騰格里沙漠南緣唐古特白刺克隆分株生長格局及枝系構型分析
2014-01-02 06:00:18周資行李真焦健李毅令凡
草業學報 2014年1期
關鍵詞:生長
周資行,李真,焦健* ,李毅,令凡
(1.甘肅農業大學林學院,甘肅蘭州730070;2.中國藥科大學生命科學與技術學院,江蘇南京210009)
克隆植物在自然界廣泛存在[1-2]。克隆生長使一個有性生殖產生的合子(即基株)具有眾多遺傳上相同的獨立新個體,即克隆分株,分株間不同的連接方式,形成植株不同的克隆構型[3]。克隆構型主要決定于間隔子長度,如相鄰分株間匍匐莖或根狀莖長度,基株產生的分株數和分枝角度等形態指標[4-5],植物的克隆構型不僅在物種間可能不同,而且不同環境中同一物種的克隆構型也可能不同,表現出形態可塑性。在資源異質性分布的環境中,克隆植物通過克隆構型的可塑性變化來調整資源獲取對策,不僅包括決定資源吸收結構的水平放置的間隔物長度和分枝強度的可塑性,而且也包括決定資源吸收結構垂直放置的分株本身的形態可塑性[4]。關于克隆植物構型與分株生長格局的研究已有報道,但多以草本植物研究為主[6-11],如馮圖等[12]對林緣曠地、林緣和林下3種不同生境中淫羊藿(Epimedium brevicornum)克隆構型及其分株種群特征進行了研究;廖明雋等[13]研究內蒙古錫林河流域草地、林地、沙地3種生境下的羊草(Leymus chinensis)種群,結果表明在不同生境中,分株個體在環境因子影響下產生株高、間隔子、分枝強度、分枝角度及根狀莖長等的可塑性變化,進而引起克隆植物分株種群特征的變化。對于灌木克隆構型及分株種群方面的研究較少[14],鮮見水分梯度下旱生灌木無性系分株生長格局及其枝系構型研究。
騰格里沙漠南緣,北接沙漠、東南臨黃土高原、西南為高山(高原)區,位于中國西北干旱荒漠區、東部季風區與西南青藏高寒區交匯部位,生態環境極其脆弱,近50年來在人類作用和區域氣候變化雙重驅動下,土地沙漠化十分嚴重,亟需進一步加強區域植被保護工作,維護生態安全。唐古特白刺(Nitraria tangutorum)為我國的特有種,廣泛分布于我國西北荒漠區的鹽化湖盆,鹽化戈壁、山前洪積扇和沙化荒灘。唐古特白刺為典型的克隆灌木,具有特殊的個體形態,萌生能力強,主要采用根出條型(壓條型)進行克隆生長[15],被沙土埋覆后能產生大量不定根形成新的植株,攔蓄和固定大量流沙形成固定和半固定的灌叢沙包,具有良好的防風固沙作用,是穩定沙漠、保護綠洲以及植被恢復和重建的優良灌木,在干旱、半干旱地區占據著十分重要的生態地位。近年來關于唐古特白刺的研究有諸多報道[16-18]。這些研究多集中在白刺的地理分布規律、種群繁殖策略和生理特性等方面,而對白刺克隆分株生長格局與枝系構型的研究尚未見報道。筆者首次嘗試選取騰格里沙漠南緣景泰、古浪和民勤三地及對照樣地蘭州的唐古特白刺種群分布區設置樣地,研究唐古特白刺克隆分株生長格局和枝系構型的形態可塑性變化特征,了解其種群的覓食行為和對環境條件的反應,探討克隆種群對資源異質性的響應特點,以期能更真實地揭示其在自然條件下種群的克隆生長適應機制,尋求唐古特白刺資源的保護策略,并為白刺的資源恢復和利用提供科學依據。
