番茄抗TYLCV基因新標記的開發及其在多抗聚合選擇中的應用
2014-02-23 10:52:58李菲菲楊文才
中國蔬菜 2014年10期
胡 霞 王 蓉 李菲菲 周 濤 楊文才
(中國農業大學農學與生物技術學院,北京 100193)
自20世紀90年代我國發現番茄黃化曲葉病毒病(TYLCV)以來,該病害已遍布我國番茄主產區,嚴重威脅我國的番茄生產(葉青靜 等,2009;李小靖和葉志彪,2010;李常保 等,2012;吳篆芳 等,2013)。盡管利用殺蟲劑等方法能在一定程度上控制該病毒傳播者——煙粉虱的數量,但是經常施用殺蟲劑會使煙粉虱產生抗藥性(Horowitz et al.,2007),也會造成嚴重的環境污染(Palumbo et al.,2001)。采用細孔網或者紫外吸收塑料膜遮蓋等物理方法(Cohen & Antignus,1994)起到了一定的防治效果,然而這些物理防治方法不能保證正常的光照,還會形成高溫高濕的環境,影響番茄植株的正常生長。利用植物自身的抗性基因是防治TYLCV 最有效的方法(Morales,2001;Lapidot &Friedmann,2002)。
番茄在生長發育過程中除受到多種病毒侵染外,還會同時受到諸如斑點病、瘡痂病和潰瘍病等細菌性病害的危害,培育抗多種病害的品種一直是番茄育種的目標(杜永臣 等,1999)。采用常規育種方法難以在田間進行抗單一病害材料的篩選,分子標記輔助選擇為解決這一難題提供了可能。Sun等(2011)根據番茄斑點病抗性基因Pto位點在抗感病材料中的序列差異建立的InDel 標記和Pei 等(2012)利用番茄瘡痂病抗性基因Rx4位點在抗感材料中的序列差異建立的InDel 標記都是功能基因分子標記,能夠對抗病個體進行準確的選擇。而對于尚未克隆的抗性基因,則可選擇與基因最緊密連鎖的標記,以提高選擇的準確性,如Yang 和Francis(2005)利用與番茄瘡痂病抗性位點Rx3緊密連鎖的標記成功地將Rx3和Pto兩個基因聚合到同一材料中,育成抗番茄斑點病和瘡痂病的材料。從1994年至今,人們已經開發了與5個抗TYLCV基因Ty-1~Ty-5緊密連鎖的可用于抗性基因選擇的分子標記(楊曉慧 等,2012),但這些標記在實際應用中都存在一些問題,如檢測太復雜、穩定性和可靠性不高等。番茄基因組的出現以及抗TYLCV基因Ty-1/Ty-3的克隆,為開發穩定可靠且檢測簡單易行的標記提供了可能。因此,本試驗的目的是開發可用于準確選擇Ty-1/Ty-3和Ty-4的分子標記,并將抗病毒病基因與抗細菌病害基因聚合,培育抗多個病害的育種材料。
