菌根真菌侵染對植物生物量累積的影響
2014-02-09 07:58:45唐旭利張倩媚
生態環境學報 2014年9期
關鍵詞:優勢
童 琳,唐旭利*,張 靜,張倩媚
1. 中國科學院華南植物園,廣東 廣州 510650;2. 中國科學院大學,北京 100049;3. 中國科學院退化生態系統植被恢復與管理重點實驗室,廣東 廣州 510650
菌根真菌侵染對植物生物量累積的影響
童 琳1,2,3,唐旭利1,3*,張 靜1,2,3,張倩媚1
1. 中國科學院華南植物園,廣東 廣州 510650;2. 中國科學院大學,北京 100049;3. 中國科學院退化生態系統植被恢復與管理重點實驗室,廣東 廣州 510650
為了從生態系統尺度探討菌根資源與植物生物量累積的關系,調查了鼎湖山不同成熟度的三個森林群落主要優勢樹種的菌根侵染情況。綜合分析各森林群落優勢樹種的個體數、生物量和菌根侵染率發現:1)菌根侵染率與徑向生長速率相關,植物生長迅速的階段菌根侵染率更高。中徑級(胸徑15~30 cm)的馬尾松(Pinus massoniana)和錐(Castanopsis chinensis)的侵染率比小徑級(胸徑1~15 cm)個體的侵染率高,而大徑級個體(胸徑30 cm以上)的侵染率略低于中徑級個體的侵染率。木荷(Schima superba)則表現出侵染率隨著胸徑增大而增高的趨勢。2)樹種在群落內的侵染率越高,其對群落生物量的貢獻率越大。如馬尾松在馬尾松林和混交林的侵染率分別為(77.30±18.02)%和(40.50±14.42)%,其對馬尾松林群落生物量的貢獻率達到87.43%,是對混交林生物量貢獻率(17.51%)的5倍。混交林和闊葉林的共有優勢樹種錐的侵染率和生物量貢獻率也有存在相同規律。3)根系碳儲量占群落總碳儲量比例較高的群落其優勢樹種平均侵染率相對較高。馬尾松林、混交林和季風常綠闊葉林中,根系碳儲量占群落總碳儲量的比例分別為55%、54%、42%,群落優勢樹種平均侵染率分別為(66.73±10.55)%、(46.97±27.28)%、(54.22±25.45)%,馬尾松林的根系碳儲量和平均侵染率均高于混交林和季風常綠闊葉林。以上結果表明,菌根真菌侵染對于植物個體生長速率以及群落水平的生物量累積具有一定的促進作用。
菌根;群落;侵染率;生物量;胸徑;根系碳儲量
叢枝菌根(arbuscular mycorrhiza,AM)和外生菌根(ectomycorrhiza,ECM)是森林生態系統中分布最為廣泛的菌根類型,約80%的維管植物能形成這兩種類型的菌根(Smith和Read,2008;Brundrett,2009)。