環境溫度和鹽度對墨吉明對蝦(Fenneropenaeus merguiensis)胚胎發育的影響*
2014-03-09 06:57:38楊世平王成桂黃海立陳兆明孫成波
海洋與湖沼 2014年4期
楊世平 王成桂 黃海立 陳兆明 孫成波
(廣東海洋大學水產學院 湛江 524088)
生物的早期發育階段是復雜的生命周期中最為敏感的階段。其中,鹽度和溫度是水生生物自然水體分布范圍的主要決定因子,影響著水生生物的生長與繁殖,尤其是影響水生生物胚胎發育的最重要的非生物因素。因此,學者們分別研究了鹽度或溫度對方斑東風螺(Babylonia areolata)、尖紫蛤(Soletellina acuta)、長毛明對蝦(Fenneropenaeus penicillatus)等胚胎發育的影響(施流章,1981;劉建勇等,2005;黃洋等,2011)。不同溫度和鹽度對凡納濱對蝦(Litopenaeus vannamei)受精卵孵化出膜的時間和受精率有顯著影響,并得出的理想孵化水溫為 28—30°C,鹽度為27.93—33.01(彭昌迪等,2002)。因此,在進行水生生物苗種的大規模生產時,必須要關注和了解鹽度和溫度等物理性質,以及該物種對這些性質的要求和適應能力。
墨吉明對蝦(Fenneropenaeus merguiensis)屬于十足目(Decapoda)、枝鰓亞目(Dendrobranchiata)、對蝦科(Penaeidae)、明對蝦屬(Fenneropenaeus),廣泛分布于我國東南沿海,以及亞洲和澳洲的熱帶和亞熱帶海域中。它具有個體大、生長快、肉質細嫩、滋味鮮美的特點,我國很早就對其進行了人工養殖。吳琴瑟(1975)就針對墨吉明對蝦的產卵、孵化、胚胎發育、幼體培育等進行了研究。吳琴瑟等(1985)還研究了墨吉明對蝦育苗期疾病的藥物防治技術。彭文國等(1996)研究了pH值、鹽度、氨氮濃度變動情況下幼體的耗氧量的變化。近年來,由于凡納濱對蝦進口親蝦的價格節節攀升、而養殖成功率卻不斷下降,嚴重影響對蝦養殖效益。廣東省湛江對蝦種苗協會建議開展墨吉明對蝦的人工繁殖和養殖技術研究,以期能替代部分凡納濱對蝦的養殖,優化我國的對蝦養殖結構。Hoang等(2002a)研究了墨吉明對蝦在池塘養殖條件生長、性腺成熟和產卵。國外學者還對其性腺催熟、產卵條件、病毒感染、小規模人工育苗等開展了相關的研究(Hoanget al,2002b;Naiket al,2002;Wanget al,2002;Zachariaet al,2002,2004b),以期能實現墨吉明對蝦的規模化養殖。有關溫度和鹽度對墨吉明對蝦胚胎發育影響也有報道,但其研究的溫度、鹽度范圍較窄。因此,本文對溫度和鹽度對墨吉明對蝦胚胎發育的影響進行研究,為開展墨吉明對蝦的人工繁殖、育種等研究提供理論依據,促進其養殖業的健康發展。
1 材料與方法
1.1 親蝦來源與用水
本實驗所用墨吉明對蝦為湛江海區采捕自然成熟的親蝦,挑選個體大、性腺飽滿的雌蝦。自然海水經沉淀、砂濾,強氯精消毒、曝氣后,再用 5×10–6的ETDA二鈉鹽消除重金屬離子。
1.2 胚胎發育觀察