1 材料與方法
1.1 試驗材料
本試驗用于標記輔助選擇的番茄(SolanumlycopersicumL.)親本有10個,OH 9242 和里格爾87-5 分別是美國和我國廣泛種植的兩個加工番茄品種,但缺少抗TYLCV 的基因;IBL 2361 和ZN 17是兩個育種中間材料;另有6 份材料分別作為不同病害抗性基因的供體(表1)。將這些材料配制一些雜交組合,加代得到F2群體,供標記輔助選擇用。
2013年6月28日將F2種子播種于288 孔育苗盤中,育苗基質為2V草炭∶1V蛭石。從幼苗上取幼嫩葉片提取DNA 供標記輔助選擇使用。2013年8月9日,將選出的與抗病親本帶型相同的F2單株定植到中國農業大學上莊實驗站日光溫室,收獲F2:3種子。將F2:3種子用4%次氯酸鈉消毒5 min,并用去離子水漂洗3~4次,干燥后保存備用。2014年1月23日播種F2:3種子。待幼苗長出2~3片真葉時,取幼嫩葉片提取DNA,供標記選擇和驗證使用。挑選抗性基因位點純合的單株于2014年3月5日移栽到中國農業大學科學園溫室,進行抗性鑒定。
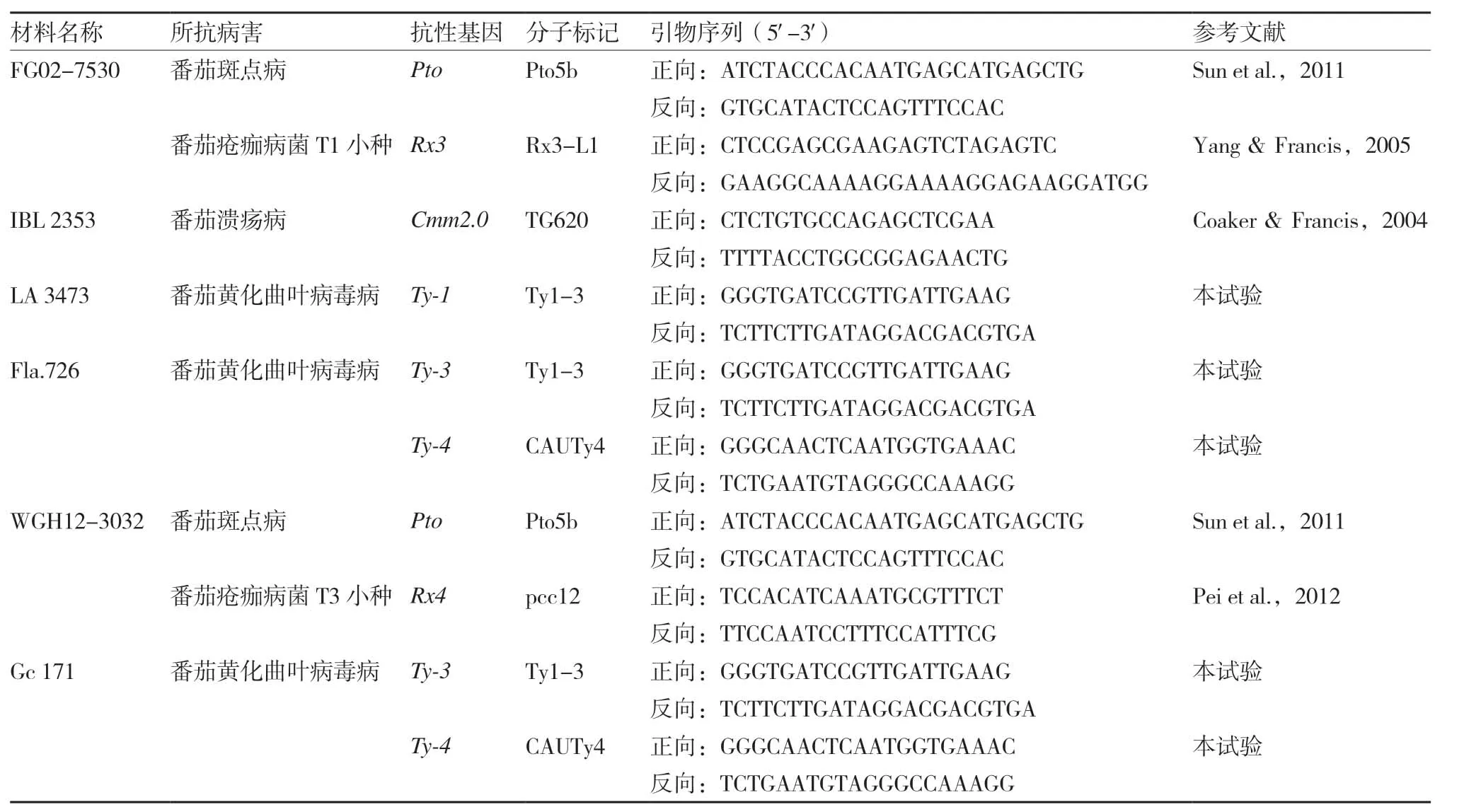
表1 所用抗病親本及標記信息
1.2 抗TYLCV 新標記設計
為了獲得穩定可靠的分子標記,本試驗根據已經克隆的Ty-1和Ty-3基因(Verlann et al.,2013)位點在抗感品種中的序列差異設計引物,獲得了功能基因分子標記Ty1-3。該標記是一個InDel 標記,從抗病材料中擴增到的片段大小為209 bp,從感病材料中擴增到的片段大小為197 bp。同時根據Ji 等(2009)報道的Ty-4在染色體上的位置,選擇與其最緊密連鎖的標記C2_At4g173000,從抗感材料中進行PCR 擴增和序列分析,發現在抗感材料中序列長度不同,據此設計了InDel 標記,命名為CAUTy4。該標記從抗病材料中擴增到的片段大小為219 bp,從感病材料中擴增到的片段大小為214 bp。原來的C2_At4g173000 是一個CAPs 標記(Ji et al.,2009),需要經過PCR、酶切、電泳等過程進行基因型檢測,新設計的兩個InDel 標記都只需要PCR 和凝膠電泳即可完成基因型檢測。
1.3 標記輔助選擇策略