在菌根共生體中,宿主植物將一部分光合作用固定的碳——己糖提供給真菌(Paul和Kucey,1981;Jakobsen和Rosendahl,1990;Nemec和Vu,1990;Shachar-Hill等,1995;Fitter,2006;Helgason和Fitter,2009;Smith和Smith,2012),菌根真菌則通過根外菌絲增大根系對土壤溶液的吸收面積,且能活化K+、Ca2+、Mg2+等土壤陽離子(Arocena和Glowa,2000;Jentschke等,2000),提高植物對氮、磷、鉀、鈣、鎂等營養元素的吸收效率(Nemec和Vu,1990;Hodge,2001,2003;Fitter,2006;Helgason和Fitter,2009;Smith和Smith,2012)。菌根共生體對于植物群落的穩定性有重要意義。近年來,我國已經開展過一些關于森林生態系統菌根資源的研究。自然生境的菌根研究集中在ECM真菌和AM真菌的種屬分類及生態分布等方面(石兆勇等,2003;Zhang和Guo,2005;房輝等,2006;賀學禮等,2010;何新華等,2012),對于生態系統尺度菌根的生態學功能研究較為缺乏。已有的菌根生態學方面的研究多以盆栽栽培實驗為主要手段,研究對象多為草本植物(Martin和Stutz,2004;蔡曉布等,2005;于永光和趙斌,2008),研究內容側重菌根真菌對宿主幼苗養分吸收和個體發育的影響(閻秀峰和王琴,2004;于永光和趙斌,2008)。目前,關于菌根對植物個體生長速率和群落水平的生物量累積的影響報道不多。
侵染率(侵染率=被侵染的根段數/觀察的總根段數)是較為客觀的菌根研究指標,能夠反映某一時段生態系統的菌根侵染狀況。土壤理化性質(如氮、磷元素形態)、降水、溫度以及植物種類都直接影響真菌孢子的萌發、菌絲長度和菌絲分支,通過測定菌根真菌的侵染率,可以觀測菌根真菌與環境條件的影響與適應特征。在群落生態學中,往往以相對多度、相對頻度和相對顯著度之和反映植物物種在群落中的相對重要值(Curtis和Mcintosh,1951),植物數量特征是群落生態學較為核心的研究指標。菌根真菌與植物根系共生在群落生態學中的重要作用較少被關注。
為了探討菌根真菌侵染對群落中植物數量和生物量累積的影響,本研究以鼎湖山自然保護區的三個森林群落為研究對象,調查各群落中優勢樹種的菌根侵染率、優勢樹種數量和生物量特征,綜合分析侵染率對樹種徑向生長速率、生物量和根系碳儲量的可能影響。對于研究森林生態系統菌根資源和生物量累積受植物、真菌調控的影響具有重要意義。
1 研究地概況
研究地點位于廣東省肇慶市的鼎湖山自然保護區(112°30′39″~112°33′41″E,23°09′21″~23°11′30″N),該區氣候屬于典型的南亞熱帶濕潤季風氣候,年均氣溫20.9 ℃,年均降雨量1929 mm,雨季(4—9月)降雨量高達年降雨量的80%。土壤為酸性赤紅壤。保護區內具有三種典型的亞熱帶植被類型:馬尾松林(PF)、針闊葉混交林(MF)和季風常綠闊葉林(BF),分別代表該區自然演替初期、中期和頂級階段。