1 材料與方法
1.1 研究區自然概況
沿騰格里沙漠南緣自南向北選擇景泰白墩子(JBDZ)、古浪馬路灘林場(GMLT)和民勤勤峰農場(MQQF)及對照地蘭州西固南山(LZNS)四地唐古特白刺種群天然分布區設置樣地。四地年平均降雨量在113.2~311.7 mm之間,年均蒸發量在1446.4~2604.3 mm之間,土壤含水量在1.82% ~9.34%之間,水分條件呈明顯的遞減趨勢,研究區水熱條件見表1。
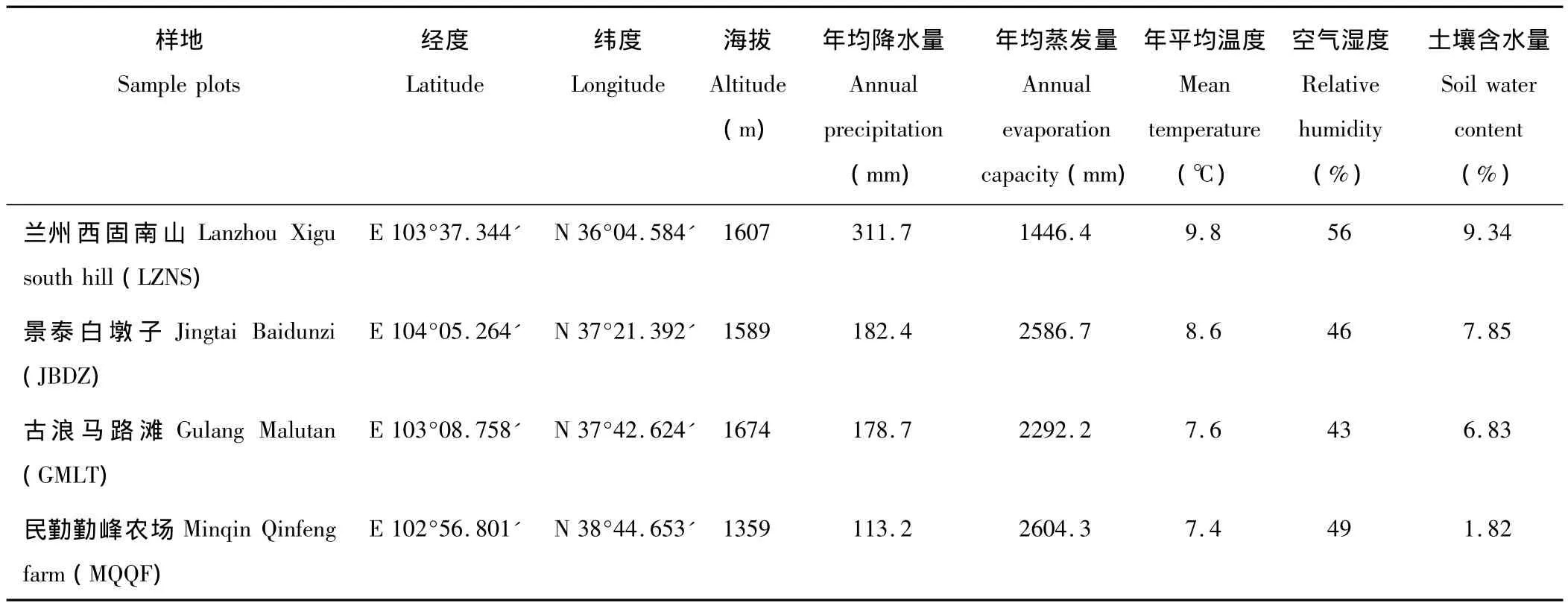
表1 研究區水熱條件Table 1 Hydrothermal conditions of the study regions
1.2 樣地調查
2010年7月-2011年8月,在四地選擇白刺大小分布均勻及無人為破壞之處各設置400 m2樣地,在每個樣地內采用“全挖法”進行“跟蹤挖掘”,即沿萌蘗根方向逐一挖出每個無性系分株。通過挖掘找出所有的連結關系,年齡最大者以及獨立的植株即為基株,其余記為分株。選取所有大齡(基株6齡以上)白刺克隆植株,以其克隆分株為對象,進行克隆生長格局及其枝系構型的研究。測計每個白刺無性系擴張域(克隆種群地上部分范圍),分株數,分株冠幅,分株高度,分株地徑,分株間距;一級分枝數目、分枝長度、分枝角度、分枝徑粗(離主莖1 cm處),二級、三級與當年生分枝數目、分枝長度,分枝角度、分枝徑粗(離上一級分枝0.5 cm處)。白刺克隆種群擴張域和冠幅均采用十字交叉法測量,并近似看作橢圓。
1.3 統計方法
1.3.1 克隆分株特征 1)分株密度:分株密度的計算,不以樣地面積為基礎,而嘗試以白刺克隆種群地上擴張域為底數,這樣似乎能夠更好反映小尺度下白刺克隆種群微生境特征。

式中,分株數為白刺的克隆分株總和,n為樣地內選測的白刺無性系個數。
2)分株間距頻率:對分株間距的分析分兩步進行,首先分別統計4個樣地白刺分株間距頻率,判斷頻率分布特征,用Kruskal Wallis法進行比較,以判斷生境內部克隆構型的變化差異;然后在此基礎上用Two-Sample Kolmogorov-Smirnov Test法,將每一樣地各白刺克隆植株與其他樣地白刺分株間距頻率分布做兩兩比較[19]。用其中出現的無差異次數占總比較次數的百分比,作為衡量2個樣地間白刺分株間距相似程度的標準。
3)體斥效應v值:隨機行走模型是對植物克隆生長的有效的近似描述,結合自回避行走模型確定白刺克隆構型,計算體斥效應v值,參照李鎮清的方法[5]。
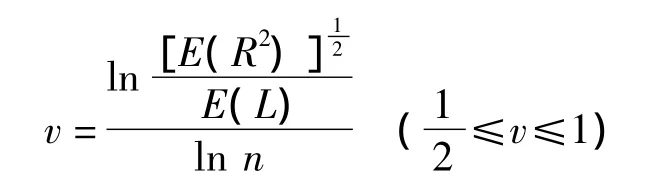
式中,假設唐古特白刺擴張域內資源環境同質,則E(L)為分株間距的算術平均。v代表體斥效應,等于期望的位移值E(R)與期望的步長值E(L)之比的對數與潛在克隆分株數(n)的對數之比。當v=1/2時,植株體斥效應最小且非密度制約,這種克隆植物屬密集型(phalanx),當v=1時,對應一維自回避行走,這種克隆植物屬游擊型(guerilla)。密集型和游擊型是兩個極端的克隆構型,其間還存在許多過渡類型的連續體。