采用堿裂解法(Yang & Francis,2005)提取番茄植株基因組DNA。鑒于有些雜交組合涉及到選擇3個或3個以上抗性基因,為了減少工作量,本試驗首先用一個功能基因標記對相應F2群體的所有植株進行篩選,只有攜帶純合抗性基因位點的植株才被用于篩選第2個基因,依此類推。由于有些群體單株數不夠多,沒有能夠獲得所有基因位點都純合的個體,因此保留了部分基因位點雜合的單株供F2:3繼續選擇。
番茄瘡痂病T1 小種抗性位點Rx3的標記檢測參考Yang 和Francis(2005)的方法,番茄瘡痂病T3 小種抗性位點Rx4的功能基因標記檢測參考Pei等(2012)的方法,番茄斑點病抗性基因Pto的功能基因標記檢測參考Sun 等(2011)的方法,番茄潰瘍病抗性位點Cmm2.0的標記檢測參考Coaker 和Francis(2004)的方法,抗TYLCV基因Ty-1/Ty-3和Ty-4的檢測采用如下方法。
PCR反應體系為10 μL,包括2 μL(約10 ng)DNA模板、3.4 μLTaq10 2×Master Mix(北京奧賽博科技發展有限公司生產)、0.3 μL 10 μmol·L-1正向引物、0.3 μL 10 μmol·L-1反 向引物和4 μL ddH2O。PCR 反應條件為:94℃變性5 min,38個循環的94℃變性45 s、55℃退火45 s和72℃延伸45 s,最后一步反應在72℃下保持5 min。PCR 產物經聚丙烯酰胺凝膠電泳和硝酸銀染色顯帶后,記錄每個單株的基因型。
1.4 抗TYLCV 鑒定
從F2中選出的單株定植到中國農業大學上莊實驗站日光溫室后自然發病,觀察并記載發病情況。F2∶3家系采用人工注射接種在日光溫室內進行抗性鑒定,病毒所屬株系來自上海的TYLCVSH2,接種后28 d 和42 d 各調查一次。病害分級參考Friedmann 等(1998)的方法進行:0級—接種和沒接種的植株一樣,沒有感病癥狀,能夠正常生長;1級—頂端葉片(葉邊緣)出現輕微泛黃;2級—葉片末端有些泛黃和輕微卷曲;3級—大面積葉片變黃,影響更為嚴重,新葉面積減少,但植株可以繼續生長;4級—植物生長發育受到嚴重影響,葉片基本變黃,甚至變紫,杯狀卷曲,停止生長。將0~2級劃分為抗病,3~4級劃分為感病(Ji et al.,2009)。
由于少數入選的F2:3家系群體較小,因此將來自于同一個F2群體的不同F2:3家系算作一個群體,按100×∑(各級病株數×各級代表值)/(調查總株數×最高級代表值)進行病情指數計算。
