整體上來看,冷蒿種群的擴散方向及分布情況與1998年調查結果類似,冷蒿植株主要分布在東北方向、東方,其次是東南方,西北方向和西南方向、西方冷蒿較少甚至沒有冷蒿生長。而且在擴散距離上也是東北,東方和東南方的距離最遠,西南、西北方向距離較近。

圖3 1998年恢復樣地的冷蒿種群擴散圖Fig.3 The diffusion of A.frigida population in restoration plot in 1998
實地調查中發現,該恢復樣地中的冷蒿植株均為直立型冷蒿。許多研究發現[10-11,19],直立型冷蒿枝條中生殖枝占絕大部分,營養枝較少,主要以產生種子的方式繁殖新個體。
圖3和圖4中出現的冷蒿種群的擴散方向和距離的趨勢類似。這種結果可能與冷蒿種子較小,種子散布時受風的影響較大,尤其是在科爾沁沙地這種風沙環境有關。科爾沁沙地冬春季節受蒙古冷高壓控制,盛行偏西或偏北風[31],所以冷蒿秋季成熟的種子順著主風向方向散布的量最多,距離最遠,而向相應的逆主風向方向散布的種子量少,距離最近。已有研究[8]表明,種子散布的數量和距離,能夠反映種群數量動態的潛在能力或種群擴展空間生態位的潛在趨勢。
從表3中可知,從數量上來說,1~4,6,10號樣線上的冷蒿分布較多,5,7,8,9號樣線上相對較少均在5株以下,即北偏東、南偏東52°方向冷蒿數量比南偏東、南偏西和北偏西方向多,占種群的大多數。從冷蒿在每條樣線上所占的總長度來看,變化趨勢與數量相似,冷蒿的叢輻大小卻表現出相反的趨勢,南偏東52°的平均個體最大,叢輻為1.37 m,其次是南偏西和北偏西方向的冷蒿個體,北偏東方向冷蒿叢輻較小,在0.26~0.37 m之間。這可能與樣線上冷蒿個體的數量多有很大關系。
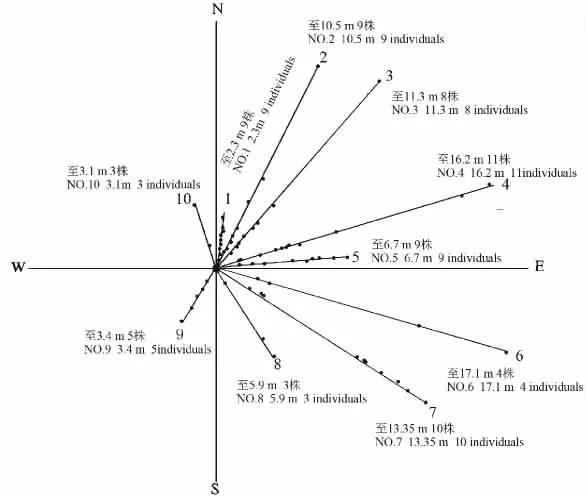
圖4 2009年恢復樣地冷蒿種群擴散圖Fig.4 The diffusion of A.frigida population in restoration plot in 2009
3 結論與討論
在長期的自然選擇下,冷蒿進化出了許多適應干旱環境的生理生態的特征,其中之一便是形成了2種表現型即直立型冷蒿和匍匐型冷蒿。不同表現型冷蒿繁殖方式上也存在差異。直立生長的冷蒿為有性繁殖,叢生枝條密集,地上地下部分均為明顯的獨立個體;匍匐生長的多為克隆繁殖,枝條較稀疏,地下部分常有與其他植株連結的老枝條。在沙漠化地區圍欄封育開始時冷蒿種群擴散以種子繁殖為主,隨著植被的恢復物種增多,土壤水分減少,種間競爭強度更大,會出現克隆繁殖。冷蒿2種繁殖方式的結合不僅能提高和維持種群的擴散能力,其聚集群體更利于提高種群的生存和競爭力,使得它在植被更新、群落恢復過程中起著非常重要的作用[32-33]。

表3 各樣線上冷蒿個體的分布Table 3 The distribution of A.frigida on each line transect
從植株個體高度上來看,直立型冷蒿高于匍匐型冷蒿,并且前者的枝條總數、每株花序數均多于后者。但直立型冷蒿的平均叢輻小于匍匐型冷蒿,冷蒿個體間的生長差異的波動性較大。從冷蒿的單株地上生物總量結果中可知,直立型冷蒿的總量大于匍匐型冷蒿,且波動范圍較廣。在地上生物量的分配上,直立型冷蒿靠近基部的生物量小于匍匐型冷蒿,但基部以上的部分均大于后者。這與直立型冷蒿個體高于匍匐型冷蒿,后者的枝條較低矮有很大關系。
