高活性谷氨酰胺酶基因glsA2在枯草芽孢桿菌BJ3-2染色體上的定點整合
2014-01-17 11:38:30吳擁軍劉艷敏王亞娟
食品科學(xué) 2014年1期
盧 彪,吳擁軍*,劉艷敏,王亞娟,唐 雪
(貴州大學(xué)生命科學(xué)學(xué)院,貴州 貴陽 550025)
高活性谷氨酰胺酶基因glsA2在枯草芽孢桿菌BJ3-2染色體上的定點整合
盧 彪,吳擁軍*,劉艷敏,王亞娟,唐 雪
(貴州大學(xué)生命科學(xué)學(xué)院,貴州 貴陽 550025)
以枯草芽孢桿菌BJ3-2的glsA1基因為同源序列,通過構(gòu)建單交換整合載體,將高活性谷氨酰胺酶基因glsA2定點整合入BJ3-2菌株染色體中,獲得能夠穩(wěn)定遺傳的重組菌株BJ3-2A2。經(jīng)檢測重組菌株谷氨酰胺酶活力為BJ3-2菌株的3.36倍。利用重組菌株BJ3-2A2發(fā)酵豆豉,全氨基酸檢測顯示,重組菌株發(fā)酵的豆豉谷氨酸含量比出發(fā)菌株高12.8%。說明枯草芽孢桿菌BJ3-2可以轉(zhuǎn)化且為RecE+菌株,glsA2基因在BJ3-2菌株染色體上可實現(xiàn)較高活性表達,并提高發(fā)酵豆豉的鮮味。
枯草芽孢桿菌;同源重組;單交換;谷氨酰胺酶基因
枯草芽孢桿菌是革蘭氏陽性細菌,具有非致病性、分泌蛋白能力強的特性和良好的發(fā)酵基礎(chǔ),是美國食品藥物管理局(Food and Drug Administration,F(xiàn)DA)和中國農(nóng)業(yè)部批準使用的安全菌株,廣泛應(yīng)用于食品發(fā)酵行業(yè)[1-2]。
枯草芽孢桿菌中的質(zhì)粒在復(fù)制時經(jīng)常出現(xiàn)不穩(wěn)定單鏈(ssDNA)形式,導(dǎo)致質(zhì)粒丟失[3],因此對于枯草芽孢桿菌的基因改造主要是針對染色體基因的改造。而枯草芽孢桿菌中染色體同源序列整合大多發(fā)生在RecE+枯草芽孢桿菌內(nèi),RecE蛋白與大腸桿菌中的RecA酶類似,是一種用以重組修復(fù)及基因交換的重組酶[4]。
枯草芽孢桿菌BJ3-2為本研究室從貴州水豆豉中分離得到的一株發(fā)酵風(fēng)味較好,可作為工業(yè)生產(chǎn)應(yīng)用的豆豉發(fā)酵菌株[5]。實驗將前期從枯草芽孢桿菌GA317中克隆得到的高活性谷氨酰胺酶基因glsA2[6]以單交換的方式定點整合入BJ3-2菌株獲得重組菌株BJ3-2A2,進而對BJ3-2菌株是否為可轉(zhuǎn)化菌株、RecE+菌株;glsA2基因整合入BJ3-2菌株染色體后能否存在活性;以及重組菌株的產(chǎn)谷氨酰胺酶能力等問題進行驗證,為今后通過雙交換重組或RED重組食品級置換BJ3-2菌株中的glsA基因[6]提供參考。