2 結果與分析
2.1 抗TYLCV基因新標記的可靠性分析
為了檢驗新設計的用于選擇抗TYLCV基因標記的可靠性,本試驗用Ty1-3 和CAUTy4 這兩個標記分析了10 份攜帶Ty-1、Ty-3、Ty-4中1個或多個基因的番茄材料和1 份新的抗病材料LA 1589 的基因型,以感病品種OH 88119 作為對照。結果顯示,兩對引物擴增得到的基因型與材料所攜帶的已知抗性基因完全一致(表2、圖1),表明用這兩個標記來檢測各自的基因型是準確可靠的。同時也發現,新的抗性材料LA 1589 不含有Ty-1、Ty-3和Ty-4基因中的任何一個。
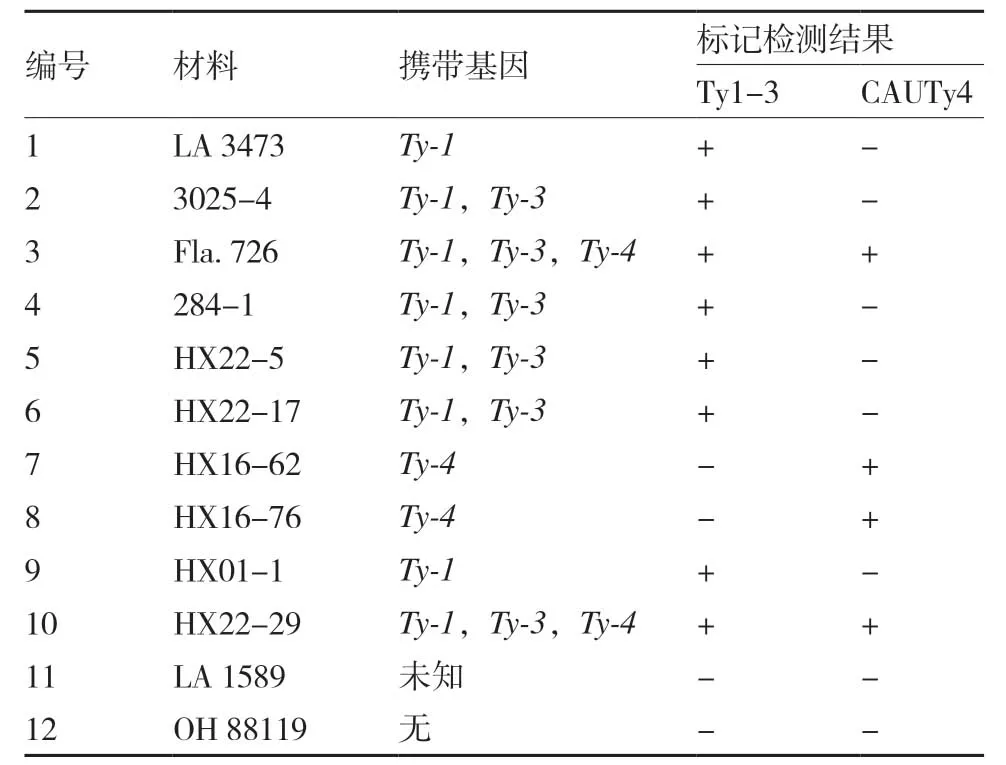
表2 12 份番茄材料經標記Ty1-3 和CAUTy4 檢測的基因型及其攜帶的抗性基因
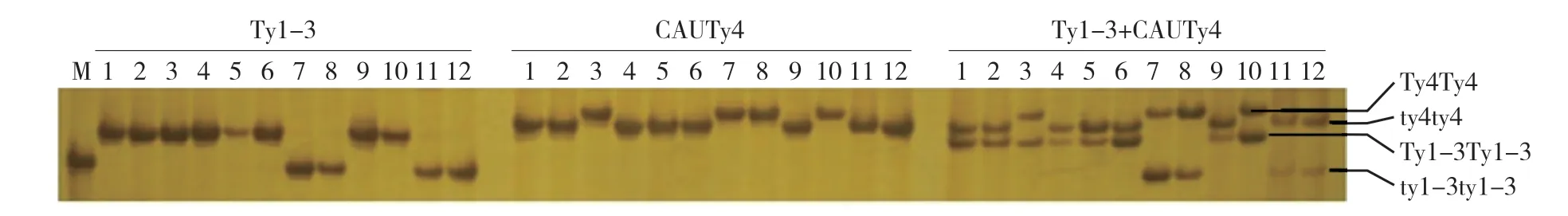
圖1 普通PCR和多重PCR的電泳對比
2.2 標記Ty1-3和CAUTy4的多重PCR體系建立
為了提高檢測效率,本試驗嘗試構建Ty1-3 和CAUTy4 這兩個標記的多重PCR 體系,在PCR 體系中同時加入兩個標記的引物,對上述12 份材料的基因組DNA 進行PCR 擴增。結果顯示(圖1),兩對引物單獨使用時都能從12 份材料中擴增出清晰的條帶,同時使用兩對引物擴增時,兩對引物的PCR 產物條帶依然清晰,表明這兩個標記可以用多重PCR 進行檢測。
2.3 不同抗性基因的聚合選擇和抗TYLCV 的鑒定結果
采用功能基因標記Ty1-3 對8個雜交組合(表3)的F1進行PCR 擴增,所有F1植株都帶有各自親本的條帶,為雜合型,表明所有單株都是真雜種,為獲得用于標記輔助選擇的F2群體提供了保障。
根據不同雜交組合所選擇基因的需要,確定了每個F2群體的播種種子數,但由于出苗等原因,有些群體沒有足夠的苗供篩選所有抗性基因位點都純合的個體,因此在實際篩選時保留少量雜合的基因型,8個組合共獲得了57個單株(表3)。
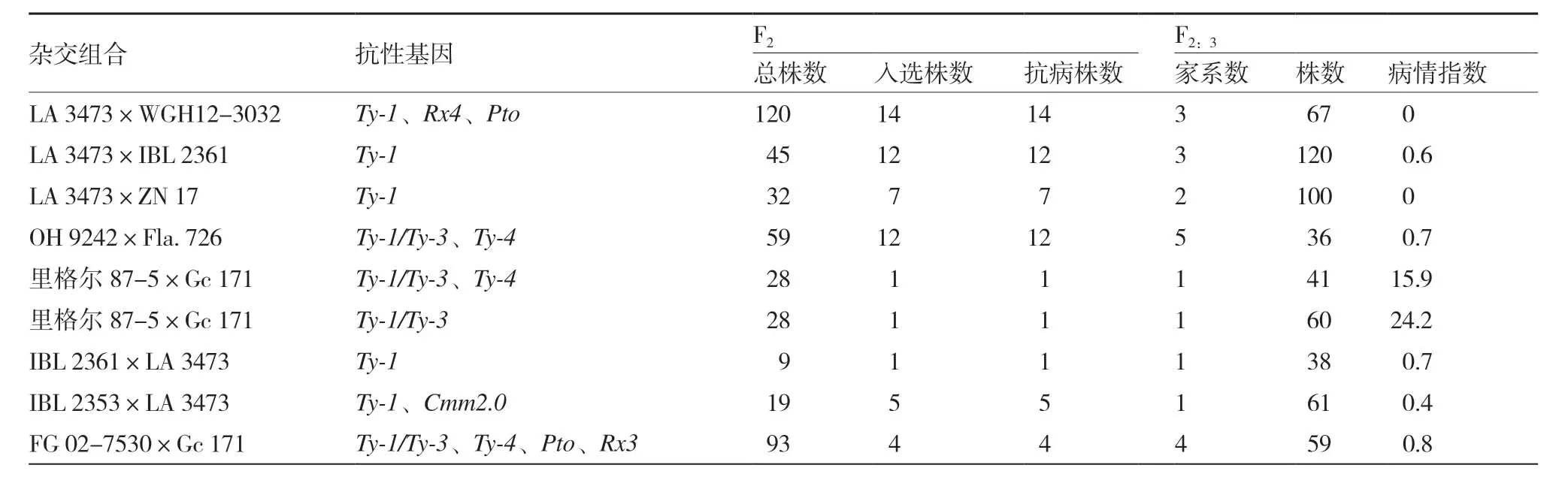
表3 各組合標記選擇獲得植株數和抗性鑒定結果