在干旱半干旱的沙地,季節性的降水對植株種子萌發及枝條生根具有較大的影響[34]。通過沙埋實驗發現沙埋時間的長短及降水對冷蒿生根數量和最長不定根的長度影響較大,但同一沙埋時間處理下2種表現型冷蒿的不定根數量和最長不定根的長度無差異。直立型冷蒿是由1粒種子經過萌發發育而成,其根系分布較深,能充分利用土壤較深處的水分和養分。而匍匐型冷蒿由基株和分株構成,其分株能擴散到距離基株35 cm處,而且基株能產生多達16個分株,這就使得其地下根系構成的網絡狀結構在土壤的橫向資源利用上遠優于直立型冷蒿。
圍欄封育樣地的冷蒿種群以最原始植株為點逐漸向東北方與偏東方向擴散分布。同時植株在其擴散方向上呈斑塊狀聚集分布。造成這一現象可能是與冷蒿種子較小并能隨著風向傳播到較遠距離,以及科爾沁沙地冬春季盛行偏西或偏北風有關。這一結論與楊允菲和祝玲[35]的研究即種子散布的數量及方向與地區的季風氣候有很大的關系相符合。
與其他地區相比,科爾沁氣候更干旱,土壤含沙量高,在其沙漠化恢復過程中,黃蒿、冷蒿等菊科植物和蟲實(Corispermum dilutum)等藜科植物則會大量出現在植被演替后期[36-37]。而冷蒿獨特且有效的繁殖方式加快了冷蒿整個種群的擴散速度,這樣就更利于科爾沁沙地退化生態系統恢復進程。因此,一方面可以結合2種表現型冷蒿的不同繁殖方式進行植被恢復,另一方面可以通過適當沙埋的方式促使冷蒿植株枝條生根產生分株以擴大其生長范圍從而獲得更多的資源,在進一步促進沙丘固定的同時,能為更多物種的入侵定居打下堅實的基礎。
[1]李玉強,趙哈林,移小勇,等.沙漠化過程中科爾沁沙地植物-土壤系統碳氮儲量動態[J].環境科學,2006,27(4):635-640.
[2]趙哈林,郭軼瑞,周瑞蓮,等.植被覆蓋對科爾沁沙地土壤生物結皮及其下層土壤理化特性的影響[J].應用生態學報,2009,20(7):1657-1663.
[3]王靜,楊持,劉美玲,等.保護冷蒿草原對防止沙質草原沙漠化作用的分析[J].內蒙古大學學報(自然科學版),2002,5:558-562.
[4]趙彩霞,鄭大瑋.內蒙古冷蒿小禾草放牧草原退化與恢復對策研究[J].草業學報,2004,13(1):9-14.
[5]楊持,寶音陶格濤,李良.冷蒿種群在不同放牧強度脅迫下構件的變化規律[J].生態學報,2001,21(3):405-408.
[6]祁永,杜麗霞,韓建國.放牧對冷蒿繁殖特性的影響[J].草地學報,2007,15(2):168-192.
[7]李金花,李鎮清,劉振國.不同刈牧強度對冷蒿生長與資源分配的影響[J].應用生態學報,2004,15(3):408-412.
[8]邢福,王艷紅,郭繼勛.內蒙古退化草原狼毒種子的種群分布格局與散布機制[J].生態學報,2004,24(1):143-148.
[9]張華,李鋒瑞,Yasuhito S,等.固沙林庇護區內土壤-植物系統的變化特征[J].水土保持學報,2003,17(3):144-148.
[10]占布拉,陳世璜,張昊,等.冷蒿的特性和生態地理分布的研究[J].內蒙古農牧學院學報,1999,20(1):1-7.
[11]李敬忠,趙書元,劉忠.內蒙古凡種野生牧草生態型的研究III冷蒿[J].內蒙古畜牧科學,1997,3:7-8.