演替初期的PF分布在海拔50~200 m,由20世紀50年代人工種植的馬尾松純林發育而成。優勢樹種為馬尾松(Pinus massoniana)、白楸(Mallotus paniculatus)和三椏苦(Evodia lepta)。演替中期的MF分布在海拔100~250 m,是闊葉樹種入侵馬尾松人工林后演替形成的群落類型,主要木本植物為馬尾松、錐(Castanopsis chinensis)和木荷(Schima superba)。演替頂級的BF分布于海拔250~400 m,已有400年的保護歷史,主要木本植物類群有錐、肖蒲桃(Acmena acuminatissima)、木荷、云南銀柴(Aporusa yunnanensis)、橄欖(Olea europaea)、窄葉半楓荷(Pterospermum lanceaefolium)、厚殼桂(Cryptocarya chinensis)、白顏樹(Gironniera subaequalis)以及臀果木(Pygeum topengii)等。其中,馬尾松為PF與MF的共有優勢樹種,錐和木荷為MF與BF的共有優勢樹種。
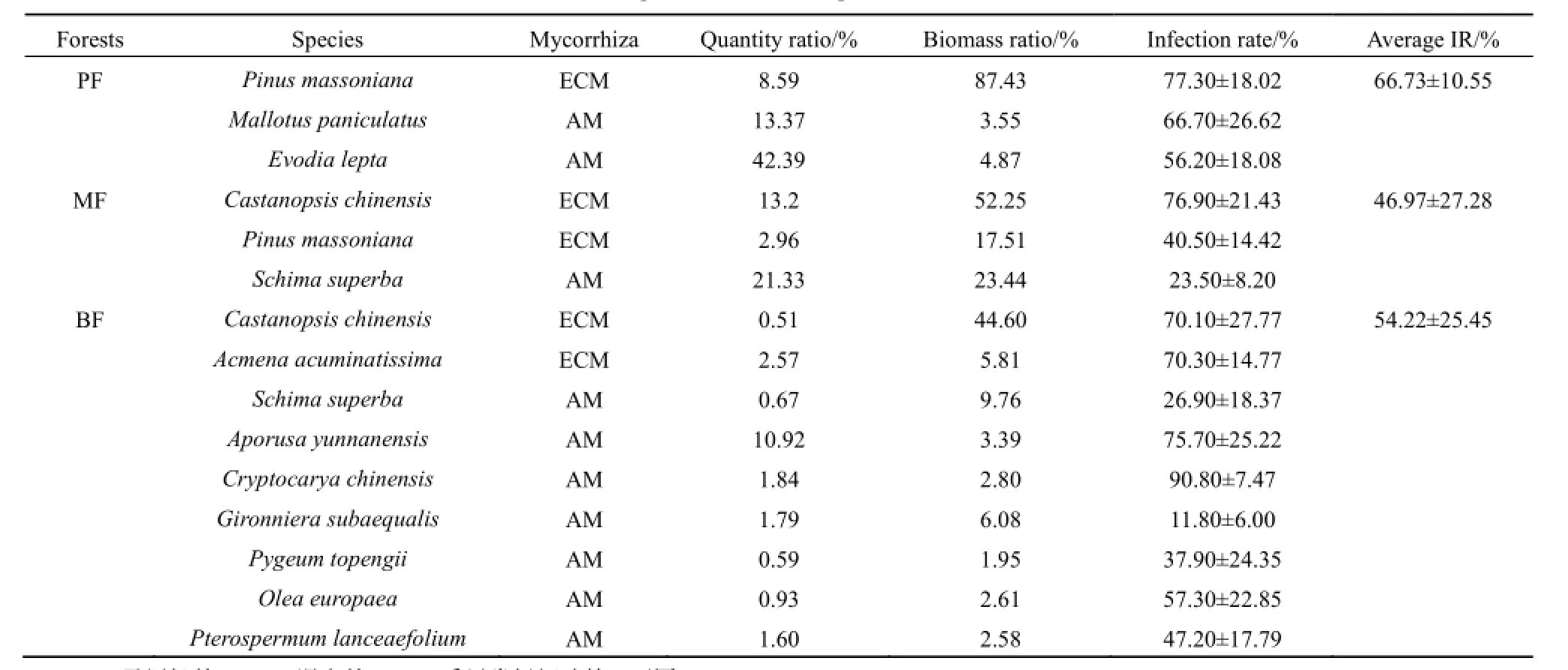
表1 三個森林群落的優勢樹種基本狀況Table 1 Properties of dominant species in three forests
2 研究方法
2.1 樣品采集
選取三個森林群落內生物量比重>2%和個體數比重>0.5%的優勢樹種為研究對象(表1),于2012年7月對各群落主要優勢樹種的代表性個體進行采樣。三個森林群落之間的共有優勢樹種馬尾松、錐和木荷的生物量隨著胸徑的分布特征為15~30 cm胸徑的個體生物量占該樹種總生物量的比重最大,胸徑1~15 cm和胸徑30 cm以上的個體生物量相對較低(圖1)。采樣時,選取1~15 cm(小徑級),15~30 cm(中徑級),>30 cm(大徑級)三個胸徑范圍內的代表性個體(若樹種缺失某一徑級范圍內個體,則該徑級范圍不作分析)。每個徑級選取5株個體進行采樣,個體間相距100 m以上,在樣木樹冠滴水線東、西、南、北各向采集根樣(仝瑞建,2011)。采集根系樣品時,先根據粗根辨識需采集的樹種,再順著側根方向挖取直徑2 mm以下的細根,同時用圖文方式記錄根系特征供鑒定。細根樣品帶回實驗室,挑洗供測定侵染率使用。
優勢樹種的個體數和生物量數據來自鼎湖山站基礎數據庫。
2.2 室內分析
2.2.1 外生菌根(ECM)侵染率測定
細根用FAA(福爾馬林-醋酸-酒精固定液,formalin–acetic acid–alcohol)固定24 h后取出,洗凈,用解剖刀將末級根切下,每份樣品取100個根段,用Stemi SV11體視顯微鏡進行鏡檢,根尖有菌套或根外皮層細胞間隙有哈蒂氏網者為外生菌根。外生菌根侵染率由ECM侵染根段數占鏡檢根段總數的百分比表示:
ECM侵染率=(ECM侵染根段數/鏡檢總根段數)×100%