將這57個單株于2013年8月9日定植到日光溫室里,植株在生長過程中受到了煙粉虱為害,植株自然發生TYLCV 病害。調查顯示,所有單株的TYLCV 病級都在0~2 之間,表現出了較好的抗性,并能正常開花坐果,特別是從FG 02-7530×Gc 171組合中選出的4個單株,植株生長健壯,沒有任何發病癥狀(圖2-A)。而同時種植在該日光溫室里的不帶抗性基因的材料則表現出TYLCV 的典型癥狀,葉片向上卷曲,葉片黃化,植株矮小,生長極其緩慢,不能開花坐果(圖2-B)。

圖2 FG 02-7530×Gc 171 組合入選植株和不帶抗TYLCV基因的植株自然發病情況
對從F2群體中選出的部分一個位點純合而另外位點雜合的單株F2:3家系進行了標記輔助選擇,連同從F2中選出的純合單株,共得到21個F2:3家系582個單株,包括4個攜帶5個抗性基因的家系、9個攜帶3個抗性基因的家系、2個攜帶2個抗性基因的家系和6個攜帶1個抗性基因的家系(表3)。
由于接種TYLCV后的兩次調查結果基本一致,因此這里只用了接種42 d后的病級及病情指數。接種42 d后,所有單株的病級都在0~2 之間,其中0級的占88.0%,1級的占8.2%,2級的占3.8%。表明采用標記選出的所有單株對所用的病毒株系都具有抗性。
從里格爾87-5×Gc 171 組合中選出兩種類型的家系,一種只帶Ty-1/Ty-3基因,另一種攜帶Ty-1/Ty-3和Ty-4基因。前者共有60個單株,其中22個為0級,18個為1級,20個為2級,病情指數為24.2。后者共41個單株,包括16個0級,24個1級和1個2級,病情指數為15.9,而且這兩個家系的病情指數都明顯高于其他家系(表3)。
3 結論與討論
用于聚合育種的分子標記必須具有檢測的可靠性和操作的簡單性,這樣才能快速準確地選到目標基因。根據已經克隆的基因在功能區域的序列差異設計標記,是獲得較為可靠的功能基因分子標記的途徑之一。已有的抗TYLCV基因定位研究表明,抗性基因Ty-1和Ty-3都位于6號染色體上(Zamir et al.,1994;Ji et al.,2007),進一步的研究指出,Ty-1和Ty-3為等位基因(Verlaan et al.,2013)。本試驗依據已經克隆的TYLCV基因Ty-1和Ty-3與感病材料中對應基因的序列差異設計引物,建立了功能基因分子標記Ty1-3。用該標記對Zamir 等 (1994)報道的攜帶Ty-1基因的材料LA 3473 進行基因型分析,結果顯示其條帶與攜帶Ty-3材料的條帶一致,也驗證了Ty-1和Ty-3是等位基因。