大蔥CMS 58不育系及保持系花藥和花粉的顯微觀察
2014-03-11 08:35:32*
中國蔬菜 2014年10期
*
(1北京市農林科學院蔬菜研究中心,農業部華北地區園藝作物生物學與種質創制重點實驗室, 北京100097;2山西大學生命科學學院, 山西太原 030005)
大蔥CMS 58不育系及保持系花藥和花粉的顯微觀察
王 嬋1,2趙 泓1劉倩純1裴雁曦2*王永勤1*
(1北京市農林科學院蔬菜研究中心,農業部華北地區園藝作物生物學與種質創制重點實驗室, 北京100097;2山西大學生命科學學院, 山西太原 030005)
利用石蠟切片技術對大蔥細胞質雄性不育系 58 A1、58 A2及其保持系58 B的花藥發育過程進行了比較,并利用掃描電鏡技術觀察了3個樣品的花粉粒。石蠟切片結果顯示:大蔥不育系58 A1花藥絨氈層提前解體,影響了正常小孢子的形成,導致花粉敗育;而58 A2的花藥絨氈層異常增厚,擠壓藥室內的小孢子,小孢子內的物質流失,導致花粉敗育。58 A2藥室內壁細胞細胞壁沒有加厚,花藥同側的2個花粉囊之間的細胞層極厚,可能是花藥成熟后無法開裂的主要原因。掃描電鏡結果顯示:花粉粒呈長球形或超長球形,具單溝,外壁為腦紋狀雕紋,不育系的成熟花粉粒很少或多數為干癟花粉粒。
大蔥;CMS;花藥;花粉;石蠟切片;掃描電鏡
大蔥(Allium fistulosum L.)是蔥科蔥屬中二、三年生草本植物,廣泛栽培于亞洲東部的中國、韓國和日本(Inden & Asahira, 1990),在我國最少有2 600年的栽培歷史(崔連偉 等,2004),常常作為一種綠色保健蔬菜、調味品。植物細胞質雄性不育(cytoplasmic male sterility,CMS)廣泛存在于高等植物中, 表現為雄性器官不能形成有活力的花粉,而雌性器官的發育和植株的營養生長正常, 其遺傳方式是母性遺傳。目前已經在300多種植物中發現雄性不育現象(Kaul, 1988)。由于CMS可免除人工去雄的復雜操作,因此是雜種種子生產的重要途徑之一,為此,許多學者將研究內容集中在植物雄性不育機理上。
高等植物的花粉粒由花粉母細胞經過減數分裂后發育形成。花粉母細胞減數分裂形成四分體,隨著胼胝質壁的溶解,四分體的4個子細胞彼此分離,成為4個各含有1個細胞核的小孢子(也稱作單核花粉粒)。小孢子經過分裂、生長發育為成熟的花粉粒(吳鴻和郝剛,2012)。Laser和Lesrten (1972)統計62例CMS花粉敗育時期后發現雙子葉植物在花粉造孢細胞至四分體時期敗育較多,而單子葉植物中不育花粉敗育的高峰出現在單核期至雙核期;同時也有學者認為花粉敗育與絨氈層及中層細胞的異常行為有關。