1 材料與方法
1.1 菌株與質(zhì)粒
B.subtilis BJ3-2、E.coli DH5α 本實驗室保存。
質(zhì)粒pMD18、pMD18-glsA1、pMD18-glsA2、pET28b本實驗室保存。
1.2 試劑與儀器
基因組提取試劑盒 美國Promega公司;質(zhì)粒提取試劑盒、DNA純化試劑盒 美國Omega公司;Bacterium Glutaminase Assay Kit 美國Genmed公司;Taq酶、限制性內(nèi)切酶、T4 DNA連接酶、DNA Marker日本TaKaRa公司;其他試劑均為國產(chǎn)分析純。
MyLycler PCR儀、Universal Hood凝膠成像分析儀、Gene Pulser Xcell 美國Bio-Rad公司;L8800全自動高速氨基酸分析儀 日本日立公司。
1.3 方法
1.3.1 常規(guī)方法[7-8]
質(zhì)粒提取、B.subtilis基因組提取、采用試劑盒方法進行;DNA酶切、片段回收、連接采用常規(guī)方法。
1.3.2 PCR引物設(shè)計
按glsA2基因序列設(shè)計引物(下劃線均為酶切位點,下同),Pgsa-F:CGCTGCAGATGCA GTGCATTG AAACAAAC (PstⅠ);Pgsa-R:CGAAGCTTCGGGCA TGCATGCTCCAACCTTCTTG (Hind Ⅲ)。
按pMD18載體上Amp基因及glsA2基因序列設(shè)計引物,Pamp-F:GCAAGCTTATGAGTATTCAACATTTC(Hind Ⅲ);Pgsa-R:CGAAGCTTCGGGCATGCATGC TCCAACCTTCTTG (Hind Ⅲ)。
按pET28b載體上kan基因序列設(shè)計引物,Pkan-F:CGGGGTACCCATGAGCCATATTCAACGGG (KpnⅠ);Pkan-R:GCTCTAGATTAGAAAAACTCATCGAGCATC(XbaⅠ)。
1.3.3 含glsA2基因的單交換整合載體的構(gòu)建
以pMD18為骨架,分別克隆入glsA1、glsA2、kan這3個基因,其中g(shù)lsA2為目的基因;glsA1為B.subtilis BJ3-2的同源序列;kan為重組菌株BJ3-2A2抗性篩選標記。構(gòu)建過程見圖1。以pMD18-glsA2質(zhì)粒為模板,利用引物Pgsa-F/Pgsa-R擴增得到含雙酶切位點PstⅠ/HindⅢ的glsA2基因,并克隆入pMD18-glsA1質(zhì)粒,獲得質(zhì)粒pMD18-glsA1-glsA2并轉(zhuǎn)化E.coli