由這個標記選擇的單株都具有很好的抗病性,表明該標記的確為功能基因分子標記,可用于對這兩個基因的選擇。而Ji 等(2009)報道的與Ty-4基因緊密連鎖的標記是CAPs 標記,需要經過PCR、酶切等相對復雜的過程,本試驗中根據該標記在抗感材料中的序列差異設計成InDel 標記,只需要PCR 即可,簡化了檢測過程。用兩個新設計的標記篩選相應的基因,所得到的植株都表現出很好的抗性,表明這兩個新標記是可靠的。
利用不同的材料將不同的抗性基因聚合到同一個材料中,能夠增加番茄材料抗病毒生理小種的范圍,大大提高植株對病害的抗性,使其抗性更加穩定 持 久(Banerjee & Kallo,1987;Vidavski et al.,2008)。Mejía 等(2005)利用不同的番茄材料在危地馬拉進行抗病性評估,含有純合Ty-2基因型的材料在當地表現為感病,含有Ty-2和Ty-3a雜合基因型的材料表現抗病。另外,含有雜合基因型Ty-2和Ty-3a的材料比只含Ty-3純合基因型的材料表現出更高的抗性水平。在本試驗中,無論是只含有Ty-1,或者Ty-1和Ty-3,還是同時含有Ty-1、Ty-3和Ty-4的材料,對所用的病毒菌株都具有較好的抗性。但在不同的遺傳背景下抗性略有差異,當以FG 02-7530、IBL 2353、IBL 2361、OH9242、ZN 17、WGH12-3032 等為受體材料時,所得到的植株都表現出很強的抗性,而以里格爾87-5 為受體時,后代中無論是攜帶Ty-1/Ty-3基因還是同時攜帶Ty-1/Ty-3和Ty-4基因的植株,其抗性都比其他材料的略差,不過同時攜帶Ty-1/Ty-3和Ty-4基因的植株抗性要好于只攜帶Ty-1/Ty-3基因的植株。這些結果表明,遺傳背景對抗性基因的表達可能有一定的影響。
群體大小是影響多基因聚合育種效率的因素之一,聚合效率隨著群體中個體數的增加而加大,如果基礎群體太小,就有可能得不到所需要的純合基因型個體。本試驗根據每個F2群體所聚合的基因數目確定的播種量播種,但由于種子發芽率偏低(種子收獲于2012年12月),因此8個F2群體中有6個沒有達到預期的供選植株數,導致從群體中選擇所有基因位點都純合的單株數偏少,在5個聚合2~5個基因的F2群體中只有2.7%的單株在所有基因位點都表現為純合型。為了獲得更多在所有位點都純合的單株,在實際選擇時保留了一些雜合基因型個體供F2:3進一步選擇,這樣做雖然最終獲得了所有位點都純合的基因型個體,但增加了世代數和選擇成本,降低了聚合效率。因此,在進行聚合育種時,可先檢測種子的發芽率,然后結合聚合的基因數確定播種量,以便充分發揮分子標記聚合育種的優勢。