2.2.2 內生菌根(AM)侵染率測定
采用Phillips和Hayman(Phillips和Hayman,1970)方法對細根染色,用解剖刀將末級根切下,每份樣品取100個根段,壓片,用AXIOPLAN 2 Iminage正立熒光顯微鏡進行鏡檢,根尖有泡囊、胞內或胞間有叢枝或菌絲者為叢枝菌根。叢枝菌根的侵染率由AM侵染根段數占鏡檢根段總數的百分比表示:
AM侵染率=(AM侵染根段數/鏡檢總根段數)×100%
2.3 數據處理
應用PASW Statistics 18.0進行統計分析和SigmaPlot 10.0作圖。
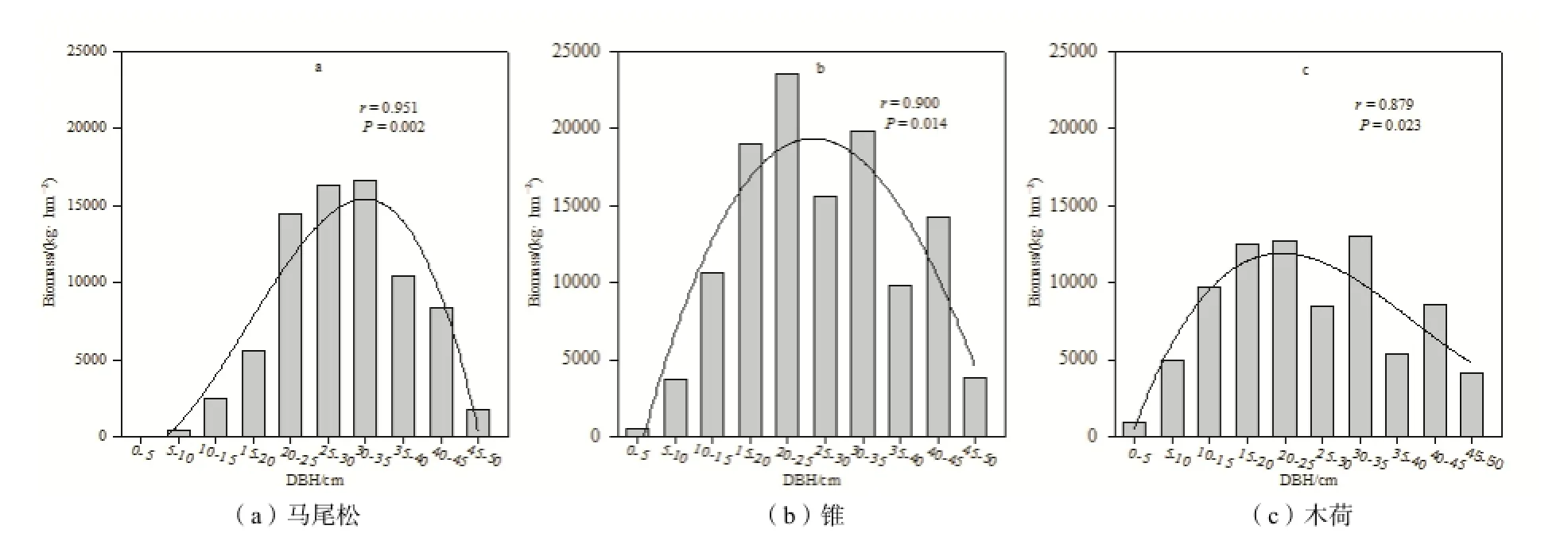
圖1 群落之間的共有優勢樹種生物量隨胸徑的分布規律Fig. 1 Biomass of codominant species with different DBHs
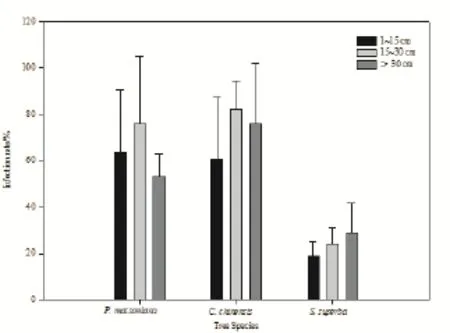
圖2 不同胸徑樹種的菌根侵染率Fig. 2 Infection rate of codominant species with different DBHs
3 結果
3.1 生物量隨胸徑的分布規律
最近(2010年)的樣地調查結果顯示,三個森林群落之間的共有優勢樹種馬尾松、錐和木荷的生物量隨著胸徑變化呈現一定規律性,15~30 cm胸徑的個體生物量占該樹種總生物量的比重較大,其中25~30 cm胸徑的個體生物量達到峰值;胸徑1~15 cm和胸徑30 cm以上的個體生物量占該樹種總生物量的比重明顯低于15~30 cm胸徑的個體(圖1)。
3.2 不同徑級優勢樹種菌根侵染率
按照群落之間的共有優勢種馬尾松、錐和木荷的生物量隨胸徑的變化規律將采樣個體按照1~15、15~30和>30 cm劃分為小、中、大3個胸徑等級,比較侵染率隨徑級大小的變化趨勢。如圖2所示,馬尾松和錐中徑級(15~30 cm)的個體平均侵染率較小徑級(1~15 cm)和大徑級(>30 cm)的高,整體呈現侵染率隨個體胸徑的增大先升高后降低的趨勢。木荷的侵染率則表現出隨著胸徑的增大而增高的趨勢(圖2),但各個樹種侵染率在三個胸徑范圍之間的差異并未達到統計學上的顯著水平(P>0.05)。
3.3 優勢樹種菌根侵染率與樹種生物量
比較群落之間的共有優勢樹種發現,侵染率越高的共有優勢樹種對群落生物量的貢獻比重往往越高。PF中馬尾松生物量占總生物量的87.43%,馬尾松在該群落的侵染率高達(77.30±18.02)%。MF中馬尾松生物量僅占群落生物量的17.51%,相應的,馬尾松在MF的侵染率也較低,為(40.50±14.42)%。錐對MF生物量的貢獻高達52.25%,在MF中錐的侵染率相應較高,達到(76.90±21.43)%。錐對BF生物量的貢獻率(44.60%)略低于對MF生物量的貢獻率,在BF內錐的菌根侵染率(70.10±27.77)%也低于其在MF內的侵染率。MF和BF的共有優勢樹種木荷的侵染率與生物量百分比之間沒有明顯的規律性(表1、圖3)。對群落之間共有優勢樹種的侵染率與生物量百分比作Pearson相關性分析發現,二者呈顯著的正相關關系(P<0.05),樹種侵染率高,則其生物量百分比也相對較高(表2)。