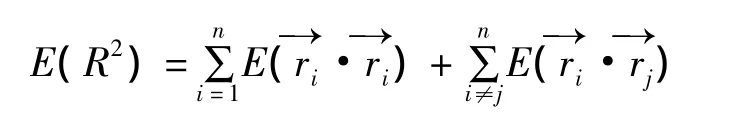
1.3.2 枝系構型 統計各樣地選取的大齡唐古特白刺所有分株及其上枝系,枝序的確定方法有多種,鑒于信息量和應用廣度,本文選取Strahler法確定枝序[20]。即在植物冠層內,由外及內確定枝序,外層的第一小枝為第一級,兩個第一級相遇為第二級,兩個第二級相遇后則為第三級,依此類推,如有不同枝級相遇,相遇后則取較高的作為枝級。
1)枝密度:同上面的計算原則。

式中,n為樣地內選測的白刺無性系個數。
2)枝徑比:
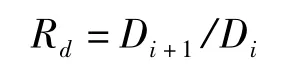
式中,Di+1和Di分別是第i+1和第i級枝條的直徑。
3)分枝率與分枝強度:采用Whithey[21]提出并經Steigraeber和Waller[22]修改的方法計算總體分枝率(Rb):
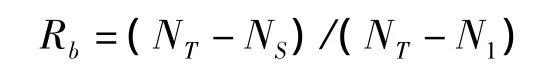
式中,NT為所有枝級中枝條的總數,NS為最高枝級的枝條數,N1為第一級的枝條總數。某一級枝條數與下一個高枝級的枝條數之比即為逐步分枝率(Ri∶i+1):
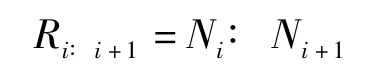
式中,Ni為第i級的枝條總數,Ni+1為第i+1級的枝條總數。
枝系分枝強度應用逐步分枝率加權平均表示,權重為逐步分枝率中低級分枝數目比例。
4)分維數:針對唐古特白刺植冠及不同級別分枝在空間分布中的分維數進行計算。運用分形幾何原理研究植物分枝格局。
樹冠分維數(DF)根據Mandelbrot[23]的分維數理論,假定k為連續分枝級別枝長之比,k<1,r為每一級分枝數目,可推導出樹冠的分維數為:
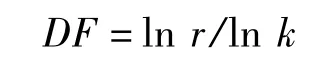
分枝分維數(DBA):參照何明珠等[24]的方法計算。
植物各級分枝的數目(SB)與枝長(LB)、分維數(D)之間的關系可按下式進行計算:
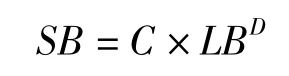
式中,C為比例系數(常量)。將上式變換得:
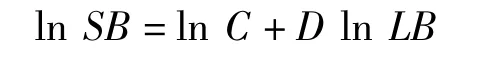
可見,SB與LB在雙對數坐標下存在一種線性關系,D為該直線的斜率。因此,分維數的求算方法是在雙對數坐標下對SB和LB的一系列值進行線性回歸,所得擬合直線的斜率(或其轉換),即為分枝分維數值(DBA)。
利用SPSS Statistics 17.0,采用One-Way ANOVA進行分析,并作Duncan多重比較,調查統計與數學整理采用Excel 2003,各量以平均值±標準誤形式表示。
2 結果與分析
2.1 克隆生長格局
2.1.1 克隆分株形態特征 四地唐古特白刺無性系分株特征存在一定差異,特別是分株密度和分株間距的差異顯著(表2)。對照點LZNS白刺各形態指標與沙漠南緣三地均存在顯著差異,除了分株間距外,分株其他指標均明顯大于沙漠南緣三地;JBDZ和GMLT水分條件比較接近,兩地白刺表現出一定程度的相似性,但分株數、分株地徑與枝密度等指標差異較大;MQQF分株間距最大,而擴張域、分株冠幅、分株株高與分株密度則最小,與其他樣地差別明顯,可能是由于水分條件相對較差所致。
