端粒和端粒酶與癌癥發(fā)生和治療的關(guān)系研究進展
2014-01-02 07:57:44陳海偉楊靜茹
赤峰學(xué)院學(xué)報·自然科學(xué)版 2014年19期
關(guān)鍵詞:小鼠
陳海偉,楊靜茹
(赤峰學(xué)院 生命科學(xué)學(xué)院,內(nèi)蒙古 赤峰 024000)
端粒、端粒酶與腫瘤的研究一直以來都是科學(xué)家研究的熱點,在2002年到2004年達到了高峰時期.2009年,美國科學(xué)家伊麗莎白·布萊克本、卡蘿爾·格雷德和杰克·紹斯塔克三人因發(fā)現(xiàn)了端粒和端粒酶保護染色體這一機制獲得了諾貝爾生理學(xué)或醫(yī)學(xué)獎,更加引起了研究者們的關(guān)注[1].
機體受環(huán)境條件或自身的影響會發(fā)生一些遺傳或表觀遺傳信息的改變,這些改變很多情況下會導(dǎo)致癌變,這是由于它們的改變占用了一些信號通路,多條信號通路的綜合改變使有潛在癌變傾向的細胞無限制生長從而導(dǎo)致癌變,進一步擴散,最終導(dǎo)致機體死亡.癌基因是高度重新排列的基因,被賦予了復(fù)雜的轉(zhuǎn)移性,還有局部復(fù)制而導(dǎo)致細胞數(shù)目改變的特征,這些改變是通過與癌癥相關(guān)的基因的改變來實現(xiàn)的.一直以來,人們努力地去尋找造成癌基因不穩(wěn)定的根本機制,最終揭示了在癌癥發(fā)生中具有突出作用的重要結(jié)構(gòu)——端粒,端粒是一種核蛋白結(jié)構(gòu),可以保護真核細胞線性染色體末端,由于真核細胞的染色體復(fù)制機制的原因,其末端在DNA復(fù)制中逐漸縮短,使得依賴細胞分裂的組織的更新令染色體的穩(wěn)定性下降.很多相關(guān)研究證明了端粒縮短對組織衰老和過度增生性疾病的作用,表明端粒的狀態(tài)與癌變風(fēng)險密切相關(guān).在大多數(shù)晚期癌癥患者細胞中,端粒酶被激活進而維持端粒長度,最新的數(shù)據(jù)顯示,端粒酶具有直接控制促癌通路的能力.本文綜述了端粒和端粒酶在正常組織原代細胞中的作用以及在癌癥的發(fā)展中所扮演的角色.
1 概述
1.1 端粒(Telomere)
1.1.1 概念
端粒是真核生物染色體(線性)末端的特殊結(jié)構(gòu),由富含鳥嘌呤G的DNA重復(fù)序列以其相關(guān)蛋白組成,是DNA蛋白復(fù)合物,對染色體結(jié)構(gòu)的穩(wěn)定以及細胞的正常有絲分裂功能的實現(xiàn)有非常重要的意義.
1.1.2 結(jié)構(gòu)
端粒是由富含鳥嘌呤核苷酸的DNA重復(fù)序列構(gòu)成,可作為大型蛋白的結(jié)合位點(見圖1).在脊椎動物中,端粒由重復(fù)序列TTAGGG組成,包括雙鏈部分以及單鏈部分,雙鏈部分較長,單鏈端粒的3′端突出,稱為3′懸突[2].單鏈端粒高度保守,末端可形成一個特殊的環(huán)狀結(jié)構(gòu),稱為t-loop,以此來固定染色體末端,保持其穩(wěn)定性[3].雙鏈的端粒序列可以特異性的結(jié)合雙鏈端粒DNA結(jié)合蛋白,即端粒重復(fù)序列結(jié)合因子1(telomeric repeat binding factor 1,TRF1)和端粒重復(fù)序列結(jié)合因子2(telomeric repeat binding factor 2,TRF2).TRF2在端粒末端保護中起關(guān)鍵的作用,可以促進t-loop環(huán)的形成,是端粒長度的負(fù)性調(diào)節(jié)因子,通過抑制TRF2顯性基因的過表達或者敲除此基因,可造成保護性的帽子結(jié)構(gòu)缺失,表現(xiàn)為3'懸端的處理以及染色體末端的連接反應(yīng)[4,5].TRF1不僅起到了調(diào)節(jié)端粒長度的作用,也可以通過調(diào)節(jié)端粒重復(fù)序列來促進DNA的復(fù)制[6,7,8].TRF1和TRF2專一性的與其他相 關(guān) 蛋白 TIN2(TRF1-interacting nuclear factor)、POT1(protection of telomeres 1)、TPP1和 RAP1(Repressor Activator Protein 1)相互作用,構(gòu)成一個復(fù)合體共同調(diào)節(jié)端粒長度及活性[9].