圖3 三個森林群落之間的共有優勢種的菌根侵染率Fig.3 Mycorrhizal infection rate of codominant species in three forests
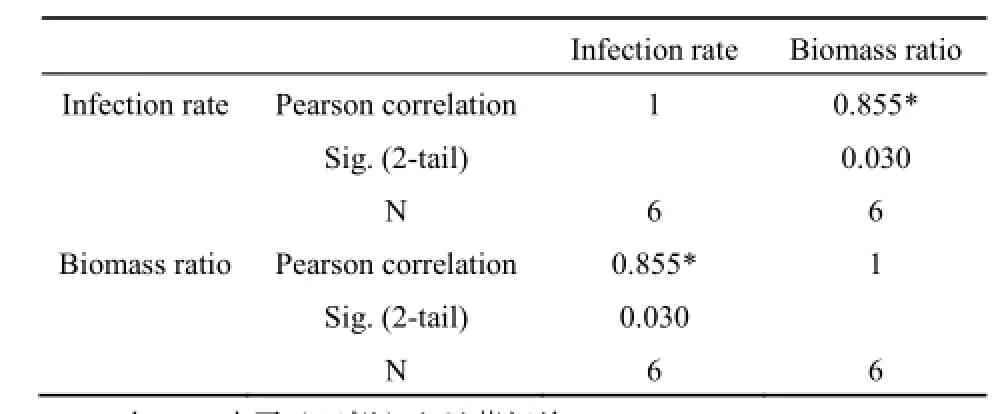
表2 群落間共有優勢種菌根侵染率與生物量百分比的相關性Table 2 Correlation between infection rate and biomass ratio
3.4 三個森林群落的菌根侵染特征
馬尾松林的馬尾松(ECM)生物量(87.43%)比重占絕對優勢地位,混交林中,錐(ECM)、馬尾松(ECM)和木荷(AM)的生物量比重分別為52.25%、17.51%和23.44%,季風常綠闊葉林的錐(ECM)和木荷(AM)對群落生物量貢獻的較大,分別為44.60%、9.76%。馬尾松林、混交林和季風常綠闊葉林的平均侵染率分別為(66.73±10.55)%、(46.97±27.28)%、(54.22±25.45)%(表2)。馬尾松林的平均侵染率相對高于混交林和季風常綠闊葉林。
4 討論
4.1 真菌侵染率與植物個體生長速率
本研究表明,胸徑從1~15 cm過渡到15~30 cm,馬尾松和錐(ECM植物)的侵染率增高,胸徑達到30 cm以上時,侵染率呈降低趨勢。對油松幼苗的研究發現外生菌根真菌侵染率與油松幼苗生長成正相關(r=0.8924,P<0.05),植株生長越迅速,其侵染率越高(張茹琴等,2011)。已有研究表明南亞熱帶地區小于12齡的馬尾松年均徑向增長率(1.31 cm·a-1)高于12齡以上的年均徑向增長率(1.03 cm·a-1)。30 cm胸徑的馬尾松樹齡約為28 a(盧翠香等,2012),幼齡期的馬尾松菌根侵染率較高,快速生長一定時間段(28 a左右,胸徑達到30 cm以上)后,生長速率降低,生物量累積緩慢。因此,菌根侵染率與徑向生長有關,植物生長迅速的時間段菌根侵染率也更高,從而提高植物根系的養分吸收效率,促進植物生長。對馬尾松采伐年齡的模型預測值為30~45 a(李賢偉等,1996;丁貴杰等,1998;韋新良等,2001),與本研究推算的馬尾松徑向生長速率劇減階段的樹齡接近。木荷的侵染率隨著胸徑的增大逐漸增高,表明木荷的徑向增長速率可能大于馬尾松和錐,或徑向生長劇減期晚于馬尾松和錐,使得木荷的侵染率呈現出在所測胸徑范圍內隨著胸徑的增大而增高的趨勢。然而,由于計算過程中將三個群落作為一個整體,而各群落演替程度不同,同一徑級樹種個體的年齡差異使分析結果的標準差較大,從而導致不同徑級之間的侵染率差異不顯著。