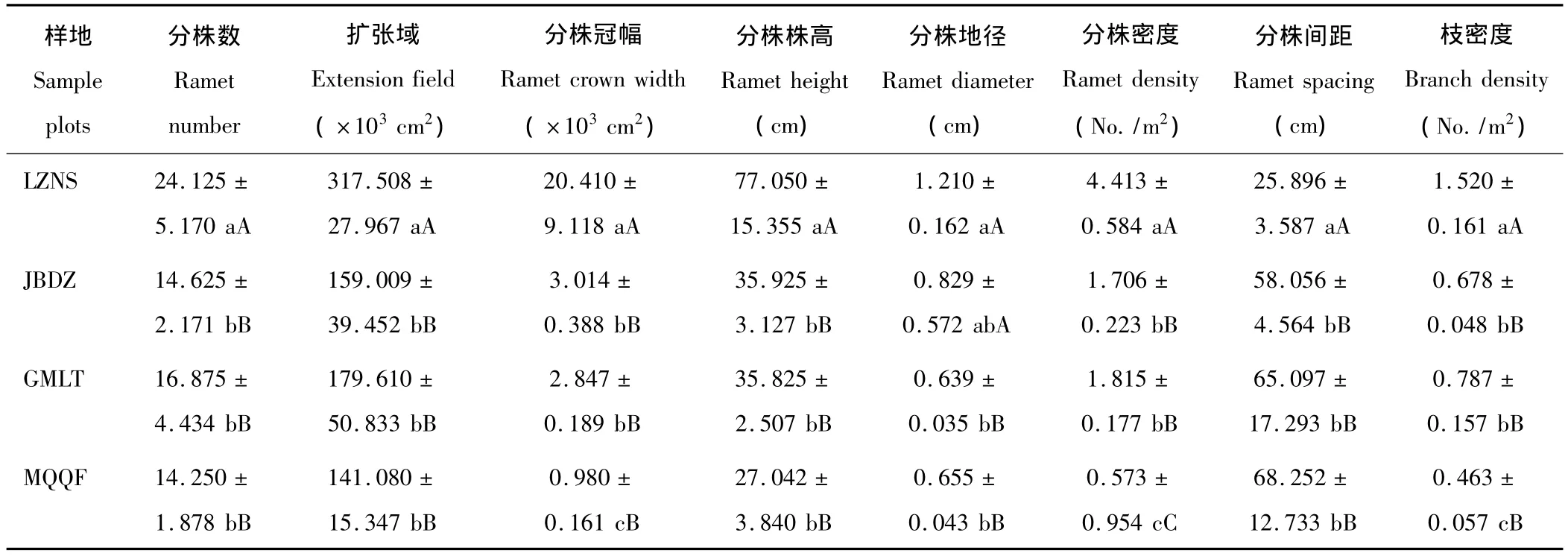
表2 唐古特白刺克隆分株形態特征比較Table 2 Comparison of morphological characteristics of N.tangutorum clonal populations in 4 plots
2.1.2 分株間距頻率 唐古特白刺平均分株間距頻率見圖1。對照點LZNS以小間距為主,其頻率顯著高于沙漠南緣三地,JBDZ與GMLT則表現出中等間距頻率較大,MQQF則大間距(>200 cm)頻率較高,各地分株間距呈現隨水分梯度減少較為明顯的遞增趨勢,可能由于在水分條件較差時,植株的密度制約較小,在有限的資源水平下盡可能的擴張水分覓食范圍,以提高生態優勢。Kruskal Wallis檢驗表明,分株間距差異性LZNS(P=0.75)、JBDZ(P=0.37)和GMLT(P=0.43)均不顯著,說明這三地白刺克隆構型變化不大,而 MQQF(P<0.05)分株間距分布的變化顯著,說明區域資源的分布不均一,導致不同立地條件的白刺無性系具有不同的克隆構型。
用Two-Sample Kolmogorov-Smirnov Test法,逐一將各樣地的每株白刺與其他樣地分株間距頻率分布做兩兩比較,結果發現:LZNS與JBDZ、GMLT和 MQQF的相似度分別為 63.8%,62.6%和 36.3%,JBDZ與GMLT和MQQF的相似度為81.7%和51.8%,而GMLT與MQQF的相似度為53.1%,整體上呈現較強的樣地關聯性,但MQQF白刺無性系與其他三地的差異較大,具有相對獨立性。
2.1.3 體斥效應v值 結合自回避行走模型確定唐古特白刺克隆生長格局,統計體斥效應v值如表3。MQQF與其他三地存在極顯著差異,由于當v=1/2時,植株體斥效應最小且非密度制約,克隆生長格局屬密集型,LZNS、JBDZ和GMLT唐古特白刺克隆生長格局均趨近密集型,特別是LZNS幾乎為典型密集格局。由于當v=1時,對應一維自回避行走,克隆生長格局屬游擊型,MQQF的v值為0.716,克隆生長格局介于密集型和游擊型之間,有一定的游擊趨勢。v值偏度系數大小與Kruskal Wallis檢驗吻合。
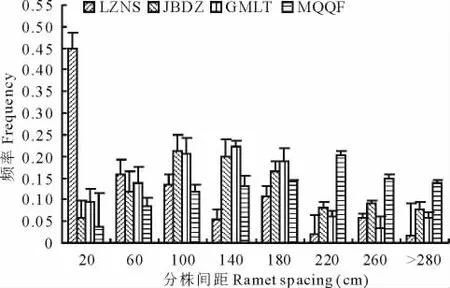
圖1 唐古特白刺克隆植株平均分株間距頻率比較Fig.1 Comparison of average ramets distance frequency of N.tangutorum clonal populations in 4 plots
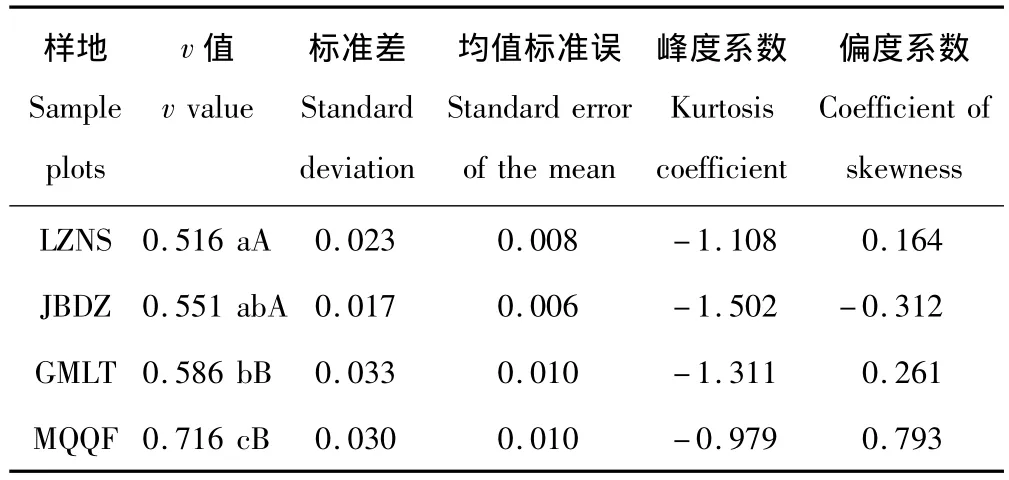
表3 唐古特白刺克隆植株體斥效應Table 3 Comparison of the excluded volume effect of N.tangutorum clonal populations in 4 plots
2.2 枝系構型