DH5α;以pMD18-glsA1-glsA2質(zhì)粒為模板,利用引物Pamp-F/Pgsa-R擴增得到首尾均帶HindⅢ酶切位點的、去除部分無關(guān)序列的簡化質(zhì)粒pMD18’-glsA1-glsA2片段,將pMD18’-glsA1-glsA2片段連接環(huán)化為pMD18’-glsA1-glsA2質(zhì)粒并轉(zhuǎn)化E.coli DH5α;以pET28b質(zhì)粒為模板,利用引物Pkan-F/Pkan-R擴增得到含雙酶切位點KpnⅠ/XbaⅠ的kan基因,克隆入pMD18’-gls A1-glsA2質(zhì)粒,獲得單交換重組質(zhì)粒pTKGG并轉(zhuǎn)化E.coli DH5α。
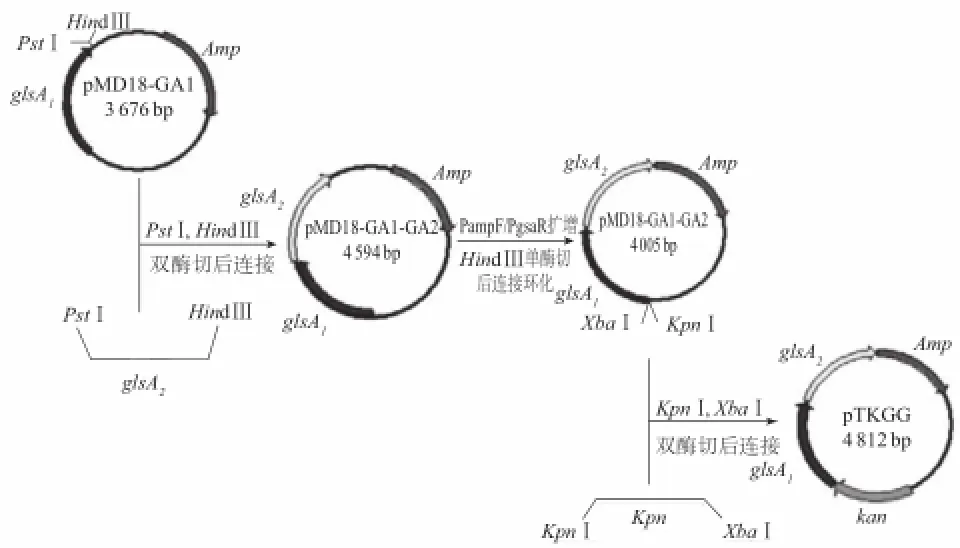
圖1 單交換載體構(gòu)建過程示意圖Fig.1 Diagram showing the construction process of a single exchange vector pTKGG
1.3.4 電轉(zhuǎn)BJ3-2菌株及重組菌株的篩選
BJ3-2菌株感受態(tài)的制備參照文獻[9]。電轉(zhuǎn)緩沖液為272mmol/L蔗糖與10%甘油的去離子水溶液。質(zhì)粒加入量約1μg,電轉(zhuǎn)條件為2.5kV,200Ω,25μF,2mm[10-11]。電擊后感受態(tài)細胞轉(zhuǎn)移至0.8 mL LB液體培養(yǎng)基中,37℃、180 r/min振蕩1 h,菌體涂布于LB固體平板上(含卡那霉素2 μg/mL),37℃培養(yǎng)過夜[12]。挑取重組子進行PCR鑒定,獲得陽性重組菌株BJ3-2A2。
由于pMD18質(zhì)粒上所帶復(fù)制子只能被大腸桿菌識別,而不能被枯草芽孢桿菌(革蘭氏陽性菌)識別[13],致使單交換載體pTKGG在BJ3-2菌株中無法復(fù)制。因此,單交換重組質(zhì)粒pTKGG通過電轉(zhuǎn)進入BJ3-2菌株中后,即通過同源重組整合到BJ3-2菌株染色體上,重組菌株通過傳代丟失單交換重組載體pTKGG。
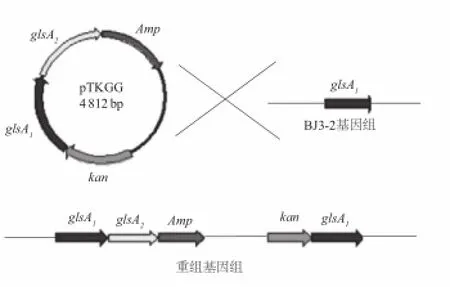
圖2 重組菌株通過傳代丟失單交換整合重組載體示意圖Fig.2 Diagram showing the integration of single exchange
圖2為pTKGG載體整合入BJ3-2菌株的示意圖[14],BJ3-2菌株在glsA1基因后將整個載體整合入染色體。因此可通過PCR檢測glsA2和kan基因來鑒定陽性重組菌。
1.3.5 傳代培養(yǎng)重組菌株BJ3-2A2檢測重組基因的穩(wěn)定性
以體積分數(shù)0.1%的接種量將重組菌株BJ3-2A2接種LB液體培養(yǎng)基連續(xù)傳代20次后利用PCR檢測基因穩(wěn)定性。