4.2 菌根侵染對群落養分積累的影響
對三個不同演替階段森林群落侵染率的分析發現群落之間共有優勢樹種菌根侵染率和樹種生物量隨著森林群落的演替呈下降的趨勢,侵染率高的共有優勢樹種對群落生物量的貢獻率往往也較大。馬尾松是演替初期的PF的建群樹種,其生物量百分比和侵染率在演替初期的PF中皆高于演替中期的MF;錐是演替中期的MF的優勢樹種,其生物量百分比和侵染率在演替中期的MF中皆高于演替頂級的BF(表2、圖3)。不同演替階段森林群落的優勢樹種類型、個體數存在較大差異。PF樹種類型比較單一,優勢樹種個體數較多,BF樹種類型豐富,優勢樹種個體數較少,MF優勢樹種類型和數量在三個群落內屬于中間位置。長期監測的結果表明,三個森林的根系碳儲量分別為群落碳儲量的55%(PF)、54%(MF)、42%(BF)(周國逸等,2005),優勢樹種平均侵染率分別為(66.73±10.55)%、(46.97±27.28)%、(54.22±25.45)%(表2)。馬尾松林的平均侵染率相對高于混交林和季風常綠闊葉林。菌根真菌侵染對植物和群落的養分凈積累有一定的積極影響,群落之間共有優勢樹種的侵染率較高的樹種,其對群落養分凈積累的貢獻比重更大。在陸地生態系統中,菌根真菌通過菌絲體、土壤微生物、植物個體和群落影響生態系統的過程與功能(Rillig,2004)。因此,菌根真菌在植物生物量累積過程中意義重大。
5 結論
本研究表明,菌根真菌侵染對于植物個體生長速率以及群落水平的生物量累積具有一定的促進作用。同一樹種的菌根侵染率隨著胸徑增大有增高的趨勢,菌根侵染對植物生長的促進作用也表現在群落之間的共有優勢樹種侵染率和生物量的聯系上,侵染率越高的共有優勢樹種對群落生物量的貢獻率往往越大,在群落競爭中占優勢地位。從群落水平看,根系碳儲量高的森林群落中,群落優勢樹種平均侵染率相對較高。然而,真菌與植物相互調節和適應的具體機制還有待進一步探索。在真菌-植物的調節與適應機制的研究中,分子生物學原理與方法有必要作為群落生態學研究的輔助手段,如利用穩定同位素標記研究菌根與植物營養元素的轉運,運用質譜成像技術研究菌根真菌的活動及其產物,以及從基因層面探索調控菌根真菌和植物發育的基因和信號物質等在未來群落生態學研究中至關重要。
致謝:感謝莫定升、向傳銀、鄧民秀在野外采樣和分析測試工作中給予的幫助;感謝李坤和徐偉強博士在論文修改方面的寶貴建議。
AROCENA J M, GLOWA K R. 2000. Mineral weathering in ectomycorrhizosphere of subalpine fir (Abies lasiocarpa (Hook.) Nutt.) as revealed by soil solution composition[J]. Forest Ecology and Management, 133(1/2): 61-70.