2.2.1 枝系形態參數 四地白刺枝系形態存在較顯著差異(表4),且LZNS與MQQF多數形態指標差異極顯著。隨著水分梯度減少,四地唐古特白刺各級分枝指標多數呈遞減趨勢,這與無性系克隆整合有密切的關系。但LZNS的各級分枝角最小,JBDZ和GMLT各級分枝角大小基本一致,而MQQF最大,這可能與無性系擴張空間大小有關,LZNS分株密度大,受克隆分枝擴張空間制約,無性系分枝大多向高層空間擴張,以期獲得更多的光資源。
白刺無性系枝徑比表現較為復雜(圖2),各級枝徑比除了MQQF的3級∶當年生,均以LZNS最大,LZNS水分條件優于沙漠南緣三地,無性系徑向生長受水分限制較小,各級分枝徑粗的差異不大,分枝間克隆整合效益顯著,白刺無性系通過徑粗的普遍生長,達到最廣泛的資源儲備,增強植株整體穩定性,以應對未來可能的環境脅迫;MQQF各級枝徑比多數為四地中最小,這可能是在水分脅迫下,白刺無性系克隆擴張受限,主要資源用以維持植株最基本的生存,一級分枝作為植株的基礎,資源優先分配,以確保植株的正常生存,從而導致1級∶2級枝徑比最小;而3級∶當年生枝徑比卻最大,可能是無性系力求資源均勻分配,以達到最大的擴張能力,白刺無性系克隆整合的結果,體現惡劣環境下的形態可塑性。JBDZ和GMLT由于水分條件比較接近,白刺無性系枝徑比表現出較強的相似性,枝徑比均呈現隨枝級遞減趨勢。
枝系分枝強度的大小決定著克隆植物利用資源的能力,分枝強度隨資源水平的上升而增加[25]。四地唐古特白刺無性系分枝強度有一定差異(圖2),LZNS白刺分枝強度顯著大于沙漠南緣三地。這表明,在水分條件較好時,白刺無性系能夠萌發更多的分蘗,在充分利用有限資源的基礎上,力爭進行種群的擴張,以占據更為有利的生境;反之,由于水分條件的限制,分枝強度的增加會導致死亡率的增加,更多的分蘗可能枯死,意味著資源與能量的浪費。四地唐古特白刺無性系分枝強度整體上呈現較為明顯的隨水分下降的線性遞減趨勢。
分枝率統計結果如表5所示,總體分枝率體現枝條產生分枝的能力,四地總體分枝率僅MQQF較小,由于資源相對貧乏,無性系的各級分枝間的克隆整合致使總體分枝率顯著下降。逐步分枝率,特別是1級∶2級分支率是支持枝與帶葉枝的數目比,能直接反映植物體的生存活力。四地1級∶2級分支率以對照點LZNS白刺最大,表明該地白刺無性系生存活力旺盛。逐步分枝率隨枝級的增加而減小,則說明各地白刺無性系資源并不主要用于分枝數量的增長。
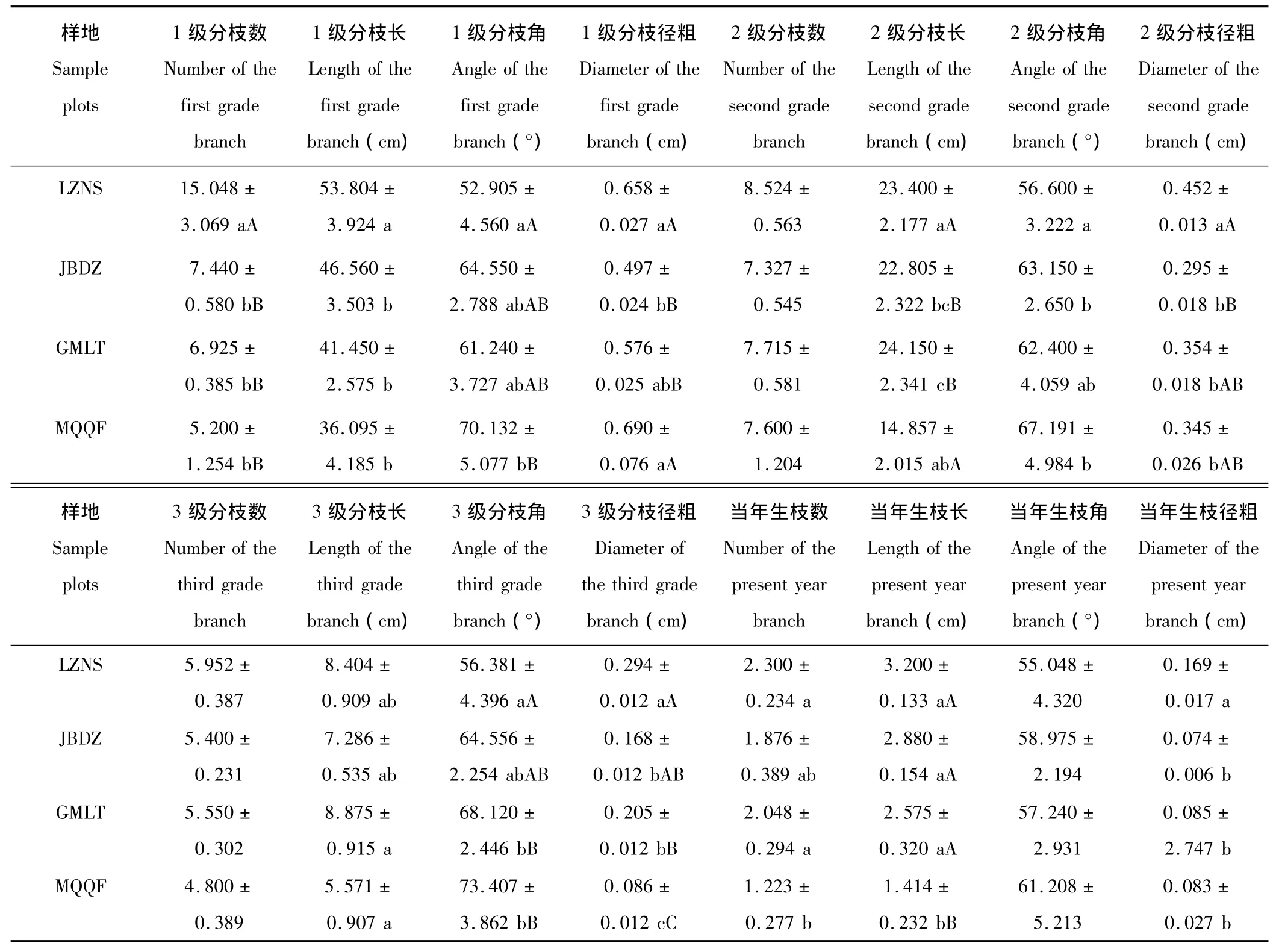
表4 唐古特白刺克隆種群枝系形態參數Table 4 Comparison of branching morphological parameters of N.tangutorum clonal populations in 4 plots