將親蝦放入 1m3的黑色產卵桶中讓其自然產卵,產卵桶上加蓋黑色塑料膜遮擋光線。每10min觀察桶內產卵情況,一旦發現產卵,立即移走親蝦。在水溫為27.5—27.8°C(自然水溫)、鹽度為28、微沸騰的充氣環境下進行孵化。從發現產卵開始,根據胚胎發育情況隨時用吸管吸取受精卵或胚胎在顯微鏡下觀察,記錄胚胎不同發育階段的特征,并對不同發育階段的胚胎進行拍照,直至幼體出膜。
1.3 溫度對胚胎發育的影響
取受精卵置于事先準備好的裝有1L干凈海水的2L 塑料桶中,置于 23°C、26°C、29°C、32°C、35°C溫度下孵化。每個溫度設3個平行組,每組100粒受精卵,鹽度均為 28。為了保證受精卵能懸浮在水中,每個實驗組均進行充氣,充氣量為微沸騰狀態。分卵的過程在10min內完成,以減少受精卵在原有溫度條件下胚胎發育的影響。觀察統計在不同溫度條件下受精卵的孵化率和幼體出膜時間,為保證各平行組胚胎的充分孵化時間,在觀察到幼體出膜 2h后進行孵化率的統計。溫度實驗控溫裝置采用白色泡沫箱內裝淡水水浴的方式進行,高溫組(29°C,32°C,35°C)采用缸內電子恒溫器進行加熱,低溫組(23°C,26°C)采用冰袋降溫,所有實驗桶均放置溫度計,以保證控溫的準確性。
1.4 鹽度對胚胎發育的影響
受精卵在裝有1L干凈海水的2L塑料桶中進行孵化。鹽度實驗組也設 5個梯度,依次為15、20、25、30、35,溫度為 27.0—27.5°C(小桶內的自然水溫)。每個鹽度梯度設有3個平行組。保證微沸騰的充氣量,分卵的過程在10min內完成。觀察統計在不同溫度條件下的孵化率和幼體出膜時間。為保證每個平行組胚胎的充分孵化時間,在觀察到幼體出膜 2h后進行孵化率的統計。高鹽度組(30,35)采用海水精進行調節,低鹽度組(15,20,25)采用經消毒、暴氣處理的自來水進行調節,鹽度采用手持式折光鹽度計進行測定。
1.5 主要觀測指標及數據處理
孵化時間是指從受精卵開始發育到幼體出膜所經歷的時間。孵化率是指受精卵經孵化后出膜的幼體總數與受精卵總數的比率。最適孵化溫度(鹽度)范圍是指孵化率超過 80%,幼蟲孵化出膜后活力正常的實驗組的溫度(鹽度)范圍。所得數據均用平均值±標準差(mean±SD)表示,用Excel 2003和SPSS13.0軟件對數據進行擬合回歸方程和單因素方差分析(ANOVA)和多重比較進行統計分析,并在P<0.05水平上對結果進行差異顯著性檢驗。
2 實驗結果
2.1 墨吉明對蝦的胚胎發育過程
受精卵 墨吉明對蝦的受精卵,呈橢圓,直徑為280—300μm。在產出10min后,見突起的卵膜,同時見到第1次和第2次成熟分裂時排出的第1極體、第2極體以及多余的精子(圖1a)。
卵裂 受精卵發生正常的完全均等卵裂。第1、2次卵裂產生的分裂球,可在同一平面觀察到,其均等性特別顯著(圖1b,圖1c)。第3次卵裂后產生8個基本等大的細胞,因其呈現明顯的螺旋卵裂特征,在顯微鏡能同時觀察到6個細胞(圖1d)。在以后的第4、5、6次卵裂分別產生16、32和64細胞,從顯微鏡下可以觀察到的胚胎邊緣的細胞來看,其卵裂球的大小基本一致(圖1e,圖1f,圖1g)。從第1次分裂產生2細胞時起至第6次分裂產生64細胞,平均每次分裂的時間為14.4min。
囊胚期 從第7次卵裂開始,卵裂后細胞形成一個完全封閉的囊胚腔,胚胎進入囊胚期,此期胚胎仍為圓形,在顯微鏡下能觀察胚胎邊緣由細胞分裂產生的分裂球(圖1h)。