1.3.6 重組菌株BJ3-2A2的谷氨酰胺酶酶活力測定
采用Bacterium Glutaminase Assay Kit檢測,基于底物谷氨酰胺在谷氨酰胺酶的作用下,轉(zhuǎn)化為谷氨酸和氨氣,進而在谷氨酰胺脫氫酶的催化下,伴隨著氧化型煙酰胺腺嘌呤二核酸(NAD+)轉(zhuǎn)化為還原型煙酰胺腺嘌呤二核苷酸(NADH),產(chǎn)生吸收峰值的變化(340 nm波長),來定量分析谷氨酰胺酶的活性。酶活力單位定義:在特定條件下,1 min內(nèi)轉(zhuǎn)化1 μmol底物所需的酶量,為一個酶活力單位,1 U。
1.3.7 發(fā)酵豆豉的氨基酸組分及含量檢測
利用BJ3-2菌株及重組菌株BJ3-2A2進行豆豉發(fā)酵。發(fā)酵豆豉研缽研磨,參照GB/T 5009.124—2003《食品中氨基酸的測定》,采用全自動高速氨基酸分析儀上樣檢測,并驗證差異性。
2 結(jié)果與分析
2.1 單交換重組質(zhì)粒pTKGG的構(gòu)建及單交換重組菌株的檢測
擴增模板為連續(xù)傳代20次后的BJ3-2A2菌株基因組。BJ3-2A2菌株經(jīng)質(zhì)粒提取驗證無質(zhì)粒存在,圖3顯示,PCR擴增glsA1、glsA2、kan基因片段大小分別約為984、924 bp和816 bp,均與預(yù)期片段大小一致。因此,glsA2、kan基因均已整合入菌株染色體上,該菌為單交換陽性重組菌。
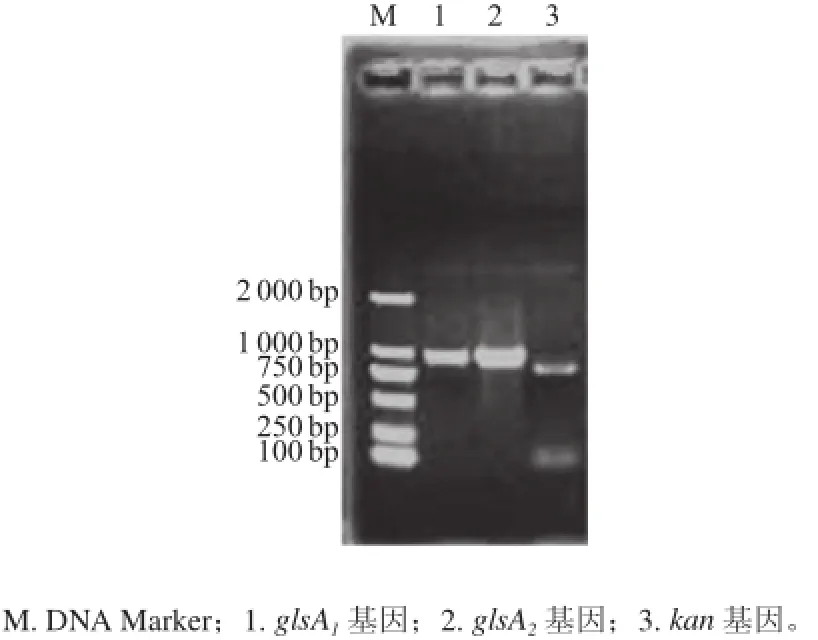
圖3 重組菌株基因組ggllssAA1、ggllssAA2、kkaann基因的PPCCRR擴增Fig.3 PCR amplification of the genes ggllssAA1, ggllssAA2and kkaann from recombinant strain
2.2 整合基因穩(wěn)定性檢測
重組菌株BJ3-2A2連續(xù)傳代20次后,利用PCR檢測菌株中g(shù)lsA2基因的穩(wěn)定性。由圖4可知,轉(zhuǎn)接培養(yǎng)20代后進行檢測的10個菌落全部為陽性,表明glsA2基因在BJ3-2A2菌株中能夠穩(wěn)定遺傳。
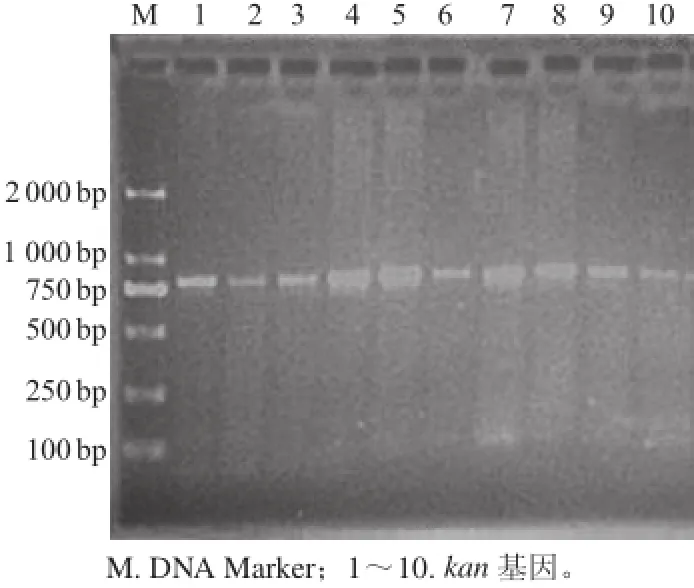
圖4 重組菌株基因組kkaann基因的PPCCRR擴增Fig.4 PCR amplification of the gene kan from recombinant strain
2.3 重組菌株BJ3-2A2谷氨酰胺酶活力
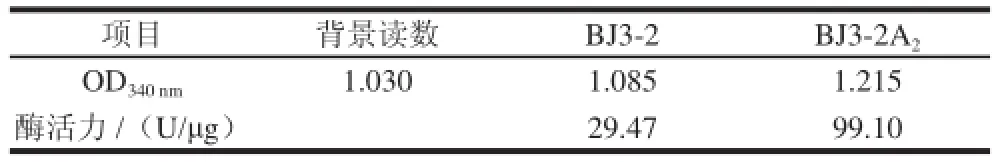
表1 重組菌株BJ3--22AA2酶活力比較Table 1 Comparison of enzymatic activities between BJ3-2 and BJ3-2A