2.2.2 枝系分維數 通過分析各級分枝數與分枝長度,統計樹冠分維數(DF)和分枝分維數(DBA),見圖3。四地的唐古特白刺無性系各級分枝樹冠分維數DF差異不大,只在2級∶3級出現波動,整體上LZNS和MQQF的趨勢一致,但是LZNS各級樹冠分維數均大于MQQF,JBDZ和GMLT的趨勢非常相似,且各級樹冠分維數差距不大;LZNS的樹冠分維數較大,說明植物枝系的空間格局比較復雜,其占據空間的能力以及利用空間資源的潛能均較高。水分條件較好時,枝系構件的空間分布趨向復雜化,這在1級∶2級和2級∶3級體現都較為明顯,隨著水分的減少,無性系為減少枝葉構件間的競爭作用,樹冠分維數呈下降趨勢,枝系的空間分布趨于簡單,向空間擴展的能力較弱。但LZNS和MQQF在3級∶當年生上表現反常,LZNS由于水分條件較好,為了更廣泛的獲取環境資源,無性系擴張需求大,新生枝條萌發頻繁;MQQF白刺3級∶當年生增大,也是為了獲取更多的資源,特別是光資源。在資源水平相對貧乏時,白刺無性系會首先將資源分配給1級分枝,其次供給新梢生長。隨著水分條件梯度遞減,四地白刺分枝分維數DBA也呈現逐漸減小的趨勢,與樹冠分維數DF的趨勢較為一致。
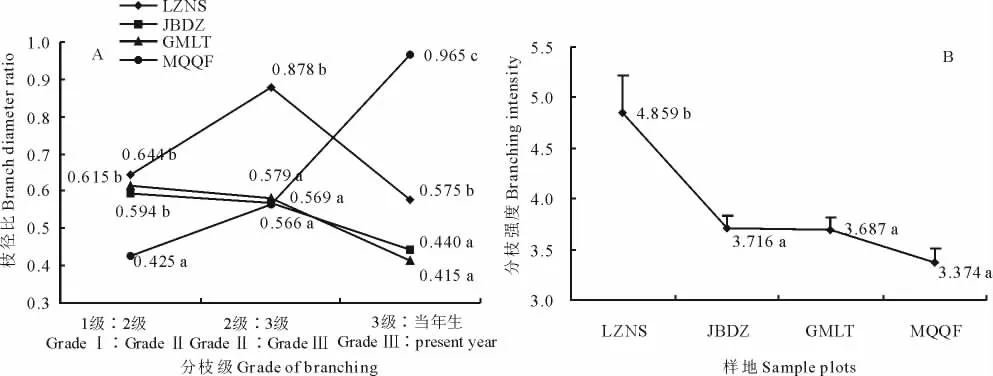
圖2 唐古特白刺克隆植株枝徑比與分枝強度Fig.2 Comparison of branching diameter ratio and intensity of N.tangutorum clonal plants in 4 plots
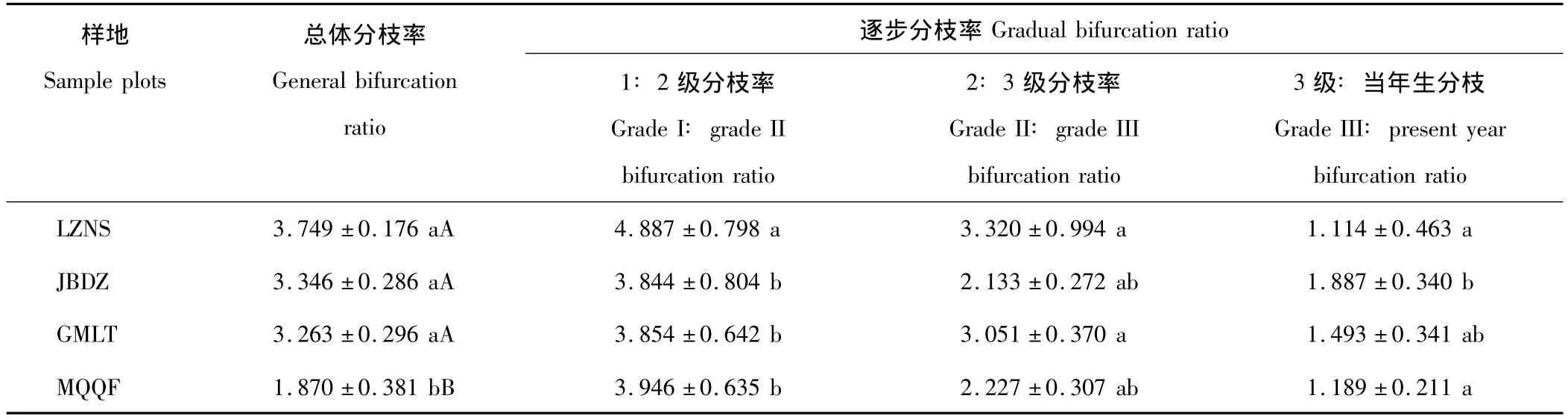
表5 唐古特白刺克隆植株分枝率Table 5 Comparison of bifurcation ratio of N.tangutorum clonal populations in 4 plots
3 結論與討論
唐古特白刺無性系分株特征表現出較強的形態可塑性,特別是分株密度和分株間距的差異最顯著。LZNS的水分條件較好,根狀莖和間隔子長度逐漸增大,克隆分株數量增加,通過分株集聚增強對生境資源的獲取,最大程度利用生境資源和占據有利的生存空間;隨著水分條件遞減,無性系分株數量逐漸減少,分株密度減小,分株間距增大,尤以MQQF表現最為明顯。在水分脅迫下,無性系為了盡可能獲取微生境資源,增加了對地下部分投資即增加間隔子和地下根莖長度,擴大分株間距,減小分株密度,以期增強種群擴散能力,占據更多生境資源和生存領域,更好的維持種群生存、繁殖和擴散[26]。本研究結果與此相似,不同樣地白刺無性系分株格局存在明顯規律性,分株間距頻率統計顯示,LZNS白刺無性系較短的分株間距頻率明顯要高于其他三地。
“游擊型”和“密集型”用來描述克隆植物的生長格局特征,也常常用于描述同一克隆植物構型可塑性變化的趨勢[27],在水分條件相對較差時,許多無性系植物的生長格局由密集型轉變成游擊型[28]。我們的研究也有相似的結果,通過自回避行走模型v值比較表明,隨著水分條件遞減,分株生長格局的游擊性越來越強。LZNS幾乎為典型密集格局,JBDZ和GMLT唐古特白刺克隆生長格局均趨近密集型,而MQQF克隆生長格局介于密集型和游擊型之間,有一定的游擊趨勢。對不同樣地分株間距的量化比較分析,可以加深對其形態可塑性的理解,白刺無性系分株通過改變聚集強度和空間占據能力,對環境的差異做出有效的響應,有利于增強種群對異質生境資源的獲取和利用。
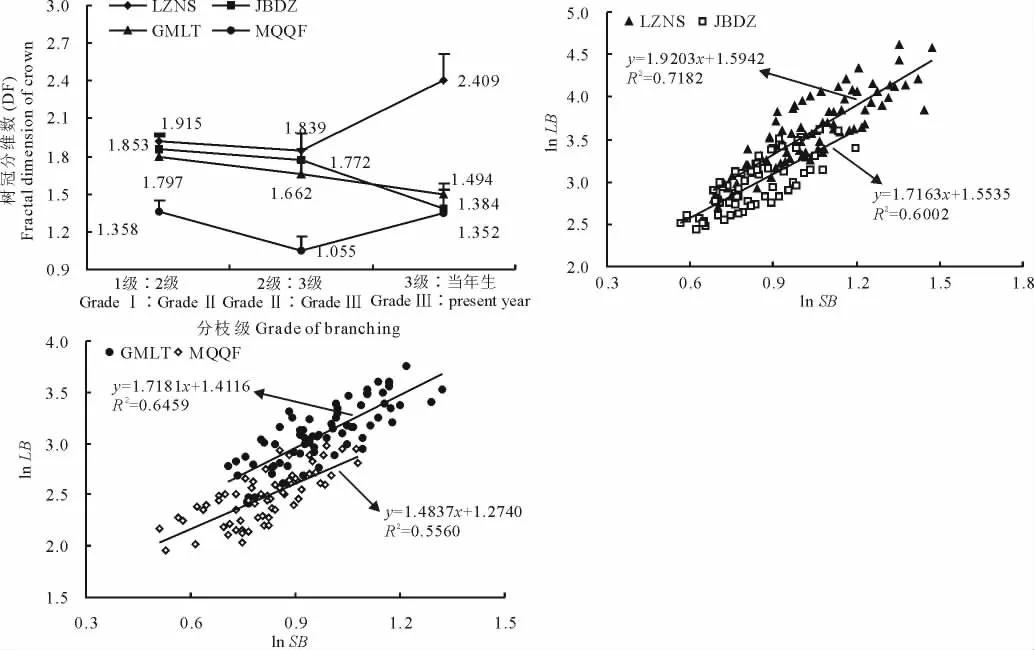