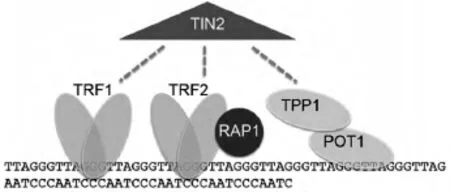
圖1 端粒保護染色體末端
1.2 端粒酶(Telomerase)
1.2.1 概念
端粒酶又稱端粒末端轉(zhuǎn)移酶,其本質(zhì)為核糖核蛋白.人體正常細胞中,端粒隨著細胞有絲分裂代數(shù)的增加而逐漸縮短,每分裂一次約縮短50—200bp,到達臨界水平時,細胞進入衰老期,停止繼續(xù)分裂,但一些特殊細胞,比如絕大多數(shù)的癌細胞和永生化細胞,可通過復(fù)制維持其端粒長度,其催化不是常規(guī)的DNA聚合酶完成,只能由特殊的逆轉(zhuǎn)錄酶也就是端粒酶以自身RNA為模板來完成端粒的延長.
1.2.2 結(jié)構(gòu)
目前對于端粒酶的研究大部分針對動物細胞,常用材料有小鼠、人類等,普遍認(rèn)為,人類端粒酶主要包括三種成分:端粒酶RNA組分(Telomerase RNA Component,TERC)、端粒酶催化亞單位 (Telomere reverse transcriptase,TERT,端粒逆轉(zhuǎn)錄酶)和端粒酶相關(guān)蛋白 (telomerase associated proteinl,TP1).端粒酶是一種大型多亞基RNA酶,TERT和TERC構(gòu)成催化中心[10,11,12,13](見圖2).
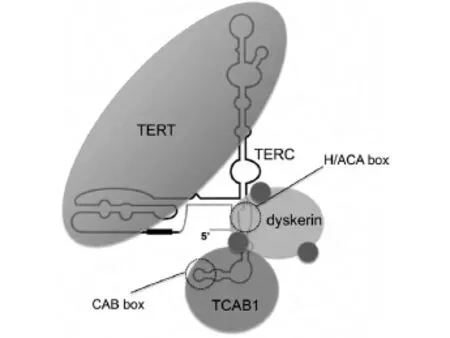
圖2 端粒酶結(jié)構(gòu)
在染色體末端,TERC作為逆轉(zhuǎn)錄模板使端粒重復(fù)序列延長.在體外TERT和TERC就足以使得端粒酶激活,而人體內(nèi),還需要其它酶的共同作用.TERC的5'端包含模板區(qū),3'端包含兩個序列作為其他端粒蛋白的結(jié)合位點.H/ACA box是蛋白dyskerin的結(jié)合位點[14,15,16],dyskerin對于端粒酶的組裝和TERC的穩(wěn)定性相當(dāng)重要.Dyskerin還有三個相關(guān)蛋白,分別為NHP2、NOP10和GAR1.TCAB1是一種WD40重復(fù)序列蛋白質(zhì),在TERC中識別CAB box[17,18].以上兩個蛋白相互作用去調(diào)節(jié)端粒酶的運行進而維持端粒穩(wěn)定性.
1.3 端粒和端粒酶的主要功能及調(diào)節(jié)
1.3.1 端粒和端粒酶的功能
在近年來的研究中,研究者們一直致力于運用各種不同的模式生物進行實驗,試圖闡述端粒的組成結(jié)構(gòu),進而確定了端粒的主要功能.經(jīng)過實驗得出端粒功能主要有:保護染色體的線性末端,防止其降解和不必要的融合;運用其自身的結(jié)構(gòu)能招募端粒酶,用來修復(fù)細胞分裂過程中出現(xiàn)的染色體末端缺失的重復(fù)序列;在細胞核中端粒能定位染色體末端,在減數(shù)分裂過程中這一作用至關(guān)重要,起到確保非錯誤性降解和等量的分裂的作用.相對哺乳動物和酵母的端粒的研究,關(guān)于植物端粒方面的研究還非常有限[19].