對相同培養(yǎng)條件下的BJ3-2菌株及BJ3-2A2菌株利用Bacterium Glutaminase Assay Kit檢測谷氨酰胺酶酶活力,結(jié)果見表1,兩株菌的酶活力分別為29.47、99.10 U/?g,重組菌株酶活力是BJ3-2菌株的3.36倍。
2.4 發(fā)酵豆豉氨基酸含量
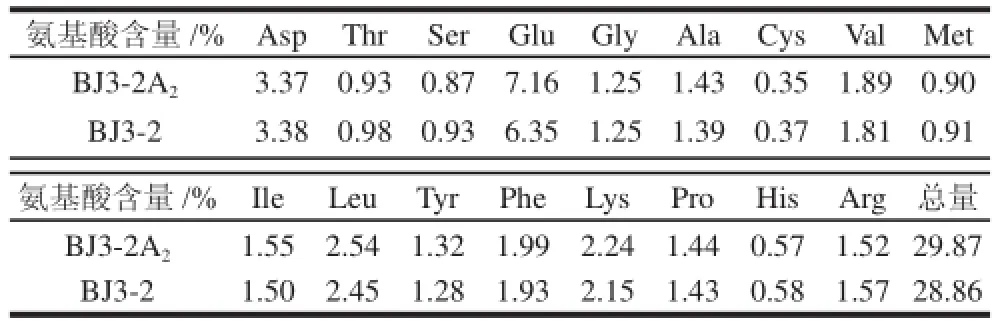
表2 豆豉氨基酸含量對比Table 2 Comparison of amino acid composition and content in douchi fermented with different strains
對BJ3-2A2、BJ3-2的豆豉發(fā)酵產(chǎn)物分別進行氨基酸含量分析,由表2可知,兩種發(fā)酵豆豉的17種氨基酸中僅谷氨酸含量有較為明顯的變化(P<0.05),BJ3-2A2菌株比BJ3-2菌株高出12.8%。分析原因與整合的glsA2基因有關(guān),谷氨酰胺酶活性提高,使谷氨酰胺轉(zhuǎn)化為谷氨酸,進而得到較高含量谷氨酸。
3 結(jié)論與討論
由于遺傳背景的原因,現(xiàn)研究多是對遺傳背景清楚、確定可轉(zhuǎn)化且具RecE+的枯草芽孢桿菌標準株(如枯草芽孢桿菌168)進行研究[15-17],而對于其他野生型枯草芽孢桿菌的分子改造研究較少。本實驗對野生型并已作為工業(yè)生產(chǎn)應(yīng)用的枯草芽孢桿菌BJ3-2進行染色體整合,證實了枯草芽孢桿菌BJ3-2為可轉(zhuǎn)化且具有同源重組酶活性的RecE+菌株,可通過同源重組方式對其染色體進行改造,為今后利用同源重組或Red/ET重組[18-20]等方法改良食品級菌株奠定基礎(chǔ)。
基因表達產(chǎn)物活性的增加主要通過在結(jié)構(gòu)基因上游調(diào)控區(qū)增加強啟動子、增加結(jié)構(gòu)基因劑量、優(yōu)化結(jié)構(gòu)基因等方式進行。實驗獲得的重組菌株BJ3-2A2較原菌株BJ3-2從glsA基因數(shù)量來看新加入1個glsA1和1個glsA2,然而重組菌株BJ3-2A2與原菌株BJ3-2的谷氨酰胺酶活力對比可以看出重組菌株的谷氨酰胺酶活力比原菌株高2.36倍。因此在忽略其他影響因素的情況下認為新增加的glsA1基因能提高1倍酶活力,而glsA2基因提高的酶活力為glsA1的1.36倍。由此可知:glsA2基因在重組菌株中能夠表達谷氨酰胺酶活性,但其酶活力較原菌株具體提高多少仍需通過雙交換置換等方式驗證。實驗采用單交換整合的方式將glsA2基因整合入枯草芽孢桿菌BJ3-2染色體上,同時質(zhì)粒上的抗性基因及其他片段也隨之整合入染色體上,因此該菌株不能直接用于食品(豆豉)發(fā)酵。