[12]Atazhanova G A,Dembitskii A D,Yakovleva T D,et al.Composition of the essential oils of Artemisia radicans and A.frigid[J].Chemistry of Natural Compounds,1999,35(4):427-429.
[13]杜占池,楊宗貴.溫度對冷蒿枝條光合和暗呼吸作用的影響[J].植物學通報,1993,(S1):14-15.
[14]周海燕,趙愛芬.冷蒿和差巴嘎蒿抗旱性機理的比較研究[J].中國沙漠,1998,18(增刊2):56-60.
[15]王靜,楊持.冷蒿抗寒生理特性的研究[J].內蒙古大學學報(自然科學版),2002,33(6):673-676.
[16]王海鷗,周瑞蓮.高溫脅迫下差巴嘎蒿和冷蒿的生理變化及其抗熱性研究[J].中國沙漠,1999,19(增刊1):64-67.
[17]Hutchings M J,de Kroon H.Foraging in plants:the role of morphological plasticity in resource acquisition[J].Advances in Ecological Research,1994,25:159-238.
[18]王靜,楊持,王鐵娟.冷蒿(Artemisia frigida)種群在放牧干擾下構件的變化[J].生態學報,2006,26(3):960-965.
[19]李金花,李鎮清.不同放牧強度下冷蒿、星毛委陵菜的形態可塑性及生物量分配格局[J].植物生態學報,2002,26(4):435-440.
[20]王靜,楊持,王鐵娟.放牧退化群落中冷蒿種群的生物量資源分配的變化[J].應用生態學報,2005,16(12):2316-2320.
[21]蘇德畢力格,李永宏,雍世鵬,等.冷蒿草原土壤可萌發種子庫特征及其對放牧的響應[J].生態學報,2000,20(1):43-46.
[22]左小安,趙學勇,趙哈林,等.科爾沁沙質草地群落物種多樣性、生產力與土壤特性的關系[J].環境科學,2007,28(5):945-951.
[23]王慶鎖,董學軍,陳旭東.油蒿群落不同演替階段某些群落特征的研究[J].植物生態學報,1997,21(6):531-538.
[24]李衍青,孫英杰,張銅會,等.科爾沁沙地不同演替階段冷蒿群落的結構特征[J].應用生態學報,2011,22(7):1725-1730.
[25]劉新民,趙哈林,趙愛芬.科爾沁沙地風沙環境與植被[M].北京:科學出版社,1996:191-211.
[26]樸順姬,王振杰,顏秀靈,等.科爾沁沙地差巴嘎蒿種群生態位適宜度分析[J].植物生態學報,2006,30(4):593-600.
[27]裴世芳,傅華,陳亞明,等.放牧和圍封下霸王灌叢對土壤肥力的影響[J].中國沙漠,2004,24(6):763-767.
[28]孫啟忠,韓建國,桂榮,等.科爾沁沙地達烏里胡枝子生物量研究[J].中國草地,2001,23(4):21-26.
[29]孫啟忠,韓建國,桂榮,等.科爾沁沙地敖漢苜蓿地上生物量及營養物質累積[J].草地學報,2001,9(3):165-170.
[30]祁永,杜麗霞,韓建國.放牧對冷蒿繁殖特性的影響[J].草地學報,2007,15(2):168-172.
[31]鄭春芳,李鳳山.科爾沁草地沙漠化成因及其可持續發展初探[J].草業科學,2006,23(4):12-16.
[32]葛俊,邢福.克隆植物對種間競爭的適應策略[J].植物生態學報,2012,36(6):587-596.
[33]董鳴,于飛海,陳勁松,等.克隆植物生態學[M].北京:科學出版社,2011:99-103.
[34]羅亞勇,趙學勇,黃迎新,等.三種一年生藜科沙生植物出苗對沙埋深度和水分條件的響應[J].草業學報,2009,18(2):122-129.
[35]楊允菲,祝玲.松嫩平原堿化草甸朝鮮堿茅種子散布機制的分析[J].植物學報,1995,37(3):222-230.
[36]趙哈林,大黑俊哉,李玉霖,等.人類放牧活動與氣候變化對科爾沁沙質草地植物多樣性的影響[J].草業學報,2008,17(5):1-8.
[37]李玉霖,孟慶濤,趙學勇,等.科爾沁沙地流動沙丘植被恢復過程中群落組成及植物多樣性演變特征[J].草業學報,2007,16(6):54-61.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