原腸期 胚胎的植物極端稍變扁平,其他細胞開始內陷,原腸腔逐漸形成。在顯微鏡下不能明顯觀察到胚胎邊緣的分裂球,可明顯觀察到內陷處的細胞分裂球(圖1i)。
肢芽期 原腸胚繼續發育,4h 33min時,觀察到胚胎兩側呈現3對肢芽凸起,說明胚胎發育進入肢芽期。在顯微鏡下觀察不到胚胎細胞的界線。在該期可清楚看到胚胎外包裹的膜(圖1j)。肢芽期發育時間較長,從不同角度觀察胚胎的形態差異較大。肢芽期的發育時間約2h 42min。

圖1 墨吉明對蝦胚胎發育各期的形態Fig.1 The morphologies of F.merguiensis embryos in each development stage
膜內無節幼體 7h 15min時,觀察到胚胎的3對附肢末端出現刺狀剛毛,標志著胚胎發育進入膜內無節幼體期(圖1k)。隨著胚胎的發育,頭部中央出現了1個棕色眼點(圖1l)。發育到10h 59min時,觀察膜內無節幼體的附肢能抖動。在11h 50min時,幼體破膜而出。膜內幼體發育時間約4h 35min。
2.2 溫度對墨吉明對蝦胚胎發育的影響
從表1可以看出,水溫對墨吉明對蝦的幼體出膜時間和孵化率有顯著的影響(P<0.05)。水溫高于35°C的實驗組,胚胎發育至囊胚期后陸續死亡,少數胚胎發育到膜內無節幼體期,但不能出膜。水溫在 23—32°C之間胚胎能發育至膜內無節幼體期,且幼體能順利出膜。水溫為 32°C的實驗組,幼體出膜時間最短555min,孵化率為92.67%±1.53%,幼體活力也正常。水溫為 23°C的實驗組,幼體出膜時間最長(1108.00±49.11)min,比水溫為 32°C的實驗組的幼體出膜時間增加了 553min,且各平行組間幼體出膜時間差較大,達96min,孵化率也僅為65.33%±5.51%。26°C組孵化率最高為 96.67%±1.15%,但 26°C、29°C和 32°C 組孵化率比較接近,各組間差異不顯著(P>0.05)。從孵化率來看,墨吉明對蝦胚胎發育在最適溫度范圍為26—32°C。

表1 不同水溫條件下墨吉明對蝦胚胎的發育情況Tab.1 Embryonic development of F.merguiensis at different temperatures
在23—32°C 的范圍內,胚胎能正常發育,幼體出膜時間與水溫之間呈負相關關系,孵化水溫(x)與幼體出膜時間(y)存在著多項式函數y= 4.1111x2–289.13x+ 5590.4(R2= 0.9946)。在最適水溫范圍內26—32°C,受精卵的孵化率與水溫之間呈負相關關系。在實驗水溫范圍內,孵化水溫(x)與孵化率(y)也存在著多項式函數y= –1.9683x2+ 109.67x–1419.8(R2= 0.9188)(圖2)。
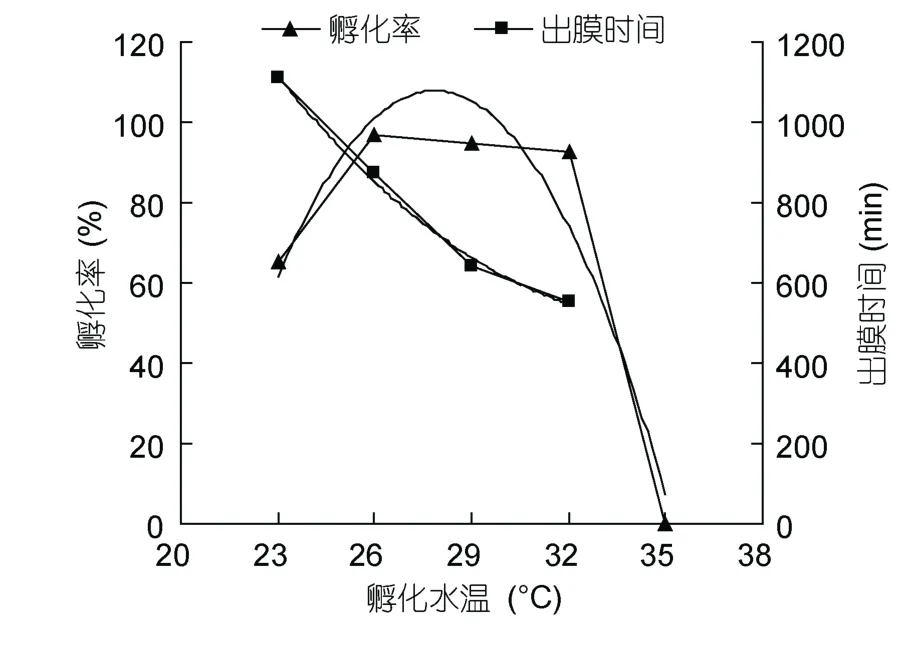
圖2 墨吉明對蝦幼體出膜時間、孵化率與水溫的關系Fig.2 Relationship between temperature and the hatching time,hatching ratio of F.merguiensis
2.3 鹽度對胚胎發育的影響