1.3.2 端粒酶調(diào)節(jié)端粒長度的機制
多數(shù)人體正常組織或者細胞不會有端粒酶的表達活性,一些有增殖潛能的細胞有低水平表達的端粒酶,如生殖細胞、造血干細胞、免疫記憶細胞、皮膚細胞等.以自身的RNA為模版,端粒酶能夠逆轉(zhuǎn)錄合成具有DNA重復(fù)序列的端粒,從而使得端粒延長,保持染色體結(jié)構(gòu)的相對恒定.
在端粒酶各個亞單位相互協(xié)作下可十分精確地調(diào)控端粒延伸的過程.首先,端粒酶逆轉(zhuǎn)錄酶TERT被激活,進而使端粒酶活化;其次,通過結(jié)合蛋白端粒酶結(jié)合于端粒,以端粒末端做為引物,與端粒酶RNA模板區(qū)的起始端結(jié)合,開始端粒DNA的合成,DNA鏈延長至模板區(qū)的末端時,延長的DNA末端與模板解開,新合成的端粒DNA仍然結(jié)合在端粒酶結(jié)合蛋白上,其末端又結(jié)合在端粒酶RNA模板區(qū)的起始位,如此反復(fù)進行,直至端粒達到適當(dāng)長度時終止[20].
1.3.3 端粒酶活性的調(diào)節(jié)
對于端粒酶活性的調(diào)節(jié)方式,一般分為兩種,一種是通過誘導(dǎo)端粒酶使其活性增強,目前是通過利用攜帶hTERT基因的載體轉(zhuǎn)染干細胞的研究方法,從而激活端粒酶,使被轉(zhuǎn)染的干細胞的端粒酶活性升高;另外一種是抑制端粒酶的活性,使端粒酶低水平的表達,甚至表達停止.研究發(fā)現(xiàn),80%甚至以上的腫瘤細胞的端粒酶都處于活化狀態(tài).
在正常組織細胞中(包括正常干細胞),端粒酶只是暫時性表達并且表達水平很低,同腫瘤細胞相,比其端粒比較長,腫瘤細胞的端粒酶的活化是腫瘤發(fā)生的一個重要步驟,且與腫瘤細胞的增殖、分化、轉(zhuǎn)移、凋亡以及細胞周期有關(guān),這就使人們把研究癌癥治療藥物的眼光落到抑制端粒酶活性上,使其實現(xiàn)成為可能.目前對于該類藥物的研究方向主要有酶直接抑制劑、端粒酶主動免疫治療和干擾端粒酶反轉(zhuǎn)錄過程等幾個方向[21].
1.4 端粒酶的檢測技術(shù)
隨著端粒酶的結(jié)構(gòu)、功能及調(diào)控的進一步研究,對端粒酶在細胞永生化以及腫瘤癌變進程中的作用有了嶄新的認(rèn)識,因此對端粒酶的檢測方法的研究成為研究端粒酶至關(guān)重要的部分.現(xiàn)在主要的技術(shù)有:端粒重復(fù)序列延伸法(TEA),端粒重復(fù)序列擴增法(TRAP),半定量TRAP法,細針抽吸端粒酶重復(fù)擴增法(FNA—TRAP),銀染端粒重復(fù)序列擴增法(SS—TRAP),接近閃爍分析端粒重復(fù)序列擴增法,酶聯(lián)免疫吸附端粒重復(fù)序列擴增法(TRAP—ELISA),原位TRAP法應(yīng),定量端粒重復(fù)序列擴增(RTQ—TRAP)[22].
1.5 端粒異常是原代細胞缺陷的關(guān)鍵驅(qū)動者
通過敲除TERC基因組分的端粒酶的小鼠模型,研究端粒逐漸縮短引起的端粒危機對生物體的影響.出乎意料的是,前幾代均表現(xiàn)正常,從而一段時期內(nèi)科學(xué)家輕率的認(rèn)為端粒酶組分對于生命是可有可無的[23].后來發(fā)現(xiàn)這些小鼠的存活是因?qū)嶒炐∈笃废涤休^長的端粒,然而,第一代(G1)TERC基因敲除純合體小鼠的持續(xù)繁殖產(chǎn)生了新生的TERC基因敲除純合體小鼠,導(dǎo)致端粒逐步喪失,最終引起了高度增生的嚴(yán)重組織變性,第4代出現(xiàn)生育能力下降,到G5–G6TERC基因敲除純合體小鼠出現(xiàn)不育現(xiàn)象,在睪丸組織中表現(xiàn)出較高的細胞凋亡和生殖細胞耗竭,出現(xiàn)骨髓造血細胞功能受損以及淋巴細胞增殖減弱.在G5–G6TERC基因敲除純合體組織中,端粒逐漸縮短的細胞遺傳學(xué)證據(jù)很清晰,包括染色體末端融合以及有絲分裂后期的染色體橋的出現(xiàn).說明這些病癥以及其嚴(yán)重程度是與端粒功能失調(diào)的程度緊密聯(lián)系的.