本試驗所用的番茄材料涉及到斑點病、瘡痂病、潰瘍病和TYLCV,但僅對所選到的植株進行了抗TYLCV 的鑒定。原因在于選擇番茄斑點病抗性基因Pto和瘡痂病抗性基因Rx4用的是功能基因分子標記,這兩個基因的功能分子標記已經被證實可直接用于抗性基因的選擇(Yang & Francis,2005;Pei et al.,2012)。瘡 痂 病 抗 性位點Rx3和潰瘍病抗性位點Cmm2.0是兩個尚未被克隆的QTL,但是已有的研究表明,利用與Rx3位點緊密連鎖的標記可準確選擇到攜帶該位點的抗性單株(Yang & Francis,2005;張曉敏 等,2009),而利用與Cmm2.0緊密連鎖的標記也可準確獲得抗潰瘍病的單株(Coaker & Francis,2004)。鑒于本試驗所用的抗性材料和分子標記與文獻中(Coaker & Francis,2004;Yang & Francis,2005;張曉敏 等,2009;Pei et al.,2012)的相同,因此推測本試驗所選的攜帶抗性基因的材料即為抗病材料。
綜上所述,本試驗開發了抗TYLCV基因Ty-1/Ty-3和Ty-4的新標記,并建立了多重PCR 體系。通過雜交結合標記輔助選擇,將抗TYLCV、斑點病、瘡痂病、潰瘍病等的基因分別聚合,獲得了7份兼抗斑點病、TYLCV 和瘡痂病的材料,6 份攜帶多個TYLCV基因(Ty-1/Ty-3和Ty-4)的材料,以及1 份兼抗TYLCV 和潰瘍病的材料,為番茄抗多個病害分子標記聚合育種提供了材料。
杜永臣,嚴準,王孝宣,李樹德,朱德蔚.1999.番茄育種研究主要進展—文獻綜述.園藝學報,26(3):161-169.
李常保,崔彥玲,張麗英,李傳友.2012.番茄黃化曲葉病毒的快速分子檢測.遺傳,34(3):366-370.
李小靖,葉志彪.2010.我國番茄黃化曲葉病發生規律和研究進 展.長江蔬菜,(2):1-5.
葉青靜,楊悅儉,王榮青,李志邈,阮美穎,周國治,姚祝平.2009.番茄抗黃化曲葉病育種研究進展.中國農業科學,42 (4):1230-1242.
吳篆芳,曹坳程,鄭建秋,郭美霞,王秋霞,李園,顏冬冬,毛連綱,馬濤濤.2013.番茄黃化曲葉病毒病研究進展.作物雜志,(1):18-23.