從表2可以看出鹽度對墨吉明對蝦的孵化率有顯著的影響(P<0.05)。鹽度為15的實驗組少數胚胎發育到囊胚期死亡,大多數胚胎發育至肢芽期后死亡。鹽度在20—35之間胚胎均能發育至膜內無節幼體期,且幼體能順利出膜。鹽度為20組的胚胎發育最慢,幼體出膜時間為960min,顯著慢于鹽度為25、30和35的實驗組(P<0.05),且孵化率也僅為 57.67%±7.09%,顯著低于鹽度為25、30和35的實驗組(P<0.05)。在鹽度為25、30和35的實驗組,其幼體出膜時間相當,同時觀察到幼體出膜,且孵化率均在90%以上。表明墨吉明對蝦胚胎發育在最適鹽度范圍為 25—35。統計結果表明,在鹽度為25、30和35的實驗組的幼體出膜時間和孵化率均無顯著差異(P>0.05),說明在最適鹽度范圍內,鹽度對墨吉明對蝦的胚胎發育沒有顯著的影響。
在鹽度為 20—35的范圍內,鹽度(x)與幼體出膜時間(y)存在著多項式函數y= 0.84x2–51.24x+ 1574.6(R2= 0.9333)。在實驗鹽度范圍內,鹽度(x)與孵化率(y)也存在著多項式函數y=–0.4438x2+ 26.537x–296.59(R2= 0.9764)(圖3)。

表2 不同鹽度條件下墨吉明對蝦胚胎的發育情況Tab.2 Embryonic development of F.merguiensis at different salinity
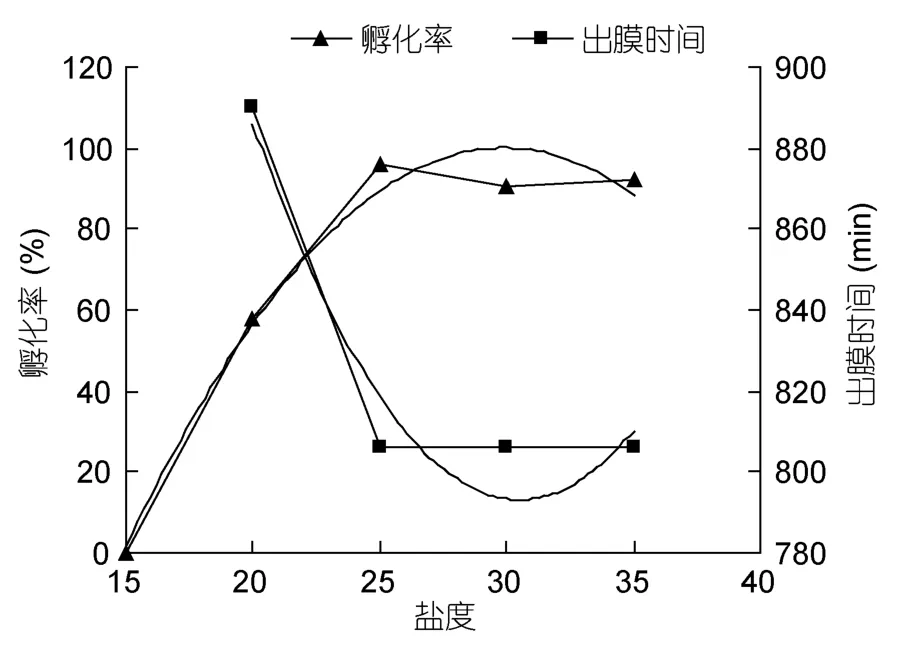
圖3 墨吉明對蝦幼體出膜時間、孵化率與鹽度的關系Fig.3 Relationship between salinity and the hatching time,hatching ratio of F.merguiensis
3 討論
3.1 溫度對墨吉明對蝦的胚胎發育
本實驗結果表明,墨吉明對蝦胚胎發育最適溫度范圍為 26—32°C。且在 23—32°C 的范圍內,胚胎能正常發育,幼體出膜時間與水溫之間呈負相關關系。與吳琴瑟(1975)報道結果相似,該作者認為在適宜的水溫范圍,墨吉明對蝦受精卵的發育速度隨溫度升高而加速其胚胎發育適宜水溫為23—32°C。說明水溫對墨吉明對蝦胚胎發育速度快慢的影響是非常顯著的。在對長毛明對蝦胚胎發育研究中,也發現溫度對幼體出膜時間有影響,在一定的水溫范圍內,溫度越高,發育越快,出膜時間越短。在鹽度為28.59,水溫為(28±0.5)°C時的孵化時間為13h 20min,而水溫為(24±0.5)°C 的孵化時間為 16h 50min(施流章,1981)。溫度對青蝦(Macrobrachium nipponense)、尖紫蛤、方斑東風螺等的胚胎發育的影響也呈現相似的規律(鄒盛希等,2002;劉建勇等,2005;黃洋等,2011)。
從實驗結果可以看出:溫度既影響墨吉明對蝦胚胎發育的速度,也影響了受精卵的孵化率。水溫29.0°C時,墨吉明對蝦的幼體出膜時間為641min,與凡納濱對蝦(11.8h)相近(彭昌迪等,2002),它們都具有熱帶型對蝦的共同點。在 26—32°C時,其孵化率均為 90%以上,而在溫度為 23°C時,其孵化率僅為65.33%,說明作為熱帶和亞熱帶品種的墨吉明對蝦期胚胎發育的溫度要求較高。溫度高可縮短孵化時間,但其孵化率為受到影響,如果進一步提高了溫度,則會引起胚胎畸形,產生敗育(陳昆慈等,2008;黃洋等,2011)。溫度過低時(23°C),墨吉明對蝦的孵化率顯著低于最適溫度內各組的孵化率(P<0.05),說明溫度過低,胚胎發育會產生滯育。