在較晚世代的端粒酶基因敲除的小鼠中,嚴(yán)重退行性的組織缺損似乎與增殖損傷、細胞凋亡加強的原代細胞以及干細胞中自我更新缺陷有關(guān)聯(lián).這些缺陷是顯而易見的,即原代細胞凋亡率較高,造成腸道腺細胞絨毛萎縮,在端粒酶缺失的小鼠中造血干細胞潛在減少.與野生型相比,經(jīng)連續(xù)輻射的G1TERT基因敲除純合體或者G1TERC基因敲除純合體小鼠的造血干細胞造血能力下降,這種缺陷在較晚世代TERT基因敲除純合體小鼠造血干細胞中更容易出現(xiàn),具有較短端粒的較晚世代的TERT基因敲除純合體小鼠與長端粒的野生型小鼠雜交產(chǎn)生的TERT基因敲除雜合體小鼠擁有一半正常的端粒和一半功能失調(diào)的短端粒,這些小鼠顯現(xiàn)出與原來較晚世代TERT基因敲除純合體小鼠相似的細胞凋亡和增殖缺陷,所以研究者一直認(rèn)為系列的端粒功能失調(diào)是原代細胞出現(xiàn)缺陷的關(guān)鍵驅(qū)動者,更加突出了端粒在細胞中的重要地位.
2 端粒、端粒酶與癌癥的關(guān)系
2.1 正常細胞和癌細胞復(fù)制中端粒的差異性
在人類成纖維細胞原代培養(yǎng)實驗中發(fā)現(xiàn)了端粒與癌細胞之間的聯(lián)系,正常細胞表現(xiàn)出有限的復(fù)制能力,原代細胞一般分裂60–80代就進入衰老狀態(tài),直至死去,與此相反,癌細胞系在細胞培養(yǎng)時可以無限制的生長下去 (見圖3).正常培養(yǎng)的人類成纖維細胞在分裂期間端粒是逐漸縮短的,這些縮短的端粒激活了細胞衰老程序,但是在腫瘤細胞中端粒卻維持原始狀態(tài)并不縮短.隨著人成纖維細胞繼續(xù)分裂,端粒逐漸縮短,端粒帽子結(jié)構(gòu)功能損壞,導(dǎo)致染色體極度不穩(wěn)定,最終細胞凋亡,這種狀態(tài)稱為危機.但是有少數(shù)細胞則可以通過一定的機制延長端粒而成為不受調(diào)控的細胞,比如人類成纖維細胞的衰老以及細胞出現(xiàn)的危機,可以通過重激活端粒酶進而合成端粒重復(fù)序列有效的化解,大部分癌癥在發(fā)生時通過重新激活端粒酶的活性以維持端粒長度.
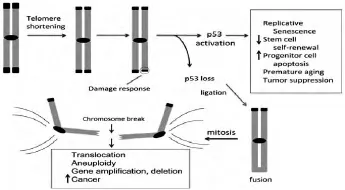
圖3 端粒縮短激活p53并且通過基因擴增和缺失驅(qū)動上皮癌形成
2.2 端粒酶結(jié)構(gòu)中TERT對其活性起關(guān)鍵作用
有學(xué)者研究了不同病變的胃黏膜端粒酶活性并對其亞單位進行檢測,結(jié)果發(fā)現(xiàn)端粒酶活性均有不同程度的表達,而正常胃黏膜中則沒有端粒酶的表達,且癌細胞中表達率最高.TERT主要在癌組織或部分癌前組織中表達,而其他兩個亞基在所有情況下均有表達,TERT在癌組織中的表達與端粒酶活性有明顯的相關(guān)性,由此提示在胃癌的發(fā)生過程中端粒酶可能具有極其重要的作用;可把TERT作為胃癌的診斷指標(biāo),同時也可以把TERT作為胃癌基因治療的靶位點.張營等研究了在肝癌中端粒酶組分的表達情況發(fā)現(xiàn),hTERTmRNA陽性表達普遍存在于肝癌組織中,說明了通過檢測肝癌中hTERTmRNA表達情況可用來作為肝癌診斷及預(yù)后估計的新標(biāo)志物.總之,這些研究結(jié)果說明對于端粒酶來說,其三種組分是缺一不可的,但是TERT只在有端粒酶活性的細胞中表達,相較于端粒酶其它兩個組分來說,它的上調(diào)可能與端粒酶活性的增強有很大聯(lián)系,從而穩(wěn)定端粒長度,支持細胞無限的分裂.