圖3 唐古特白刺克隆植株樹冠分維數(DF)與分枝分維數(DBA)Fig.3 Comparison of fractal dimension of crown(DF)and fractal dimension of branch(DBA)of N.tangutorum clonal populations in 4 plots
白刺無性系枝系構型存在顯著差異,表明各地白刺無性系生長過程均存在有效克隆整合,以調整資源在無性系各部分的分配。有研究表明,分枝角度隨生境可利用性的增強而減小[29],克隆生長型內部調節機制的主要目標是:1)避免克隆內分枝間的競爭;2)最大吸收資源;3)占據最多的空間[30]。我們的研究表明,LZNS白刺無性系的各級分枝角度最小,且與其他三樣地差異性普遍顯著,小的分枝角度可能有助于避免分株太擁擠,進而導致激烈的克隆內競爭和克隆適合性的降低,隨著水分的減少,分株密度減小,分枝角度也呈增大趨勢,這與光的利用有密切關系。分枝強度是與有利生境直接相關的簡單生長反應[31]。在有利的生境中,許多無性系植物的普遍反應是分枝強度提高[26]。我們的研究表明,隨樣地水分梯度遞減,白刺分枝強度逐漸減小并呈現較為顯著的線性規律。
枝徑比、分枝率和分維數也能很好的表征白刺無性系枝系構型的可塑性。LZNS枝徑比普遍較大,而MQQF的3級∶當年生枝徑比較大,這與無性系長期遭受環境脅迫自身克隆整合有關。分枝率在四地差異較顯著,特別是LZNS總體分枝率和逐步分枝率都顯著高于MQQF,說明其生長活力的強大,而MQQF白刺無性系受環境脅迫較強,分枝率偏低。分維數反映植株枝系的復雜程度,LZNS的樹冠分維數與分枝分維數均較大,枝系構件的空間分布趨向復雜化,有利于利用空間資源。而隨著水分條件遞減,無性系為減少枝葉構件間的競爭作用,分維數呈下降趨勢,枝系的空間分布趨于簡單。
研究表明唐古特白刺克隆種群的覓食行為主要體現在,處于水分條件好的生境時,分株間距縮短,分枝角度變小,而分枝強度加大,克隆構型趨于密集型。從而更多分株可能放置于條件好的斑塊下,利于整個基株對資源的獲取。相反,在水分條件差的生境,分株間距增加,分枝角度加大,而分枝強度變小,克隆構型趨于游擊型,利于基株迅速跨越和逃離條件差的斑塊以獲取水分。
生境中的資源狀況脅迫和干擾程度形成選擇壓力,使植物在營養生長、無性生殖和有性生殖中按需分配能量,合成初級代謝產物或次級代謝產物來應對選擇壓力,形成自身的適應對策,最終與生境相互作用過程中表現出的相應的適合度[32]。白刺無性系分株生長格局特征和枝系構型變化反映整個種群生長發育的基本規律和特點,也反映環境信息的變化,是白刺長期適應環境形成的生態對策。本文通過對分株生長格局及枝系構型的分析,探討唐古特白刺無性系生長規律和環境適應機制,有助于進一步認識異質環境下白刺無性系的生存策略。
[1]Van Groenendael J,De Kroon H.Clonal Growth in Plants:Regulation and Function[M].The Hague:SPB Academic Publishing,1990.