楊曉慧,國艷梅,王孝宣,高建昌,杜永臣.2012.番茄抗黃化曲葉病基因與基因工程研究最新進展.園藝學報,39(11):2283-2290.
張曉敏,Francis D M,楊文才.2009.我國部分番茄主栽品種抗瘡痂病評價和標記輔助選擇.華北農學報,24(4):183-187.
Banerjee M K,Kallo O.1987.Inheritance oftomato leaf curl virusresistance inLycopersicon hirsutumf.glabratum.Euphytica,36:581-584.
Coaker G L,Francis D M.2004.Mapping,genetic effects,and epistatic interaction of two bacterial canker resistance QTLs fromLycopersicon hirsutum.Theor Appl Genet,108:1047-1055.
Cohen S,Antignus Y.1994.Tomato yellow leaf curl virus(TYLCV),a whitefly-borne geminivirus of tomatoes.Adv Dis Vector Res,10:259-288.
Friedmann M,Lapidot M,Cohen S,Pilowsky M.1998.A novel source of resistance totomato yellow leaf curl virusexhibiting a symptomless reaction to viral infection.J Amer Soc Hort Sci,123 (6):1004-1007.
Horowitz R,Denholm I,Morin S.2007.Resistance to insecticides in the TYLCV vector,Bemisia tabaci// Czosnek H.Tomato yellow leaf curl virusdisease.Netherlands:Springer:305-325.
Ji Y,Schuster D J,Scott J W.2007.Ty-3,a begomovirus resistance locus near thetomato yellow leaf curl virusresistance locusTy-1on chromosome 6 of tomato.Mol Breed,20:271-284.
Ji Y,Scott J W,Schuster D J.2009.Molecular mapping ofTy-4,a newtomato yellow leaf curlvirusresistance locus on chromosome 3 of tomato.J Amer Soc Hort Sci,134:281-288.
Lapidot M,Friedmann M.2002.Breeding for resistance to whitefly- transmitted geminiviruses.Ann Appl Biol,140:109-127.
Mejía L,Teni R E,Vidavski F,Czosnek H,Lapidot M,Nakhla M K,Maxwell D P.2005.Evaluation of tomato germplasm and selection of breeding lines for resistance to begomoviruses in Guatemala.Acta Hortic,695:251-255.
Morales F J.2001.Conventional breeding for resistance toBemisia tabacitransmitted geminiviruses.Crop Protect,20:825-834.
Palumbo J C,Horowitz A R,Prabhaker N.2001.Insecticidal control and resistance management forBemisia tabaci.Crop Protect,20:739-766.
Pei C C,Wang H,Zhang J Y,Wang Y Y,Francis D M,Yang W C.2012.Fine mapping and analysis of a candidate gene in tomato accession PI128216 conferring hypersensitive resistance to bacterial spot race T3.Theor Appl Genet,124:533-542.
Sun W Y,Zhao W Y,Wang Y Y,Pei C C,Yang W C.2011.Natural variation ofPtoandFengenes and marker-assisted selection for resistance to bacterial speck in tomato.Agric Sci China,10 (6):827-837.
Verlaan M G,Hutton S F,Ibrahem R M,Kormelink R,Visser R G F,Scott J W,Edwards J D,Bai Y L.2013.Thetomato yellow leaf curl virusresistance genesTy-1andTy-3are allelic and code for DFDGD class RNA dependent RNA polymerases.PLoS Genet,9(3):e1003399.
Vidavski F,Czosnek H,Gazit S,Levy D,Lapidot M.2008.Pyramiding of genes conferring resistance totomato yellow leaf curl virusfrom different wild tomato species.Plant Breed,127(6):625-631.
Yang W C,Francis D M.2005.Marker-assisted selection for combining resistance to bacterial spot and bacterial speck in tomato.J Amer Soc Hort Sci,130:716-721.
Zamir D,Ekstein M I,Zakay Y,Navot N,Zeidan M,Sarfatti M,Eshed Y,Harel E,Pleban T,van-Oss H,Kedar N,Rabinowitch H D,Czosnek H.1994.Mapping and introgression of atomato yellow leaf curlvirustolerance gene,Ty-1.Theor Appl Genet,88:141-146.