2.3 端粒和端粒酶與P53的關(guān)系
2.3.1 端粒與P53蛋白通路及細胞衰老
端粒極度縮短誘導(dǎo)細胞和組織的改變,這個變化主要集中在p53抑癌蛋白這一細胞壓力傳感器上,它可以對癌基因的激活、DNA損傷信號和缺氧等各種刺激做出反應(yīng),起到強制細胞周期停止或使細胞凋亡的作用.實驗證明,在端粒酶基因敲除的小鼠中,p53蛋白是穩(wěn)定的,并且p53缺失顯著削弱細胞周期阻滯、降低了細胞死亡率.在人工培養(yǎng)細胞的中,通過破壞端粒結(jié)合蛋白TRF2使得端粒帽子結(jié)構(gòu)損壞,進而強烈的激活p53基因.以人類成纖維細胞中的端粒為例,端粒TRF2的功能被打亂,出現(xiàn)一些導(dǎo)致染色體內(nèi)部雙鏈斷裂的典型DNA損傷蛋白,顯示的病灶有組蛋白變體γH2AX、DNA損傷蛋白p53BP1、Mre11復(fù)雜化以及磷酸共濟失調(diào)毛細血管擴張癥突變(ataxia telangiectasia mutated,ATM)等,促進細胞衰亡.在端粒酶基因敲除小鼠繁殖的較晚世代中,表現(xiàn)出早衰綜合癥,包括毛發(fā)變得灰白無光澤,生存期縮短等不良反應(yīng),說明端粒縮短可能會引起許多老化現(xiàn)象,是重要的遺傳毒性信號,這個信號可以激活DNA損傷信號,從而激活p53基因,加速細胞衰老進程;活性氧可以引起DNA損傷,進一步加強p53對DNA損傷檢查點的反應(yīng),進一步加速端粒的侵蝕.
總之,這些數(shù)據(jù)證明在衰老的進程中,依賴于p53基因的DNA損傷信號的重要性,端粒功能失調(diào)會引起p53基因的表達,進而加速異常組織細胞的衰老或凋亡.
2.3.2 端粒縮短抑制腫瘤發(fā)生
為確定端粒耗損是否影響癌變過程,將TERC基因敲除純合體與p53基因敲除純合體小鼠進行雜交研究,結(jié)果發(fā)現(xiàn),在早期階段,端粒功能異常給細胞以及器官帶來的不利影響可被p53基因敲除明顯減弱.同代TREC和p53雙基因敲除純合體小鼠與只有TREC基因敲除純合體的小鼠相比,前者存活時間長于后者,多數(shù)G5-G6 TREC和p53雙基因敲除純合體純合體小鼠睪丸組織的凋亡被減弱,G6TREC和p53雙基因敲除純合體小鼠睪丸大小要比G6只有TREC基因敲除純合體的小鼠的正常.由此說明,在TREC基因敲除純合體小鼠中,抑制細胞生長和凋亡的反應(yīng)可在p53的功能喪失的情況下完全喪失,導(dǎo)致器官衰亡消弱,與端粒相關(guān)的組織退化得到改善.雖然對G6TREC和p53雙基因敲除純合體小鼠與G6唯TREC基因敲除純合體小鼠的睪丸進行比較發(fā)現(xiàn)后者更接近正常,但G6TREC和p53雙基因敲除純合體小鼠中約有一半小鼠的生殖小管表現(xiàn)出不同程度的萎縮,而且在G7-G8雙基因敲除純合體的小鼠中重新出現(xiàn)萎縮,由此表明小鼠體內(nèi)還存在一些不依賴于p53信號介導(dǎo)的端粒功能異常反應(yīng).