[2]Jackson J B C,Buss L W,Cook R C.Population Biology and Evolution of Clonal Organisms[M].New Haven:Yale University Press,1985.
[3]Evans J P.The effect of local resource availability and clonal integration on ramet functional morphology in Hydrocotyle bonariensis[J].Oecologia,1992,89:265-276.
[4]董鳴.資源異質性環境中的植物克隆生長:覓食行為[J].植物學報,1996,38(10):828-835.
[5]李鎮清.克隆植物構型及其對資源異質性的響應[J].植物學報,1999,41(8):893-895.
[6]Slade A J,Hutehings M J.Clonal integration and plasticity in foraging behaviour in Gleehoma hederacea[J].Journal of Ecology,1987,75:1023-1036.
[7]Zhang X S.Evolution and maintenance of the environmental component of the phenotypic variance:benefit of plastic traits under changing environments[J].The American Naturalist,2005,166(5):569-580.
[8]Sultan S E.Phenotypic plasticity for plant development,function and life history[J].Trends in Plant Science,2000,5(12):537-542.
[9]李潔英,解安霞,白昌軍,等.周期性去葉對地毯草克隆生長的影響[J].草業學報,2011,20(3):115-121.
[10]李希來,楊元武,張靜,等.不同退化程度“黑土灘”高山嵩草克隆生長特性[J].草業學報,2003,12(3):51-56.
[11]羅學剛,董鳴.匍匐莖草本蛇莓克隆構型對不同海拔的可塑性反應[J].應用生態學報,2002,13(4):399-402.
[12]馮圖,黎云祥,楊子松,等.不同生境中淫羊藿克隆構型和分株種群特征[J].生態科學,2005,24(4):298-303.
[13]廖明雋,王其兵,董鳴,等.內蒙古錫林河流域不同生境中羊草的克隆構型和分株種群特征[J].植物生態學報,2002,26(1):33-38.
[14]許璐,蘇雪,董莉娜,等.青藏高原東緣中國沙棘的克隆多樣性及克隆結構[J].草業學報,2012,21(3):117-124.
[15]張道遠,王紅玲.荒漠區幾種克隆植物生長構型的初步研究[J].干旱區研究,2005,22(2):219-224.
[16]賈曉紅,李新榮.騰格里沙漠東南緣不同生境白刺(Nitraria)灌叢沙堆的空間分布格局[J].環境科學,2008,29(7):2046-2053.
[17]劉建泉.甘肅民勤西沙窩唐古特白刺群落的生態特性[J].植物資源與環境學報,2002,11(3):36-40.
[18]王尚德,康向陽.唐古特白刺研究現狀與建議[J].植物遺傳資源學報,2005,6(2):231-235.
[19]De Kroon H,Knops J.Habitat exploitation through morphological plasticity in two chalk grassland perennials[J].Oikos,1990,59:39-49.
[20]Oohata S,Shidei T.Studies on the branching structure of trees.1.Bifurcation ratio of trees in Horton’s law[J].Japanese Journal of Ecology,1971,21:7-14.
[21]Whithey G G.The bifurcation ratio as an indicator of adaptive strategy in woody plant species[J].Bulletin of the Torrey Botanical Club,1976,103:67-72.
[22]Steigraeber D A,Waller D M.Non-stationary of Tree Branching Pattern and Bifurcation Ratio[M].London:Proceedings of the Royal Society,1986.
[23]Mandelbrot B B.The Fractal Geometry of Nature[M].SanFrancisco:Freeman,1982.
[24]何明珠,張景光,王輝.荒漠植物枝系構型影響因素分析[J].中國沙漠,2006,26(4):625-630.
[25]董鳴.克隆生長的特性——覓食性[J].生態學報,1996,38(10):828-835.
[26]單保慶,杜國楨,劉振恒.不同養分條件下和不同環境類型中根莖草本黃帚橐吾的克隆生長[J].植物生態學報,2000,24(1):24-51.
[27]王昱生,李景信.羊草種群無性系生長格局的研究[J].植物生態學與地植物學學報,1992,16(3):234-242.
[28]Pitella L F.Application of the-3/2 power law to clonal herbs[J].American Nature,1984,123(4):442-448.
[29]王麗娟,孫棟元,趙成義,等.準噶爾盆地梭梭、白梭梭植物構型特征[J].生態學報,2011,31(17):4952-4960.
[30]Berndtsson R,Chen H.Variability of soil water content along a transect in a desert area[J].Journal of Arid Environments,1994,27(2):127-139.
[31]Hadley N F,Szarek S R.Productivity of desert ecosystem:producer,consumer and decomposer organism[J].Bioscience,1981,31(10):747-753.
[32]Slade A J,Hutchings M J.The effects of nutrient availability on foraging in the clonal herb Glechoma hederacea[J].Journal of Ecology,1987,75(1):95-112.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