另外對花粉超微結構的觀察也揭示了植物CMS敗育的機理。目前已經有學者在細胞學水平上研究了大蔥不育系敗育的原因。席湘媛(1991)曾指出大蔥花粉敗育發生在單核小孢子期的早期,王曉靜等(2007)也對大蔥不育機理進行了研究,總結出敗育有兩種類型:中層延遲解體和絨氈層提前解體;席湘媛(2000)還對大蔥的花粉進行了超微結構的觀察,總結出了發育過程中脂滴、各種細胞器等的變化情況。
大蔥CMS 58有兩個不育系58 A1和58 A2,兩者敗育的表型不同:前者為花藥提前開裂,后者表現為花藥褐化。為了探究這兩個不育系的敗育類型,本試驗對兩者及其保持系的花藥進行了石蠟切片觀察,且對花藥和花粉進行了掃描電鏡觀察,以期為大蔥育種中CMS 58的利用提供理論指導。
1 材料與方法
1.1 試驗材料
供試材料為已經選育成的穩定遺傳的大蔥不育系58 A1、58 A2及保持系58 B,2012年6月種植于北京市農林科學院蔬菜研究中心試驗地,于2013年開花前采集不同大小的花蕾進行顯微觀察。
1.2 試驗方法
1.2.1 利用壓片法確定花藥發育時期 用醋酸洋紅染色法壓片觀察花藥,根據花粉發育的一些明顯特征來確定花藥發育的時期,從而確定四分體時期的花蕾大小。
1.2.2 石蠟切片的制備與觀察(蘇立娟,2005;Li et al., 2006;王原媛 等,2010) 采集不同發育時期的可育及不育花蕾,在FAA固定液中固定2~24 h。不同濃度的乙醇依次脫水后,用二甲苯透明,用石蠟浸蠟及包埋,輪轉式切片機切片,切片厚度為6 μm。展片與烤片之后用二甲苯脫蠟,乙醇梯度脫水,蘇木精染色后用中性樹膠封片, OLYMPUS顯微鏡觀察并照相。
1.2.3 掃描電鏡樣品的制備與觀察(曲波 等,2006) 取上述各樣品的成熟花藥,置于防塵處干燥,在解剖鏡下將花粉撒到樣品托的雙面膠帶上,在IB-5離子濺射儀中真空噴金鍍膜,在日立s-3400N型掃描電鏡下觀察并拍照。每個樣品測量20粒花粉的極軸長和赤道軸長,取平均值。同時,直接將各樣品的成熟花藥撒到樣品托的雙面膠帶上,同樣對其進行掃描電鏡觀察。掃描電鏡在北京農業生物技術研究中心完成。
2 結果與分析
2.1 大蔥花藥發育時期的確定