Ink4a/Arf基因編碼兩個不同的腫瘤抑制蛋白:細胞周期蛋白依賴性激酶抑制劑p16和p53激活因子p19ARF,因子p19ARF對p53蛋白調(diào)節(jié)主要是通過阻斷p53的降解,檢測異常的細胞周期,致使p53穩(wěn)定.Ink4a/Arf突變小鼠長出淋巴瘤與肉瘤,具有短潛伏期和高外顯率;但是,在G4/G5TERC-/-Ink4a/Arf-/-的小鼠顯示出降低腫瘤的發(fā)病率和增長潛伏期的功效,腫瘤的范圍沒有變化,顯示出p53突變的組織中,端粒缺失對腫瘤有抑制作用,說明了在腫瘤發(fā)生過程中的依賴p53的端粒檢查點的反應(yīng)以及端粒維持的重要性.
以上實驗表明,在腫瘤發(fā)生過程中依賴p53的端粒檢查點的反應(yīng)以及端粒維持的重要性,說明端粒缺陷會引起依賴p53的DNA損傷修復(fù)促進細胞衰亡,防止癌變的發(fā)生,而在p53缺陷可以顯著削弱端粒失調(diào)所帶來的細胞衰亡等不利現(xiàn)象.
3 端粒酶抑制劑——癌癥抑制物質(zhì)的研究成果
基于端粒酶在癌癥中的重要作用,無數(shù)的研究者將端粒酶作為癌癥的治療點,對端粒酶抑制劑的研究,主要是有以下途徑:(1)利用反義 hTR(human telomerase RNA,hTR)、反義寡核苷酸(Antisense oligonucleotides,ODN)、肽苷酸(peptide nucleic acid,PNAS)、錘頭狀核酶 (Hammerhead Ribozyme)等手段抑制端粒酶RNA的模板作用,進而抑制端粒酶活性.(2)對端粒酶蛋白組分的抑制劑:端粒酶蛋白抗體;蛋白激酶C(protein kinase C,PKC)調(diào)節(jié)劑;TERT抑制劑.(3)其他抑制劑.
在對治療癌癥的研究過程中發(fā)現(xiàn)了許多植物提取物對端粒酶有一定的作用,比如:目前已證實的植物生物堿傳統(tǒng)中藥苦參的主要生物堿之一苦參堿、喜樹堿、小檗堿、長春堿、苦參堿、秋水仙堿、三尖杉酯堿等,小檗屬植物的一種具有生物活性的成分小檗胺,白頭翁根中分離提取的樺木酸衍生物2、3-羥基樺木酸,紅紋馬先蒿中所含的苯丙素苷類化臺物毛蕊花苷,從我國和印度的傳統(tǒng)民間草藥珠子草的乙酸乙酯提取物中純化得到的亞甲基雙氧木質(zhì)素,茶葉的主要活性成分茶多酚中的兒茶素類物質(zhì),從中國云南省所產(chǎn)的豆科藤本植物相思子的種子中分離得到的一種新的單體蛋白相思子蛋白P2,均有抑制端粒酶活性的作用.因而也可以作為癌癥抑制的有效物質(zhì),但是仍然需要在臨床中不斷實驗.
4 結(jié)論和展望
從一系列人類系統(tǒng)和小鼠模型的研究表明,端粒酶對調(diào)節(jié)端粒的長度有重要作用,端粒功能的完好對于細胞增殖是至關(guān)重要的.在端粒酶中,TERT是關(guān)鍵的亞基,在端粒酶活性中起主要作用.端粒功能失調(diào)在原代細胞出現(xiàn)缺陷中扮演著重要的角色.端粒、端粒酶通過與p53等抑癌基因的相互作用,證明了其在細胞衰亡和癌變中的關(guān)鍵地位,并激起了人們對端粒酶抑制劑的研究興趣.
端粒與端粒酶對癌癥的重要性已被確定,但是更具體的作用機制仍然不清晰,端粒酶抑制劑的研究大部分也處于實驗階段,繼續(xù)端粒與端粒酶這個領(lǐng)域的研究將會得到更新、更重要的成果,例如它們?nèi)绾握{(diào)節(jié)原代細胞、組織、有機體的年齡以及癌基因是如何發(fā)展等方面探索.
〔1〕孔令平,汪華僑.端粒和端粒酶與衰老、癌癥的潛在關(guān)系——2009年諾貝爾生理學(xué)或醫(yī)學(xué)獎簡介[J].自然雜志,第31卷.第6期.