BRUNDRETT M C. 2009. Mycorrhizal associations and other means of nutrition of vascular plants: understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis[J]. Plant Soil, 320(1/2): 37-77.
CURTIS J T, MCINTOSH R P. 1951. An upland forest continuum in the prairie-forest border region of Wisconsin[J]. Ecology, 32(3): 476-496.
FITTER A H. 2006. What is the link between carbon and phosphorus fluxes in arbuscular mycorrhizas? A null hypothesis for symbiotic function[J]. New Phytologist, 172(1): 3-6.
HE X H, DUAN Y H, CHEN Y L, et al. 2012. A 60-year journey of mycorrhizal research in China: Past, present and future directions[J]. Science China Ser. C Life Sciences, 53(12): 1374-1398.
HELGASON T, FITTER A H. 2009. Natural selection and the evolutionary ecology of the arbuscular mycorrhizal fungi (Phylum Glomeromycota) [J]. Experimental Botany, 60(9): 2465-2480.
HODGE A. 2001. Arbuscular mycorrhizal fungi infuence decomposition of, but not plant nutrient capture from, glycine patches in soil[J]. New Phytologist, 151(3): 725-734.
HODGE A. 2003. Plant nitrogen capture from organic matter as affected by spatial dispersion, interspecific competition and mycorrhizal colonization[J]. New Phytologist, 157(2): 303-314.
JAKOBSEN I, ROSENDAHL L. 1990. Carbon flow into soil and external hyphae from roots of mycorrhizal cucumber plants[J]. New Phytologist, 115(1): 77-83.
JENTSCHKE G, BANDES B, KUHN A J, et al. 2000. The mycorrhizal fungus Paxillus involutus transports magnesium to Norway spruce seedlings[J]. Evidence from stable isotope labeling. Plant and Soil, 220(1/2): 243-246.
MARTIN C A, STUTZ J C. 2004. Interactive effects of temperature and arbuscular mycorrhizal fungi on growth, P uptake and root respiration of Capsicum annuum L[J]. Mycorrhiza, 14(4): 241-244.
NEMEC S, VU J C V. 1990. Effects of soil phosphorus and Glomus intraradices on growth, nonstructural carbohydrates, and photosynthetic activity of Citrus aurantium[J]. Plant Soil, 128(2): 257-263.
PAUL E A, KUCEY RMN. 1981. Carbon flow in plant microbial associations[J]. Science, 213(4506): 473-474.
PHILLIPS J M, HAYMAN D S. 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection[J]. Transactions of the British Mycological Society, 55(1): 157-160.
RILLIG M C. 2004. Arbuscular mycorrhizae and terrestrial ecosystem processes[J]. Ecology letters, 7(8): 740-754.
SHACHAR-HILL Y, PFEFFER PE, DOUDS D, et al., 1995. Partitioning of intermediary carbon metabolism in vesicular-arbuscular mycorrhizal leek[J]. Plant Physiology, 108(1): 7-15.
SMITH S E, READ D J. 2008. Mycorrhizal symbiosis[M]. 3rd edn. Academic Press, London.
SMITH S E, SMITH F A. 2012. Fresh perspectives on the roles of arbuscular mycorrhizal fungi in plant nutrition and growth[J]. Mycologia, 104(1): 1-13.
ZHANG Y, GUO L D. 2005. Two new records of arbuscular mycorrhizal fungi in China[J]. Mycosystema, 24(3): 465-467.
蔡曉布, 錢成, 彭岳林, 等, 2005. 環境因子對西藏高原草地植物叢枝菌根真菌的影響[J]. 應用生態學報, 16(5): 859-864.
丁貴杰. 1998. 貴州馬尾松人工建筑材林合理采伐年齡研究[J]. 林業科學, 34(3): 40-46.
房輝, Damodaran PN, 曹敏. 2006. 西雙版納熱帶次生林中的從枝菌根調查[J]. 生態學報, 26(12): 4179-4185.
賀學禮, 劉雪偉, 李英鵬. 2010. 沙坡頭地區沙冬青AM 真菌的時空分布[J]. 生態學報, 30(2): 370-376.
李賢偉, 胡庭興, 楊禎祿. 1996. 馬尾松天然林采伐年齡的研究[J]. 四川農業大學學報, 14(13): 437-439.
劉永俊, 石國璽, 毛琳, 等, 2011. 施肥對垂穗披堿草根系中叢枝菌根真菌的影響[J]. 應用生態學報, 22(12): 3131-3137.