利用普通光學顯微鏡,通過醋酸洋紅染色法對采集到的花蕾進行花粉發育時期的觀察。在對花粉發育的各個時期進行全面觀察后發現,四分體時期是最容易觀察和識別的時期,即使不使用任何染液,也可以很輕易地正確識別。為了提高工作效率,對處于四分體時期的30個花蕾的形態特征進行了統計。處于四分體時期的花蕾的平均長度、直徑和柄長分別為(2.40±0.15) mm、(1.46±0.21) mm和(3.73±1.84 )mm。經多次反復驗證,將花蕾長度作為判定小孢子發育的四分體時期較為可靠。以此長度為分界線,花蕾長度約為3.4 mm和5.4 mm時可分別收集到單核花粉粒和成熟花粉粒時期,花蕾約為2.2 mm時可收集到花粉母細胞發育的各個時期。由此,在田間通過測量花蕾的大小就可以估算出其花藥發育的大致時期,為后續試驗樣品的采集奠定了基礎。
2.2 大蔥雄性不育系及其保持系花藥發育過程
通過石蠟切片觀察大蔥保持系58 B的花藥,發現每個花蕾中有6個花藥,花藥橫切面似蝴蝶形。每個花藥均由中央的一個藥隔及周圍的4個花粉囊組成的。4個花粉囊分為左右兩半,中間由藥隔相連,每個花粉囊由花粉囊壁和花粉囊室(藥室)組成。在大蔥6個花藥外圍有兩輪花被,每輪3片(圖1)。花粉囊壁由外至內分別由表皮、藥室內壁、中層及絨氈層四部分組成。

圖1大蔥單個花蕾的橫切圖
處于減數分裂時期的花粉母細胞,細胞體積較大,核內物質濃厚,染色極深。此時期中層細胞不發達,絨氈層細胞大,細胞質濃厚,有規則地排列在小孢子母細胞的一周(圖2-A)。小孢子母細胞經減數分裂后形成四分體,四分體被透明的胼胝質壁包圍,此時絨氈層細胞仍整齊排列,但部分中層細胞已經開始解體(圖2-B)。隨后,四分體細胞外的胼胝質被絨氈層細胞分泌的胼胝質酶分解,釋放出小孢子,絨氈層細胞開始解體(圖2-C)。此后小孢子經雙核期、三核期后發育為呈卵圓形的成熟花粉粒,同時伴隨著絨氈層和中層細胞的消失。藥室內壁細胞內壁縱向條紋狀加厚,但在花藥同側上下2個花粉囊間的若干細胞壁并未加厚(圖2-D)。當花粉粒完全成熟時,暴露在空氣中的花藥失水,纖維層收縮,花藥由此處開裂,散出花粉粒。這就是晴天有利于花藥散粉,而陰雨天時花藥不容易散粉的原因。
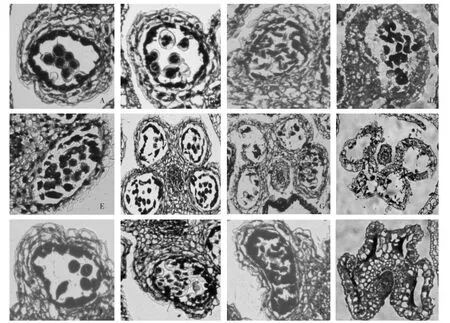
圖2大蔥不育系及保持系花藥發育的顯微結構
不育系58 A1的花藥會提前開裂,表現出不育性。對其花藥發育進行觀察,發現在小孢子母細胞減數分裂時期,部分絨氈層已經開始提前解體 (圖2-E),四分體時期藥室內側環狀的絨氈層細胞結構已經消失,藥室中可以見到絨氈層細胞的斷片。由于絨氈層細胞的提前解體,可以明顯地觀察到扁平的中層細胞(圖2-F)。小孢子時期絨氈層完全消失,部分小孢子細胞胞質外流,呈透明不規則球形(圖2-G)。到花粉粒成熟期,藥室內成熟的花粉粒極少,藥室內壁細胞細胞壁加厚,出現特有的縱向條狀,花藥同側的2個花粉囊之間的細胞層較厚(圖2-H)。