3.2 鹽度對墨吉明對蝦的胚胎發育
Roberts(1971)報道,每一個物種都有一個獨特的鹽度范圍適合胚胎發育和孵化。從實驗結果來看,鹽度也會影響墨吉明對蝦胚胎發育的速度和受精卵的孵化率。鹽度為 15時,其胚胎不能正常發育和孵化。在鹽度為 20時,幼體的出膜時間為 960min,其孵化率也僅為 57.67%,孵化效果明顯差于鹽度為 25、30和35的實驗組。但在鹽度為25、30和35的各實驗組之間,其幼體出膜時間和孵化率均沒有顯著差異(P>0.05),說明墨吉明對蝦胚胎發育的鹽度范圍較廣。分布和形態結構相似的長毛明對蝦胚胎發育時,同一溫度條件下,各鹽度組幼體的出膜時間相差不大(施流章,1981)。凡納濱對蝦在鹽度為25.39—33.01范圍內,最高孵化率為96.4%(鹽度為27.93),但兩端孵化率低于墨吉明對蝦,分別為76.7%和82.4%(彭昌迪等,2002)。說明墨吉明對蝦胚胎發育的適宜鹽度比凡納濱對蝦寬,孵化率也較高。本實驗的結果顯示,在鹽度為 25時,孵化率為 96%,是各鹽度組中最高的。而 Zacharia等(2004a)在鹽度為 35,溫度為 33°C和29°C時,取得了墨吉明對蝦較高的孵化率(分別為86.5%和 86.5%)。與本實驗的結果存在一定的差異,是否與墨吉明對蝦親蝦來源不同有關,還有待進一步研究。水溫和鹽度對墨吉明對蝦胚胎發育速度快慢的都有顯著的影響,但從水溫和鹽度二者來比較,影響發育速度快慢的主要因素還是溫度。
致謝在實驗過程中得到馮楚嶂、陳旭東和何炳強同學的幫助,謹致謝忱。
劉建勇,卓健輝,2005.溫度和鹽度對方斑東風螺胚胎發育的影響.湛江海洋大學學報,25(1):1—4
吳琴瑟,1975.墨吉對蝦人工育苗.水產與教育,2:42—47
吳琴瑟,謝永康,楊燕忠等,1985.藥物防治墨吉對蝦育苗期疾病提高成活率的試驗.福建水產,(4):5—10
鄒盛希,李永吉,2002.溫度對青蝦胚胎及幼體發育影響的初步觀察.內陸水產,27(4):15—15
陳昆慈,朱新平,杜合軍等,2007.溫度和鹽度對寶石鱸胚胎發育的影響.中國水產科學,14(6):1032—1037
施流章,1981.溫、鹽度與長毛對蝦卵的孵化及無節幼體發育的關系.水產學報,5(1):57—63
黃 洋,黃海立,呂廣煊等,2011.溫度和鹽度分別對尖紫蛤胚胎發育的影響.海洋科學,35(10):117—122
彭文國,鄭建民,1996.墨吉對蝦幼體耗氧量的初步研究.水產科技情報,23(4):169—172
彭昌迪,鄭建民,彭國文等,2002.南美白對蝦的胚胎發育以及溫度與鹽度對胚胎發育的影響.上海水產大學學報,11(4):310—316
Hoang T,Lee S Y,Keenan C Pet al,2002a.Observations on growth,sexual maturity and spawning performance of pond-rearedPenaeus merguiensis.Aquaculture Research,33(11):863—873