劉永俊, 鄭紅, 何雷, 等, 2009. 檸條根系中叢枝菌根真菌的季節性變化及影響因素[J]. 應用生態學報, 20(5): 1085-1091.
盧翠香, 徐峰, 覃引鸞, 等, 2012. 人工林馬尾松晚材率、年輪寬度和組織比量變異研究[J]. 廣西林業科學, 41(2): 81-85.
石兆勇, 陳應龍, 劉潤進. 2003. 西雙版納地區龍腦香科植物根圍的AM真菌. 菌物學報, 22(3): 402-409.
孫玥, 全先奎, 賈淑霞, 等, 2007. 施用氮肥對落葉松人工林一級根外生菌根侵染及形態的影響[J]. 應用生態學報, 18(8): 1727-1732.
仝瑞建, 劉雪琴, 王發園, 等, 2011. 沙田柚根圍AM真菌的生境適宜性和季節變化性[J]. 應用與環境生物學報, 17(5): 684-687.
韋新良, 郭仁鑒, 趙斌. 2001. 浙江省馬尾松天然林生長模型及采伐年齡的確定[J]. 浙江林學院學報, 18(4): 333-336.
閻秀峰, 王琴. 2004. 兩種外生菌根真菌在遼東櫟幼苗上的混合接種效應[J]. 植物生態學報, 28(1): 17-23.
于永光, 趙斌. 2008. 不同pH水平下兩種菌根真菌對紫云英生長的影響及其相互作用[J]. 菌物學報, 27(2): 209-216.
張茹琴, 唐明, 張海涵. 2011. 四種外生菌根真菌對油松幼苗的抗猝倒病和促生作用[J]. 菌物學報, 30(5): 812-816.
周國逸, 周存宇, LIU Shuguang, 等, 2005. 季風常綠闊葉林恢復演替系列地下部分碳平衡及累積速率[J]. 中國科學D: 地球科學, 35(6): 502-510.
Influence of mycorrhizal colonization on plant biomass accumulation.
TONG Lin1,2,3, TANG Xuli1,3*, ZHANG Jing1,2,3, ZHANG Qianmei1
1. South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystem, Chinese Academy of Sciences, Guangzhou 510650, China
To reveal the relations between mycorrhizal resources and biomass accumulation at system scale, we analysed possible impact of mycorrhizal colonization on growth rate and biomass accumulation of dominant tree species in three forests at Dinghushan Nature Reserve. Mycorrhizal infection rate, quantity percentages and biomass of doiminant tree species in a given ecosystem were investigated in this study. Our research shows that: 1) Infection rate changed accordingly with growth rate. Trees that grow more rapidly might have higher infection rate. 2) Plant with higher infection rate turned to have lager biomass ratio in its community. e.g., infection rate of Pinus massoniana were (77.30±18.02)% and (40.50±14.42)% in PF and MF, respectively, its biomass ratio was 87.43% in PF, which was five times of that (17.51%) in MF. This tendency was the same for Castanopsis chinensis, a codominant species in MF and BF. 3) Infection rate and root carbon content were relatively higher in PF (pine forest, early successional stage) than that in MF (mixed coniferous and broadleaved forest, middle successional stage) and BF (monsoon evergreen broadleaved forest, late successional stage). In PF, MF and BF, root carbon storage ratio was 55%, 54% and 42%, mycorrhizal infection rate was (66.73±10.55)%, (46.97±27.28)% and (54.22±25.45)%, respectively. This study thus suggests that mycorrhizal colonization may promote plant growth and biomass accumulation of forest community.
mycorrhiza; community; infection rate; biomass; diameter at breast height; root carbon storage ratio
Q948
A
1674-5906(2014)09-1520-06
童琳,唐旭利,張靜,張倩媚. 菌根真菌侵染對植物生物量累積的影響[J]. 生態環境學報, 2014, 23(9): 1520-1525.
TONG Lin, TANG Xuli, ZHANG Jing, ZHANG Qianmei. Influence of Mycorrhizal Colonization on Plant Biomass Accumulation [J]. Ecology and Environmental Sciences, 2014, 23(9): 1520-1525.
國家自然科學基金項目(31270499);中國科學院青年創新促進會資助
童琳(1990年生),女,碩士研究生,主要從事生態系統生態學和菌根共生學研究。E-mail: tonglin@scbg.ac.cn通信作者:E-mail: xltang@scib.ac.cn
2014-07-22
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