不育系58 A2的花藥在發育過程中會褐化,表現出不育性。對其花藥發育進行觀察,與保持系的相比,其小孢子母細胞時期及四分體時期,組成花粉囊壁的4層細胞沒有明顯異常,絨氈層細胞一直保持環狀結構,細胞內胞質濃厚,染色極深,與保持系沒有明顯差別 (圖2-I、J)。但是在小孢子時期,本應開始程序性死亡的絨氈層細胞異常增厚,中層細胞略有增厚,花粉囊向內萎縮,單個藥室酷似鞋狀(圖2-K)。中層細胞明顯增厚,花粉囊繼續向內萎縮,小孢子被擠壓成一條染色極深的團塊狀物質而無法發育成花粉粒。藥室內壁細胞細胞壁沒有加厚,花藥同側的2個花粉囊之間的細胞層極厚(圖2-L)。這種結構是58 A2的花藥成熟后無法開裂的主要原因。
2.3 大蔥雄性不育系及保持系花粉粒特征
大蔥保持系58 B成熟花藥的掃描電鏡可看出許多成熟花粉粒從花藥藥室中散出(圖3-A)。花粉粒為超長球形,赤道面觀為窄橢圓形,極面觀為一裂圓形,極赤比(P/E)為2.48,具有單溝,溝長達到兩極(圖3-B)。花粉粒外壁具有腦紋狀雕飾,表面多孔穴和坑(圖3-C)。
大蔥不育系58 A1的成熟花藥中無明顯的花粉粒(圖3-D)。極少數的花粉粒顯示其為長球形,赤道面觀為稍寬的橢圓形,極面觀為一裂圓形,極赤比為1.92。具有單溝,溝較深,溝長一端達到極端,另一端終止于極端附近(圖3-E)。花粉外壁的紋狀及表面與保持系相似,但其上有一個明顯裂口(圖3-F)。
大蔥不育系58 A2的成熟花藥與保持系的相比,藥室明顯變扁,說明花粉囊萎縮內陷(圖3-G)。成熟的花粉粒多為空癟狀(圖3-H),呈超長球形,赤道面觀為不規則形。與保持系相比,該花粉粒的單溝較深且達兩極。外壁也為腦紋狀,但表面的孔和穴相對較少(圖3~I)。
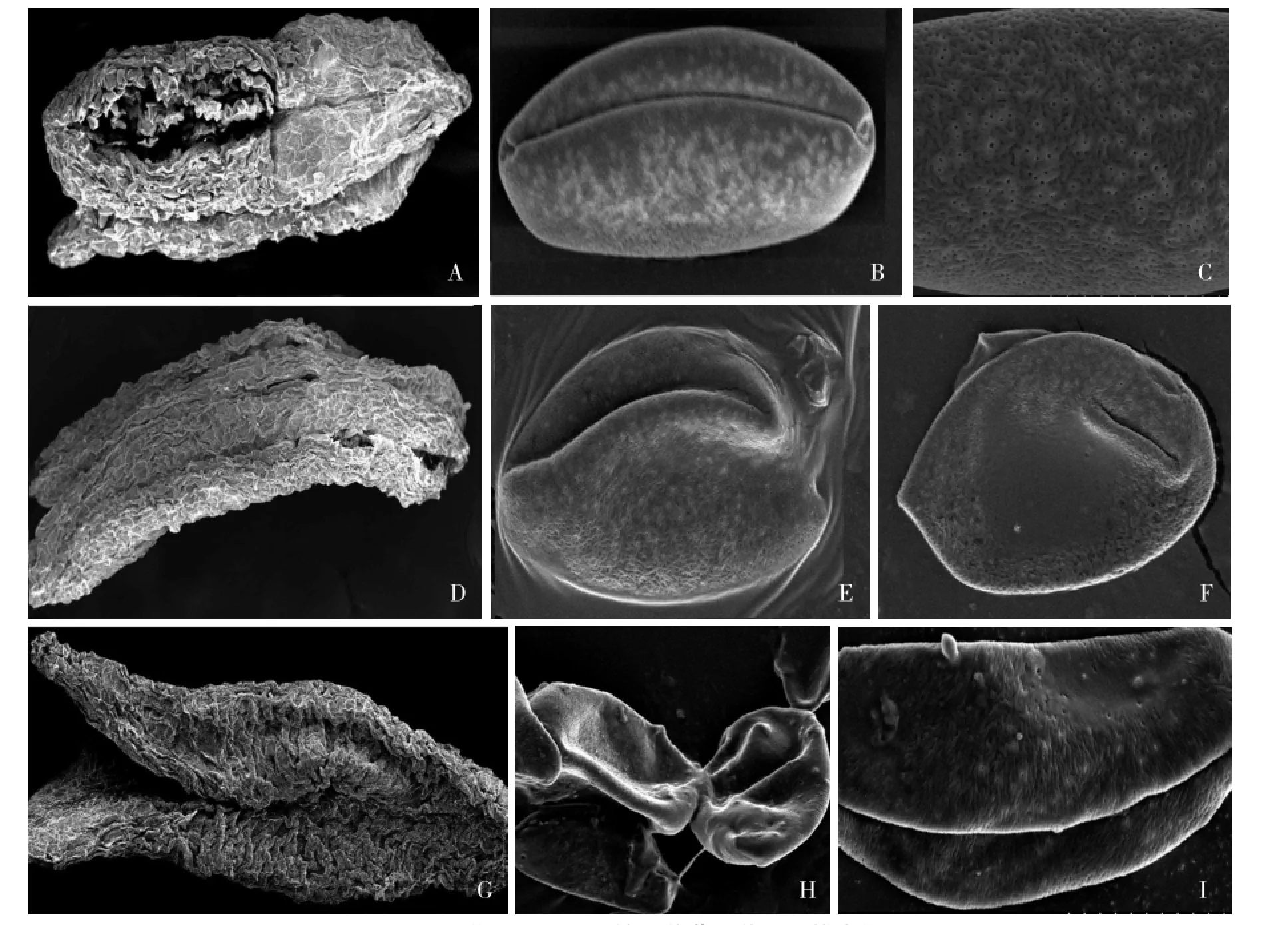
圖3大蔥不育系及保持系花藥及花粉掃描電鏡
3 結論與討論
3.1 石蠟切片觀察花藥發育
本試驗通過觀察不育系與保持系的花藥石蠟切片,發現這兩個大蔥不育植株在細胞學上以不同的方式表現出敗育現象,其中58 A1花藥中的絨氈層,在小孢子母細胞減數分裂時期就開始提前解體,其成熟期小孢子近球形,光學顯微鏡下透明;而58 A2花藥中絨氈層在小孢子單核期異常增厚,其成熟期小孢子極度皺縮,光學顯微鏡下染色極深。之前的相關研究表明了大蔥的敗育發生在單核小孢子時期(席湘媛,1991;王曉靜 等,2007),這與本試驗的結果不完全相同,這可能是由于研究對象為不同來源的雄性不育系導致的。因此,正如Laser和Lesrten(1972)報道的那樣,敗育可能發生在花粉發育的各個階段,只是有些比較明顯,而有些不明顯從而不易觀察到。