[1] HONG H A, HUANG J M, KHANEJA R, et al. The safety of Bacillus subtilis and Bacillus indicus as food probiotics[J]. Journal of Applied Microbiology, 2008, 105: 510-520.
[2] 董晨, 曹娟, 張跡, 等. 耐高溫α-淀粉酶基因在枯草芽孢桿菌中的高效表達[J]. 應(yīng)用與環(huán)境生物學(xué)報, 2008, 14(4): 534-538.
[3] ULF B, MARION W, THORSTEN E. Versatile expression and secretion vectors for Bacillus subtilis[J]. Current Microbiology, 2006, 52(2): 143-148.
[4] 鄒立扣, 王紅寧, 潘欣. 枯草芽孢桿菌整合載體研究進展[J]. 生物技術(shù)通訊, 2003, 14(6): 525-527.
[5] 賈東旭, 吳擁軍, 李耀中, 等. 細菌型豆豉發(fā)酵芽孢桿菌的篩選與鑒定[J]. 食品科學(xué), 2009, 30(5): 217-221.
[6] 詹壽年, 吳擁軍, 郭倩倩, 等. 高活性谷氨酰胺酶豆豉芽孢桿菌的篩選及glsA基因的克隆[J]. 貴州農(nóng)業(yè)科學(xué), 2011, 39(6): 119-122.
[7] 朱芳, 鄧思, 羅立新. 分選酶A在pET32a(+)原核表達載體中的表達和鑒定[J]. 生物技術(shù)通報, 2011(6): 219-222.
[8] 聞潔君, 江文正, 郝文麗, 等. HIV-1nef基因的克隆及原核表達研究[J]. 細胞與分子免疫學(xué)雜志, 2010, 26(4): 348-350.
[9] PARK H W, GE Baoxue, BAUER L S, et al. Optimization of Cry3A yields in Bacillus thuringiensis by use of sporulation-dependent promoters in combination with the STAB-SD mRNA sequence[J]. Applied and Environmental Microbiology, 1998, 64(10): 3932-3938.
[10] 張曉慧. 海藻糖改善枯草芽孢桿菌電轉(zhuǎn)化方法的研究[D]. 南京: 南京農(nóng)業(yè)大學(xué), 2010.