〔2〕Palm,W.et al.(2008)How shelterin protects mammalian telomeres.Annu.Rev.Genet.,42,301–334.
〔3〕Griffith,J.D.et al.(1999)Mammalian telomeres end in a large duplex loop.Cell,97,503–514.
〔4〕van Steensel,B.et al.(1998)TRF2 protects human telomeres from end-toend fusions.Cell,92,401–413.
〔5〕Celli,G.B.et al.(2005)DNA processing is not required for ATMmediated telomere damage response after TRF2 deletion.Nat.Cell Biol.,7,712–718.
〔6〕van Steensel,B.et al.(1997)Control of telomere length by the human telomeric protein TRF1.Nature,385,740–743.
〔7〕Smogorzewska,A.et al.(2000)Control of human telomere length by TRF1 and TRF2.Mol.Cell.Biol.,20,1659–1668.
〔8〕Sfeir,A.et al.(2009)Mammalian telomeres resemble fragile sites and require TRF1 for efficient replication.Cell,138,90–103.
〔9〕de Lange,T.(2005)Shelterin:the protein complex that shapes and safeguards human telomeres.Genes Dev.,19,2100–2110.
〔10〕Greider,C.W.et al.(1989)A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis.Nature,337,331–337.
〔11〕Lingner,J.et al.(1997)Reverse transcriptase motifs in the catalytic subunit of telomerase.Science,276,561-567.
〔12〕Nakamura,T.M.et al.(1997)Telomerase catalytic subunit homologs from fission yeast and human.Science,277,955–959.
〔13〕Meyerson,M.et al.(1997)hEST2,the putative human telomerase catalytic subunit gene,isup-regulated in tumor cellsand during immortalization.Cell,90,785-795.
〔14〕Mitchell,J.R.et al.(1999)A telomerase component is defective in the human disease dyskeratosis congenita.Nature,402,551–555.
〔15〕Cohen,S.B.et al.(2007)Protein composition of catalytically active human telomerase from immortal cells.Science,315,1850–1853.
〔16〕Venteicher,A.S.et al.(2008)Identification of ATPases pontin and reptin as telomerase components essential for holoenzyme assembly.Cell,132,945–957.
〔17〕Venteicher,A.S.et al.(2009)A human telomerase holoenzyme protein required for Cajal body localization and telomere synthesis.Science,323,644–648.
〔18〕Tycowski,K.T.et al.(2009)A conserved WD40 protein binds the Cajal body localization signal of scaRNP particles.Mol.Cell,34,47–57.
〔19〕劉由頁,宋涵,李鳳蘭,等.植物端粒與端粒酶研究進展[J].北京林業(yè)大學(xué)學(xué)報,2012(5).
〔20〕王聯(lián)群,劉德伍.端粒、端粒酶與干細胞[J].中國組織工程研究與臨床康復(fù),2009(10).
〔21〕惠子健.端粒酶的研究進展[J].山西職工醫(yī)學(xué)院學(xué)報,2010(1).
〔22〕洪日,陳紅風(fēng).端粒酶檢測技術(shù)新進展[J].臨床和實驗醫(yī)學(xué)雜志,2007(11):163-164.
〔23〕Blasco,M.A.et al.(1997)Telomere shortening and tumor formation by mouse cells lacking telomerase RNA.Cell,91,25–34.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(2014年3期)2014-07-31 18:57:34
 赤峰學(xué)院學(xué)報·自然科學(xué)版2014年19期
赤峰學(xué)院學(xué)報·自然科學(xué)版2014年19期
- 赤峰學(xué)院學(xué)報·自然科學(xué)版的其它文章
- 一類解析函數(shù)的系數(shù)不等式和卷積性質(zhì)
- 地方高校本科生物學(xué)實驗體系構(gòu)建的研究
- 計算機輔助教學(xué)在《數(shù)學(xué)建模》課程中的運用
——赤峰學(xué)院《數(shù)學(xué)建模》教學(xué)心得體會 - 關(guān)于數(shù)字電子技術(shù)基礎(chǔ)課程中寄存器教學(xué)的探討
- 關(guān)于數(shù)字電子技術(shù)基礎(chǔ)課程教學(xué)改革的探討
- 應(yīng)用統(tǒng)計類專業(yè)探究性教學(xué)模式探索與實踐
——以變量選擇準(zhǔn)則為例