王曉靜等(2007)在對大蔥雄性不育系及保持系花藥和花粉發育的細胞學比較研究中指出,大蔥雄性不育系的小孢子有兩種敗育類型,第一種是絨氈層細胞正常發育,但中層細胞發生了延遲解體的現象,與逐漸縮小的藥室一起擠壓小孢子造成不育。第二種類型是絨氈層提前解體, 不能提供小孢子發育所需的營養, 造成小孢子的細胞質和細胞核逐漸解體, 僅剩下內空的花粉粒。謝潮添等 (2005)利用光學顯微鏡對大白菜雄性不育兩用系的不育花藥進行了觀察,認為絨氈層細胞中脂類代謝的異常可能是引起花粉敗育的原因,但經過電鏡對超微結構進行觀察,發現不育花藥中絨氈層細胞的異常不是導致花粉敗育的原因,而是花粉異常的結果。楊曉麗等(2008)在對芝麻核雄性不育系小孢子敗育過程的超微結構觀察中發現,不育小孢子外壁與可育小孢子外壁存在明顯差異,尤其是缺少基粒棒。在小孢子發育成花粉粒的過程中,不育小孢子內含物集縮并逐漸減少。
在本試驗中,58 A1與第二種敗育類型相似, 58 A2既不屬于第一種敗育類型,也不屬于第二種敗育類型。在四分體發育成熟時,本應開始程序化死亡的絨氈層細胞沒有及時解體,可能首先發生了小孢子細胞質的泄漏,絨氈層細胞吸收泄漏出來的養分而異常增厚,逐漸失去營養的小孢子核物質也逐漸解體,外形上極度皺縮。
目前報道的大多數植物雄性不育系的敗育都是由于絨氈層出現異常行為。絨氈層能夠合成胼胝質酶,分解四分體的胼胝質壁。絨氈層還可以形成并分泌孢粉素,參與花粉粒外壁的形成,并為小孢子發育提供營養。絨氈層細胞結構與功能異常均會引起雄性不育。而謝潮添等(2005)發現不育材料的花藥在造胞細胞時期就已經表現異常。
3.2 掃描電鏡觀察花粉粒
通過掃描電鏡觀察花粉粒,本試驗發現大蔥不育系58 A1及保持系的花粉粒為長球形或超長球形,極面觀都為一裂圓形,外壁紋狀及表面的空穴相似,這些特征與楊德奎等(2009)的報道結果相一致。不育系58 A2的正常花粉粒很少或者多數為干癟的花粉粒。兩個不育系花粉粒單溝的深淺或長短有區別:不育系58 A1的單溝深且只達一極,不育系58 A2的單溝也較深。從花藥外形上看,58 A2的藥室變扁,這與石蠟切片花粉囊壁向內萎縮的結果相一致。
崔連偉,杜雪晶,紀淑娟. 2004.中國大蔥育種研究概述. 吉林蔬菜, (3):37-38.
曲波,翟強,許玉風.2006. 4種忍冬屬植物花粉的形態比較.沈陽農業大學學報, 37(1):96-98.
蘇立娟. 2005.百合科植物花藥形態及花粉發育的研究〔碩士論文〕.北京:首都師范大學.