Hoang T,Lee S Y,Keenan C Pet al,2002b.Spawning behaviour ofPenaeus(Fenneropenaeus)merguiensisde Man and the effect of light intensity on spawning.Aquaculture Research,33(5):351—357
Naik S D,Belsare S G,Sharangdhar M Tet al,2002.Seed production trial of banana prawn(Fenneropenaeus merguiensis)on small scale in indoor hatchery.Journal of Maharashtra Agricultural Universities(India),27(3):296—299
Roberts M H,1971.Larval development ofPagurus longicarpusSay reared in the laboratory.II.Effects of reduced salinity on larval development.The Biological Bulletin,140(1):104—116
Wang Y T,Liu W,Seah J Net al,2002.White spot syndrome virus(WSSV)infects specific hemocytes of the shrimpPenaeus merguiensis.Diseases of Aquatic Organisms,52(3):249—259
Zacharia S,Kakati V S,2002.Effect of eyestalk ablation on ovarian maturation in the banana prawn,Fenneropenaeus merguiensisde man under different environmental conditions.Journal of the Marine Biological Association of India,45(1):111—114
Zacharia S,Kakati V S,2004a.Optimal salinity and temperature for early developmental stages ofPenaeus merguiensisDe man.Aquaculture,232(1):373—382
Zacharia S,Kakati V S,2004b.Ovarian maturation in the banana shrimp,Fenneropenaeus merguiensis(De Man)by Serotonin(5-hydroxytryptamine)injection.Indian Journal of Fisheries,51(1):11—15