[11] YANG Mingming, ZHANG Weiwei, BAI Xiaoting, et al. Electroporation is a feasible method to introduce circularized or linearized DNA into B. subtilis chromosome[J]. Molecular Biology Reports, 2010, 37: 2207-2213.
[12] 劉萍, 夏立秋, 胡勝標, 等. 外源基因在蘇云金桿菌染色體上的定點整合及表達[J]. 微生物學(xué)報, 48(5): 661-666.
[13] 石愛琴. 分子伴侶對枯草芽孢桿菌表達pga的影響以及pga的整合表達[D]. 杭州: 浙江理工大學(xué), 2009.
[14] 鄒立扣, 王紅寧, 潘欣. 枯草芽孢桿菌整合載體研究進展[J]. 生物技術(shù)通訊, 2003, 14(6): 525-527.
[15] 侯化鵬. 枯草芽孢桿菌中性蛋白酶nprE的定向進化研究[D]. 武漢:華中農(nóng)業(yè)大學(xué), 2008.
[16] 夏雨成, 玉梁, 李達倩, 等. 枯草芽孢桿菌分泌載體構(gòu)建及其對脂肪酶A的分泌表達[J]. 安徽農(nóng)業(yè)科學(xué), 2011, 39(30): 18435-18437.
[17] 劉大偉, 王正祥. PspA外源表達對枯草芽孢桿菌168蛋白質(zhì)分泌的影響[J]. 天然產(chǎn)物研究與開發(fā), 2008(20): 855-858.
[18] 唐恒明, 劉先凱, 高美琴, 等. 炭疽芽孢桿菌假想S-層蛋白SLP 缺失突變體的構(gòu)建[J]. 生物技術(shù)通訊, 2009, 20(2): 161-165.
[19] 喻江, 范國權(quán), 李璐, 等. 枯草芽孢桿菌224 YPIQ基因敲除及其對溶血性的影響[J]. 東北農(nóng)業(yè)大學(xué)學(xué)報, 2010, 41(7): 74-78.
[20] 薛可, 李峰, 羅光彬, 等. 利用Red同源重組系統(tǒng)進行牛β-酪蛋白基因敲除[J]. 遺傳, 2007, 29(5): 570-574.
Targeted Integration of Glutaminase Gene glsA2with High Activity into the Bacillus subtilis BJ3-2 Chromosome
LU Biao, WU Yong-jun*, LIU Yan-min, WANG Ya-juan, TANG Xue
(College of Life Science, Guizhou University, Guiyang 550025, China)
With glsA1in BJ3-2 as the homologous sequence, glutaminase gene glsA2with high activity was targeted and integrated into the Bacillus subtilis BJ3-2 chromosome by constructing a single-exchange integrative vector. A recombinant strain BJ3-2A2was obtained with good genetic stability. The results of detection showed that glutaminase activity in the recombinant strain was 3.36 times higher than that in the original strain BJ3-2. The amino acid contents in douchi fermented with BJ3-2A2and the original BJ3-2 were measured, and the results showed that glutamate content in fermentation products from the recombinant strain was increased by 12.8% as compared to that observed with the original strain. All the analyses show that B. subtilis BJ3-2 is transformable as a RecE+strain and that glutaminase gene glsA2can be highly expressed in the BJ3-2 chromosome, thus improving the flavour of douchi.
Bacillus subtilis; homologous recombinant; single-exchange; glutaminase-encoding gene
Q939.97
A
1002-6630(2014)01-0141-04
10.7506/spkx1002-6630-201401027
2012-12-20
國家自然科學(xué)基金項目(31260394);貴陽市科技計劃項目(筑科工合同字[2010]第1-68號);貴州省科技重大專項(黔科合重大專項字[2013]6013號)
盧彪(1987—),男,碩士研究生,研究方向為應(yīng)用微生物。E-mail:liaoyuan612@163.com
*通信作者:吳擁軍(1971—),男,教授,博士,研究方向為食品生物技術(shù)。E-mail:wyjbio@163.com