王曉靜,沈火林,楊學研,程杰山. 2007.大蔥雄性不育系及保持系花藥和花粉發育的細胞學比較研究. 中國瓜菜, (2):7-10.
王原媛,張定宇,黃國春. 2010.植物石蠟切片的固定與保存.安徽農學通報, 16(1):198-200.
吳鴻,郝剛. 2012.植物學. 北京:高等教育出版社:128-139.
席湘媛. 1991.大蔥雄性可育系及雄性不育系的花藥花粉發育的比較研究. 植物學報,33(10):770-775.
席湘媛. 2000.大蔥小孢子母細胞至二胞早期花粉發育的超微結構觀察. 云南植物研究,22(2):161-165.
謝潮添,楊延紅,葛麗麗,王瑞,田惠橋. 2005.白菜核雄性不育花藥超微結構的研究. 實驗生物學報, 38(6):501-512.
楊德奎,宋艷梅,王露. 2009.蔥屬花粉形態研究.山東師范大學學報:自然科學版,24(1):130-132.
楊曉麗,張海洋,郭旺珍,鄭永戰,苗紅梅, 魏利斌,張天真. 2008.芝麻核雄性不育系ms86-1 小孢子敗育過程的超微結構.作物學報, 34(11):1894-1900.
Inden H, Asahira T. 1990.Japanese bunching onion (Allium fistulosum L.)//Brewster J L, Rabinowitch H D. Onions and allied crops. Vol 3. Boca Raton, Florida, United States:CRC Press: 159-178.
Kaul M L H. 1988.Male sterilily in higher plants. Berlinf, Germany:Springer-Verlag Press: 10-22.
Laser K D, Lesrten N R. 1972.Anatomy and cytology of microsporogenes in cytoplasmic male setrile angiosperms. Botanical Review, 38:427-454.
Li Y Y,Yang Q,Yan J Y,Chen M. 2006.Cytoplasmic male-sterility of onion(Allium cepa L.)63A is associated with premature degeneration of the tapetum. Acta Agronomica Sinica,3(32):369-372.
Abstract:Comparision of anther and pollen development process between welsh onion(Allium fistulosum L.)CMS 58 A1, 58 A2and its maintainer line 58 B was conducted by using paraffin section technology. The pollen grains of these 3 samples were observed by scanning electron microscope. The results showed that the reason for welsh onion infertility might be caused by the following reasons:tapetum of 58 A1degenerated in advance leading microspore could not generate normally; while the tapetum of 58 A2thickened abnormally and squeezed the microspore leading its cytoplasm and nucleus flew away. The endothecium of 58 A2did not thicken in microspore stage, but the cell layer between 2 anthers was extremely thick. It caused the anthers could not split at maturity. The results of scanning electron microscope revealed that the pollen grain was of long ball or perprolate shape. The type of aperture was monocolpate. The ornamentation of outer wall was brain striatum. The sterile pollen was few or the majority of them were wizened.
Microscopic Observation on Anther and Pollen of Welsh Onion CMS 58 Male Sterile Line and Maintainer Line
WANG Chan1,2, ZHAO Hong1, LIU Qian-chu1, PEI Yan-xi2*, WANG Yong-qin1*
〔1Beijing Vegetable Research Center, Beijing Academy of Agriculture and Forestry Sciences,Key Laboratory of Biology and Genetic Improvement of Horticultural Crops (North China), Ministry of Agriculture, Beijing 100097, China ;2School of Life Science, Shanxi University, Taiyuan 030005, Shanxi, China〕
Welsh onion; CMS; Anther; Pollen; Paraffin section; Scanning electron microscopy
王嬋,碩士研究生,專業方向:蔬菜分子遺傳學,E-mail:wangchanwww@163.com
*通訊作者(Corresponding authors):裴雁曦,教授,博士生導師,專業方向:植物分子遺傳學,E-mail:peiyanxi@sxu.edu.cn;王永勤,研究員,碩士生導師,專業方向:蔬菜遺傳育種,E-mail:wyqty@sohu.com
2014-02-26;接受日期:2014-05-19
國家自然科學基金項目 (31372066),北京市農林科學院科技創新能力建設專項(KJCX20140111)
